
Abstract
This study investigated the impact of lidocaine-induced neurotoxicity on microRNA (miRNA) expression in the spinal cord of rats. Sprague–Dawley rats underwent intrathecal catheterization and were randomly assigned to receive either 10% lidocaine or normal saline for three consecutive days. Post-treatment, the paw withdrawal threshold significantly increased, accompanied by notable histopathological changes. Additionally, 470 miRNAs exhibited altered expression following lidocaine treatment, with miR-155-5p, miR-3544, and miR-675-5p showing significant changes. Gene Ontology analysis identified cellular metabolic processes as the most significantly enriched functions. Kyoto encyclopedia of genes and genomes pathway analysis revealed that the enriched signaling pathways are associated with neural injury and neuroprotection, and are involved in regulating cellular metabolism. The Mitogen-Activated Protein Kinase (MAPK) signaling pathway was notably enriched, with Mitogen-activated protein kinase kinase kinase 10 (Map3k10) and Mitogen-activated protein kinase kinase kinase 14 (Map3k14) identified as target genes of miR-155-5p. Following lidocaine treatment, there was an observed increase in the expression of MAP3K10 and MAP3K14 at both the mRNA and protein levels. These results indicate that miR-155-5p, miR-3544, and miR-675-5p might be significantly involved in lidocaine-induced neurotoxicity by influencing cellular metabolism. Furthermore, miR-155-5p/MAPK shows potential therapeutic value for treating lidocaine-induced neurotoxicity.
Introduction
Lidocaine is the preferred anesthetic for neuraxial procedures in outpatient surgery due to its rapid onset and relatively short duration of action (Frisch et al., 2018). Additionally, lidocaine’s short half-life enhances postoperative recovery, making it a preferable choice over general anesthesia and other local anesthetics (Kashanian et al., 2023). In addition to spinal anesthesia, lidocaine is also employed extensively in nerve blocks (Bantie et al., 2020; Qiu et al., 2020). However, concerns about lidocaine-induced neurotoxicity persist.
The risk of persistent lumbosacral neuropathy is approximately 1 in 1300 with a single spinal administration and 1 in 200 with continuous spinal administration of lidocaine (Johnson, 2000). Transient neurological symptoms, such as pain and numbness, are also associated with spinal lidocaine anesthesia (Forget et al., 2019). The mechanisms of lidocaine-induced neurotoxicity include apoptosis (Chen et al., 2023a; Zheng et al., 2020), mitophagy (Chen et al., 2023b; Zheng et al., 2024), and neuroinflammation (Ding et al., 2021; Zhang et al., 2022b). MicroRNAs (miRNAs), key regulators in various cellular processes, have been implicated in the neurotoxicity induced by methamphetamine abuse (Deng et al., 2022) and show potential for addressing general anesthetic-induced neurotoxicity (Minz et al., 2023). miRNAs are emerging as therapeutic targets in neuroinflammation and neurotoxicity (Brás et al., 2020; Zhao et al., 2020). Some miRNAs enhance neurological function recovery in rats with acute spinal cord injury (Lai et al., 2022) and inhibit lidocaine-induced apoptosis of dorsal root ganglia in vitro (Li et al., 2015).
This study aims to explore changes in the miRNA expression profile in the spinal cord of rats with lidocaine-induced neurotoxicity and to investigate their potential roles using bioinformatics analyses.
Materials and Methods
All procedures adhered to national and international animal care and ethical guidelines. Approval was obtained from the Medical Ethics Committee of Xiangya Hospital, Central South University (2019020099).
Animals
Nineteen adult male Sprague–Dawley rats (230–280 g, 8 weeks old) were sourced from Hunan SJA Laboratory Animal Company (Changsha, China). The rats were acclimated for 1 week under controlled temperature (22 ± 1°C), humidity (40–60%), and a 12-h light/dark cycle. Standard food and water were provided ad libitum.
Surgical procedure
The rats were anesthetized with 1–2% isoflurane (RWD, Shenzhen, China). A PE-10 microspinal catheter (Smiths Medical, Minneapolis, MN, USA) was then introduced into the subarachnoid space via the L5/L6 intervertebral space. The catheter tip was advanced approximately 2.5 cm cranially to the L3 level. Post-catheterization observation lasted 3 days. Rats with traumatic nerve injury, neurological deficits, or catheterization failure were excluded. A total of three rats were excluded.
Model establishment
A total of 10% lidocaine was made by dissolving 100 mg of lidocaine hydrochloride powder (purity >99%, CAS number 6108–05-0; Meilunbio, Dalian, China) in 1 mL of preservative-free normal saline. pH and osmolarity were measured using the SevenCompact™ S220 (Mettler Toledo, Zurich, Switzerland) and Osmolab One (Beijing YASN, Beijing, China), respectively (see Supplementary Table S1 for details). Rats were randomly divided into Group L and Group N, with eight rats in each group. Rats in Group L received intrathecal injections of 10% lidocaine, while rats in Group N received intrathecal injections of normal saline. Rats received intrathecal injections of 10% lidocaine or normal saline at a dose of 0.12 µL/g daily for three consecutive days. Injections were administered between 8 and 9 AM, with each injection lasting 90 s. Afterward, 10 µL of normal saline was administered through the catheter for flushing.
Neurobehavioral tests
The paw withdrawal threshold (PWT) was measured using Electronic von Frey (IITC Life Science Instruments, Woodland Hills, CA, USA) to evaluate neurobehavioral changes. These tests were conducted by a blinded investigator between 8 and 9 AM, both prior to the initial injection on day 0 (baseline) and 24 h after the last injection on day 4. Each rat underwent three measurements for each hind paw during the tests.
Histopathological analysis
Spinal cord segments from L2 to L6 were harvested and fixed in 4% paraformaldehyde. Paraffin sections were then prepared for hematoxylin and eosin (H&E) and toluidine blue staining. For H&E staining, the paraffin sections were treated with hematoxylin for 5 min, followed by eosin for 2 min. Toluidine blue staining was performed by incubating the sections in 1% toluidine blue at room temperature for 2–3 min. For immunofluorescence analysis, frozen sections were utilized. After antigen retrieval using sodium citrate buffer and blocking with 5% bovine serum albumin, primary antibodies Iba1 (011-27991, 1:500; Wako, Osaka, Japan) and glial fibrillary acidic protein (3670S, 1:200; CST, Danvers, MA, USA) were applied and incubated overnight at 4°C. Following washes with PBS, the corresponding secondary fluorescent antibodies were introduced. The H&E and toluidine blue stained sections were examined using a light microscope (Leica DMLB2; Leica Microsystems, Wetzlar, Germany), while the immunofluorescence slides were examined with a fluorescence microscope (Leica DMI4000; Leica Microsystems, Wetzlar, Germany).
RNA isolation
The L2 to L6 spinal cord segments were removed en bloc and stored in liquid nitrogen. The extraction of total RNA was performed using Trizol reagent (Invitrogen, Carlsbad, CA, USA), and subsequent purification was conducted using RNeasy Mini Kit (Qiagen, Hilden, Germany) following the manufacturer’s guidelines. The integrity of RNA was assessed using agarose gel electrophoresis, whereas the purity and concentration of RNA were determined using a Nanodrop ND-1000 Spectrophotometer (Thermo Fisher Scientific Inc., Waltham, MA, USA). After quality control, the RNA was preserved at a temperature of −80°C.
Microarray analysis
miRNAs were labeled with Hy3™ fluorescent dye using the miRCURY™ Array Power Labeling Kit (Exiqon, Vedbæk, RM, Denmark) according to the manufacturer’s instructions. Labeled miRNAs were hybridized to the miRCURY™ LNA Array (Exiqon), which targets all registered miRNAs in miRBASE version 19.0. Hybridization was conducted using a 12-array Hybridization System (NimbleGen Systems, Inc., Madison, WI, USA). The slides were subsequently subjected to a washing procedure using the Wash Buffer Kit (Exiqon), followed by scanning with Axon GenePix 4000B microarray scanner (Axon Instruments, Foster City, CA, USA). Image intensity was analyzed using GenePix Pro (version 6.0; Axon Instruments), with an average of four replicates per probe. Data were normalized using the formula: Normalized Data = (Foreground-Background)/Median, where the Median is the 50th percentile of intensity. miRNAs were statistically analyzed between the two groups.
Establish the network of miRNAs and target mRNAs
The miRDB v5 database (accessible at http://mirdb.org/miRDB/) was utilized for the prediction of target genes regulated by miRNAs. Based on these predictions, a miRNA-mRNA network was constructed. miRNAs and mRNAs with a target score of at least 70 were selected for network construction using Cytoscape software (version 3.9.0; Cytoscape Consortium, San Diego, CA, USA).
Gene ontology and Kyoto encyclopedia of genes and genomes pathway analyses
Gene ontology (GO) and Kyoto encyclopedia of genes and genomes (KEGG) pathway analyses were conducted on the targeted genes to predict the functions of miRNAs. GO (http://geneontology.org/) is an internationally recognized system for the classification of gene function, encompassing three domains: biological process, molecular function, and cellular component. Data were downloaded using the R package “GO.db” and analyzed with “TopGO.” KEGG pathway analysis (http://www.genome.jp/kegg/) identified pathways associated with the miRNAs. Statistical significance was evaluated using Fisher’s exact test, while the false discovery rate was calculated by the Benjamini-Hochberg method. The Enrichment Score was defined as “-log10(p value),” while the statistical significance was determined using a threshold of p < 0.05.
Quantitative real-time polymerase chain reaction (RT-qPCR)
For miRNA and mRNA analyses, total RNA was reverse transcribed into cDNA using the All-in-One™ miRNA First-Strand cDNA Synthesis Kit and the All-in-One™ First-Strand cDNA Synthesis Kit (GeneCopoeia, Suzhou, China). cDNA was then amplified with the Applied Biosystems 7300 Fast Thermocycler (Applied Biosystems, Foster City, CA, USA) using the All-in-One™ miRNA qPCR Kit and the All-in-One™ qPCR Kit (GeneCopoeia). U6 and β-actin were used as housekeeping genes for miRNA and mRNA, respectively. Relative gene expression levels were analyzed using the 2−ΔΔCT method and each experiment was performed in triplicate. Primers for miRNA were procured from GeneCopoeia with the following catalog numbers: rno-miR-3544 (cat#RmiRQP1798), rno-miR-675-5p (cat#RmiRQP1102), rno-let-7f-5p (cat#RmiRQP0012), rno-miR-155-5p (cat#RmiRQP0890), and rno-miR-1-3p (cat#RmiRQP1188), U6 primer (cat#RmiRQP9003). mRNA primers are listed in Supplementary Table S2.
Western blot
Total protein was isolated from spinal tissues using Radio Immunoprecipitation Assay Lysis Buffer (Beyotime, Shanghai, China). The concentration of protein was measured with the Bicinchoninic Acid Protein Assay Kit (Beyotime). Membranes were incubated with primary antibodies, including GAPDH (ab181602, 1:5000; Abcam, Cambridge, UK), MAP3K10 (BS61034, 1:500; Bioworld Technology, Inc., Akron, OH, USA), and MAP3K14 (A11585, 1:500; ABclonal, Inc., Wuhan, China). After washing, the membranes were treated with a secondary antibody (ab288151, 1:2000; Abcam). Signals were detected using the Bio-Rad GelDoc 2000 system (Bio-Rad, Hercules, CA, USA). Intensity was quantified using ImageJ software (version 1.53a; NIH, Bethesda, MD, USA). GAPDH was used for normalization.
Statistical analysis
Quantitative data are presented as mean ± standard deviation and analyzed using GraphPad Prism (version 9.5.0; GraphPad Software Inc., San Diego, CA, USA). Comparative analyses between two groups with normal distribution were made using Student’s t-test. Behavioral data were analyzed with two-way ANOVA, followed by Bonferroni’s multiple-comparison test. The statistical significance was determined using a threshold of p < 0.05.
The application of artificial intelligence (AI)
In this study, we employed OpenAI’s ChatGPT-4.0 for language refinement, with initial usage commencing on August 27, 2024. We provided draft text and used prompts such as “Please correct the grammar in this paragraph” or “Please refine this paragraph to enhance readability” to direct the AI’s responses. While ChatGPT-4.0 exhibited commendable performance in language enhancement, it is essential to acknowledge that the generated text may exhibit biases inherent in the training data. Furthermore, we meticulously reviewed the refined content and confirmed its accuracy and relevance.
Results
Neurobehavioral dysfunction and histopathological changes in a rat model of lidocaine-induced neurotoxicity
On day 0, before treatment, there was no significant difference in PWT between Group L and Group N (p > 0.05; Fig. 1A, n = 8). On day 4, following treatment, PWT in Group L was significantly higher compared with Group N (p < 0.0001; Fig. 1A, n = 8). H&E staining (Fig. 1B, n = 3) and toluidine blue staining (Fig. 1C, n = 3) of spinal cord sections from Group N revealed normal nerve fiber structure. In contrast, Group L exhibited significant histopathological alterations, characterized by neuronal damage and glial cell proliferation. Notably, Group L demonstrated proliferation and activation of microglia (Fig. 1D, n = 3) and astrocytes (Fig. 1E, n = 3), suggesting that neuroinflammatory processes contribute to lidocaine-induced nerve injury.
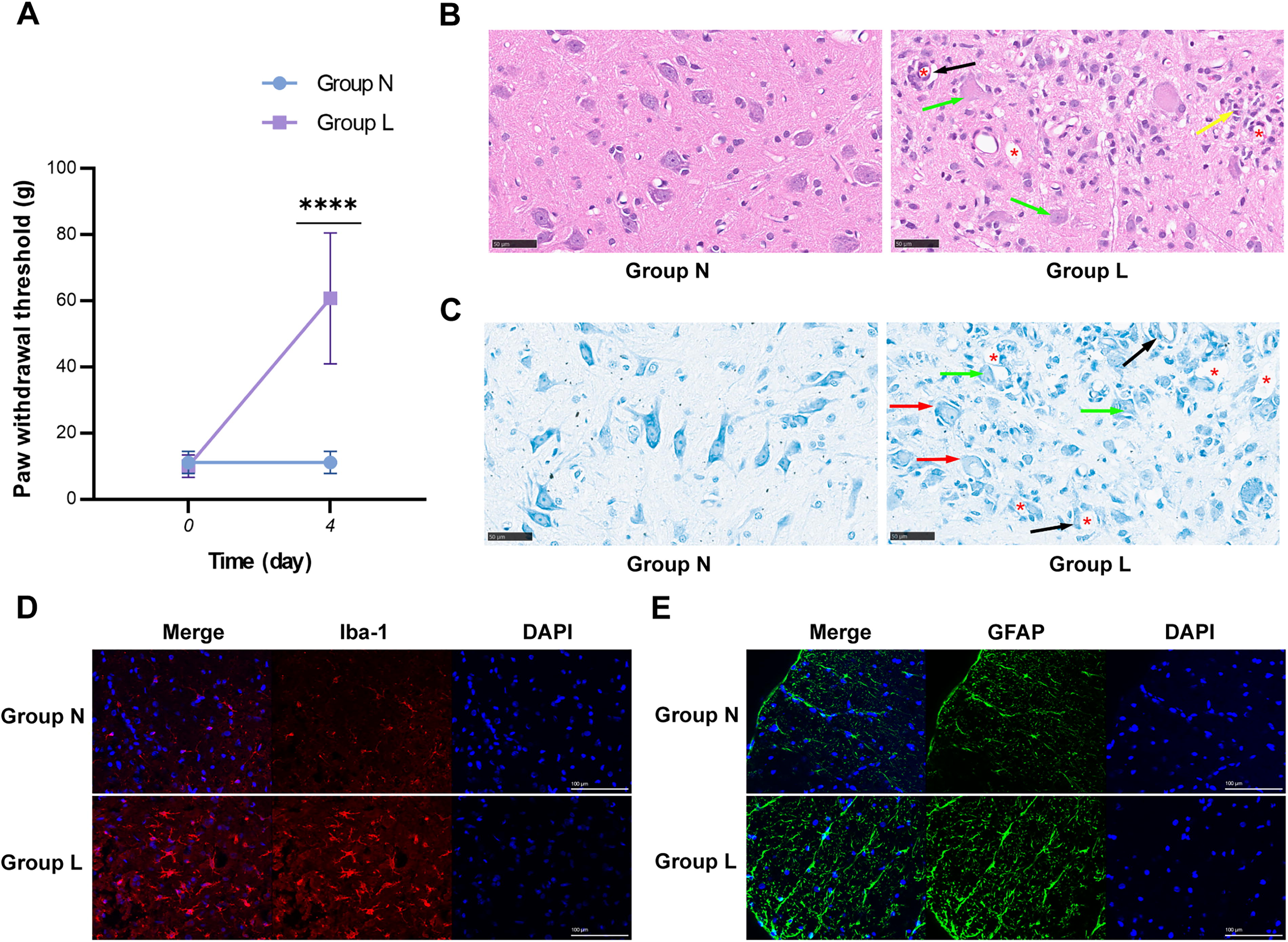
Rat model of lidocaine-induced neurotoxicity.
miRNA profiles based on microarray analysis
Median normalization of microarray raw data corrected overall deviations in the experiment (Fig. 2A, n = 3). In the normalized data, 470 miRNAs were identified (Supplementary Table S3). Hierarchical clustering analysis of these 470 miRNAs (282 upregulated and 188 downregulated) revealed differences between Group L and Group N (Fig. 2B, n = 3). The volcano plot (Fig. 2C, n = 3) indicated that 72 miRNAs were upregulated (fold change ≥1.5, Group L vs. Group N), while 27 miRNAs were downregulated (fold change ≥1.5, Group N vs. Group L). Among these, five differentially expressed (DE) miRNAs (let-7f-5p, miR-1-3p, miR-155-5p, miR-675-5p, and miR-3544) showed statistically significant changes (p < 0.05). Specifically, let-7f-5p, miR-1-3p, and miR-155-5p were upregulated, while miR-675-5p and miR-3544 were downregulated in Group L compared with Group N, as shown in the heatmap (Fig. 2D, n = 3). Detailed information on these five DE miRNAs is provided in Table 1.

Microarray analysis of miRNAs in the spinal cord of rats with lidocaine-induced neurotoxicity (n = 3).
Detailed Information on Differentially Expressed miRNAs
Expression verification of the DE miRNAs
The expression of the five DE miRNAs (let-7f-5p, miR-1-3p, miR-155-5p, miR-675-5p, and miR-3544) identified by microarray analysis was confirmed by RT-qPCR, which showed consistent expression patterns. Statistically significant changes were observed in miR-155-5p (p < 0.01), miR-3544 (p < 0.001), and miR-675-5p (p < 0.001) in the RT-qPCR analysis (Fig. 3, n = 5).
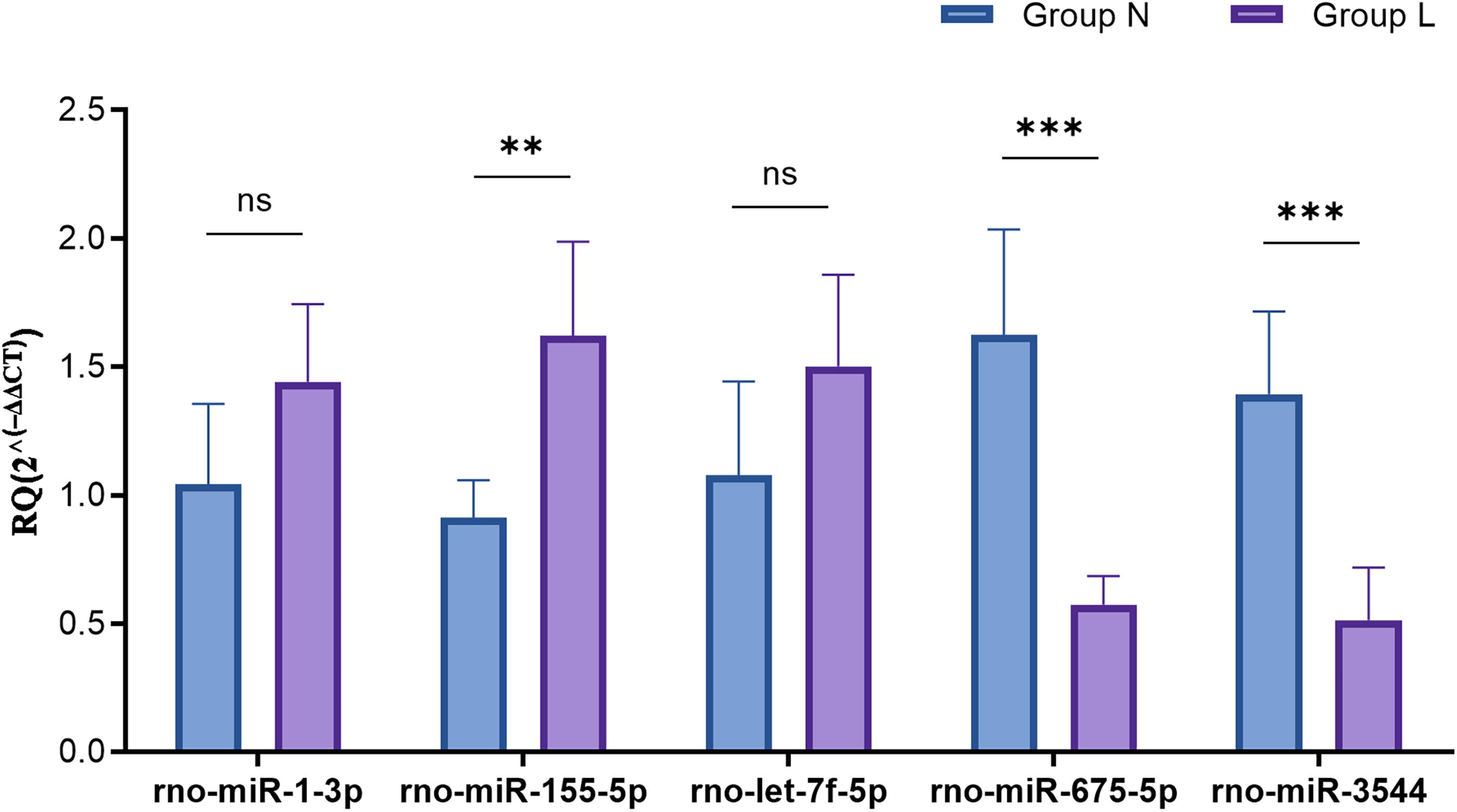
Verification of the expression changes of the DE miRNAs between the two groups using RT-qPCR (n = 5). **p < 0.01, ***p < 0.001. RT-qPCR, quantitative real-time polymerase chain reaction.
The network of the verified DE miRNAs and their target mRNAs
miRNA-mRNA networks were constructed for miR-155-5p, miR-3544, and miR-675-5p (Fig. 4), with each DE miRNA predicted to target 103, 56, and 9 mRNAs, respectively. Detailed information on these predicted target mRNAs is provided in Supplementary Table S4.
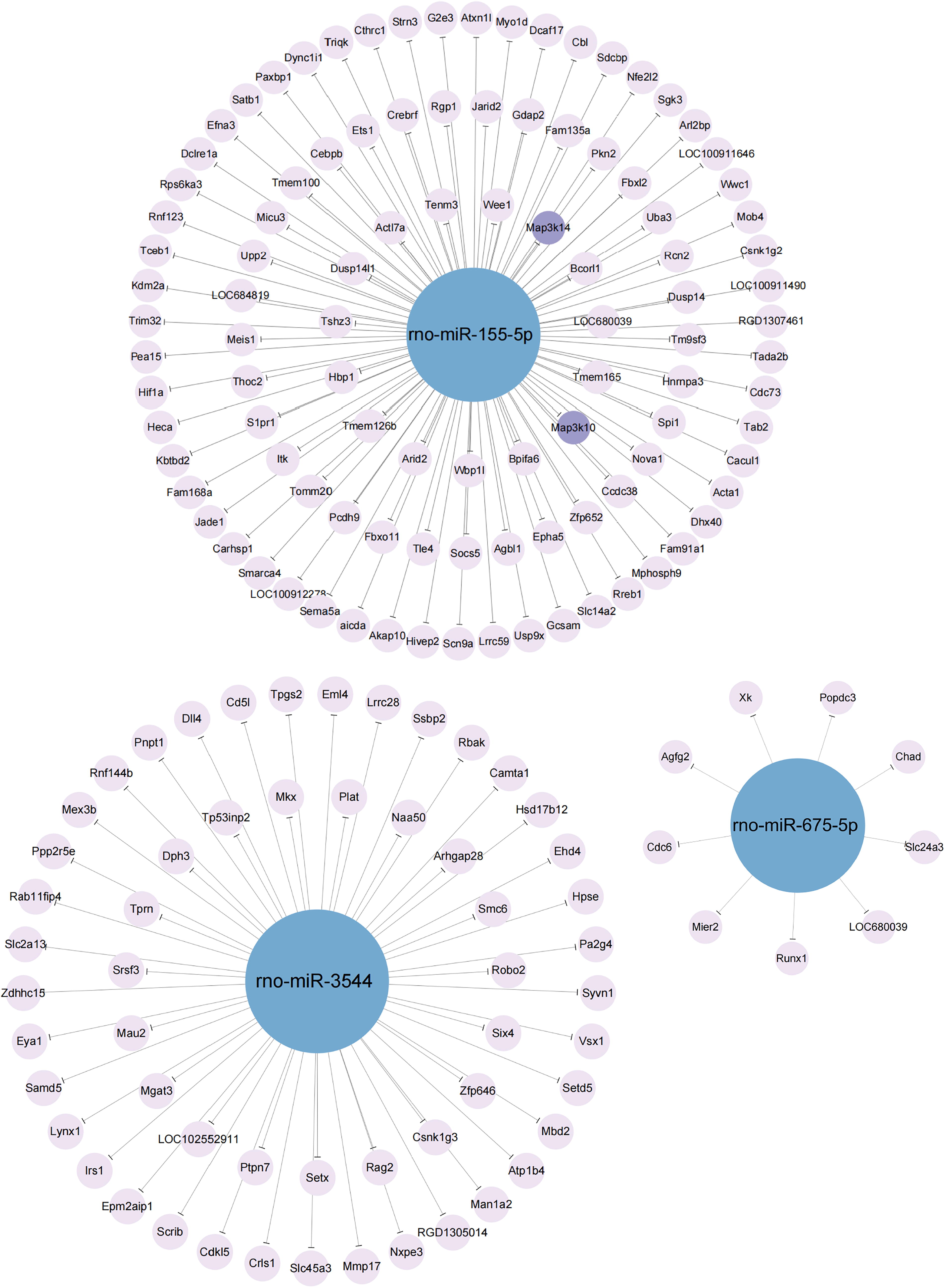
The network of the verified DE miRNAs (miR-155-5p, miR-3544, and miR-675-5p) and their predicted target mRNAs. The blue nodes represent miRNAs, while the purple nodes represent mRNAs. The two deep purple nodes (Map3k10 and Map3k14) were selected for further verification. Map3k10, Mitogen-activated protein kinase kinase kinase 10; Map3k14, Mitogen-activated protein kinase kinase kinase 14.
Functional and pathway analyses of the verified DE miRNAs
GO enrichment analysis of miR-155-5p, miR-3544, and miR-675-5p revealed significant enrichment in biological processes related to metabolic regulation (Fig. 5A). The most enriched molecular functions included protein binding, ion binding, and enzyme binding (Fig. 5B). For cellular components, the cytoplasm, nucleus, and membrane-bounded organelles were the most significantly enriched (Fig. 5C). KEGG pathway analysis identified the Mitogen-Activated Protein Kinase (MAPK) signaling pathway, cyclic Guanosine Monophosphate-Protein Kinase G (cGMP-PKG) signaling pathway, Arginine biosynthesis, and Prolactin signaling pathway as significantly enriched (Fig. 5D).

GO and KEGG analyses of miR-155-5p, miR-3544, and miR-675-5p. Bubble plots illustrate the top 10 statistically significant GO terms for
Expression of mitogen-activated protein kinase kinase kinase 10 (Map3k10) and mitogen-activated protein kinase kinase kinase 14 (Map3k14)
The deep purple nodes in Figure 4 represent Map3k10 and Map3k14, which are targets of miR-155-5p with high target scores and are involved in the MAPK signaling pathway. The binding sites for miR-155-5p are shown in Figure 6A. RT-qPCR and Western blot analyses were performed to assess the expression. Results indicated a significant increase in mRNA levels of Map3k10 and Map3k14 in Group L compared with Group N (p < 0.001; Fig. 6B, n = 6). Although the protein levels of MAP3K10 (p = 0.1498) and MAP3K14 (p = 0.0698) were also higher in Group L, the changes were not statistically significant (Fig. 6C and D, n = 3).

The binding sites and expression levels of Map3k10 and Map3k14.
Discussion
The utilization of lidocaine in clinical settings for local, regional, and axial anesthesia has the potential to cause nerve damage. This study aimed to develop a rat model of lidocaine-induced neurotoxicity using intrathecal injection of 10% lidocaine, simulating clinical spinal block administration. Hypoalgesia observed in rats 24 h post-injection reflects the neurobehavioral change, affirming the model’s validity (Zhao et al., 2024). Further histopathological analysis confirmed that lidocaine induces neurotoxic effects (Koo et al., 2021; Kroin et al., 2010), accompanied by neuroinflammatory responses consistent with our previous research findings (Zhang et al., 2022b).
Local anesthetic neurotoxicity involves several signaling pathways, including the Caspase, Phosphoinositide 3-Kinase (PI3K)/Protein Kinase B (Akt), and MAPK pathways (Verlinde et al., 2016). These pathways are involved in various biological processes such as apoptosis, proliferation, differentiation, and metabolism in nerve tissue (Li et al., 2021; Seo et al., 2002; Zhang et al., 2022a). While it is suggested that miRNAs regulate these pathways, studies on miRNAs’ role in lidocaine-induced neurotoxicity are limited. Through microarray analysis, we identified miRNAs with altered expression in response to lidocaine. RT-qPCR further revealed significant expression changes in miR-155-5p, miR-3544, and miR-675-5p.
The role of the three DE miRNAs in lidocaine-induced neurotoxicity was investigated by predicting their target mRNAs and conducting functional and pathway analyses. GO analysis revealed that miR-155-5p, miR-3544, and miR-675-5p predominantly function in the cytoplasm and membrane-bound organelles of neural cells, including mitochondria, endomembrane system, and nucleus. These miRNAs likely influence metabolic processes by regulating protein, enzyme, and kinase binding in neural cells. The metabolism of neural cells, including energy, lipid, and amino acid metabolism (Boison and Steinhäuser, 2018; Lee et al., 2021), which is closely related to the cellular structures and molecular functions identified in the GO analysis. These findings indicate that miR-155-5p, miR-3544, and miR-675-5p may regulate macromolecule metabolism in spinal cells.
Cellular metabolism is crucial for maintaining homeostasis by facilitating enzyme-catalyzed reactions that regulate the activity, localization, interaction, and stability of proteins and macromolecules, thereby modulating key signaling pathways (Chen and Tsai, 2022; Yang, 2005). Additionally, cell metabolism plays a critical role in nerve injury and neuroprotection (Lee et al., 2020; Vellingiri et al., 2022), indicating that miR-155-5p, miR-3544, and miR-675-5p may influence lidocaine-induced neurotoxicity through metabolic modulation.
KEGG analysis of miR-155-5p, miR-3544, and miR-675-5p revealed enriched pathways including MAPK, Arginine biosynthesis, cGMP-PKG, and Prolactin signaling pathway. MAPKs are crucial in neuroinflammation and neurodegeneration (Iba et al., 2023), with inhibition reducing inflammatory metabolites in microglia and providing neuroprotection (Plastira et al., 2020). Lidocaine activates p38 MAPK in dorsal root ganglion neurons, contributing to neurotoxicity (Haller et al., 2006); however, p38 MAPK inhibitors can enhance neuronal survival and attenuate neurotoxicity (Lirk et al., 2006). Post-nerve injury modulation of MAPK reduces inflammation, apoptosis, and neuropathic pain while promoting nerve regeneration (Huang et al., 2024). Prolactin supports neuronal plasticity via MAPK pathways (Molina-Salinas et al., 2021), while polyamines from L-arginine and cationic arginine-rich peptides offer neuroprotective effects (MacDougall et al., 2019; Saha et al., 2023). The cGMP-PKG pathway supports neural stem cell proliferation and neuroprotection (Huan et al., 2020; Ölmestig et al., 2017). Together, these enriched signaling pathways play significant roles in neural injury and protection.
MAPK signaling pathway is involved in metabolic reprogramming and regulates glucose and lipid metabolism (Asl et al., 2021; Chen et al., 2024). Increased activity of MAPK promotes glucose uptake and mitochondrial function (Bengal et al., 2020) and influences lipid metabolism, which can impact conditions such as non-alcoholic fatty liver disease (Yao et al., 2023). MAPK/PI3K pathway is also key in stimulating ribosome biogenesis and protein synthesis (Anjum and Blenis, 2008). L-arginine shifts the metabolism of neural stem cells from glycolysis to oxidative phosphorylation (Xu et al., 2023), and its metabolites, including nitric oxide and glutamate, show potential in treating metabolic disorders (Szlas et al., 2022). cGMP-PKG pathway contributes to the proliferation of neural stem cells after cerebral ischemia and aids glucose uptake (Tian et al., 2018; Xiao et al., 2021). Additionally, prolactin levels rise in response to metabolic changes, supporting adaptation (Macotela et al., 2022). These signaling pathways, as noted in KEGG, interact and work together to regulate cellular metabolism.
KEGG analysis of miR-155-5p, miR-3544, and miR-675-5p revealed their significant involvement in neurotoxicity and neuroprotection through intracellular metabolic modulation, as indicated by GO analysis. These miRNAs regulate cellular metabolism in lidocaine-induced neurotoxicity, potentially impacting neurological function.
The MAPK signaling pathway, consisting of various MAPKs and MAP kinase kinases, is crucial for cellular signaling via a phosphorylase cascade (Fang and Richardson, 2005; Hepworth and Hinton, 2021). KEGG analysis indicated that this pathway is significantly enriched by the three DE miRNAs. It plays an important role in various cellular metabolic processes and in regulating neurotoxicity from local anesthetics. Notably, miR-155-5p targets Map3k10 and Map3k14, both of which are components of the MAPK pathway. These kinases are vital for gene transcription in neural cells and are linked to neurogenesis, neuroplasticity, and neuroinflammation (Mao et al., 2016; Upadhyay et al., 2019). Experimental findings revealed higher mRNA and protein levels of MAP3K10 and MAP3K14 in the spinal cord of model rats, suggesting their involvement in neurotoxicity.
This study underscores the interconnected roles of various signaling pathways in lidocaine-induced neurotoxicity, affecting neuronal metabolism and function, and enhances the understanding of the biological mechanisms behind lidocaine-induced neurotoxicity. The validation of miR-155-5p expression and its target genes, Map3k10 and Map3k14, offers new potential targets for future therapeutic strategies aimed at neuroprotection during the use of local anesthetics. This may lay the groundwork for developing new treatment approaches by modulating the pathways through which miR-155-5p influences cellular metabolism, particularly those associated with energy metabolism and oxidative stress. These findings suggest promising avenues for therapeutic interventions in this emerging field. However, further experimental validation is necessary to confirm these results and to evaluate the changes in the expressions of MAP3K10 and MAP3K14 following miR-155-5p intervention, as well as the neurobehavioral effects and other indicators of neurotoxicity.
Footnotes
Acknowledgment
The authors thank all the members of the Anesthesiology Laboratory at Xiangya Hospital, Central South University, for their assistance.
Authors’ Contributions
L.L.: Investigation, data curation, formal analysis, visualization, funding acquisition, and writing—original draft. Z.Z.: Conceptualization, methodology, investigation, and supervision. Z.J.: Investigation and data curation. A.T.: Validation and data curation. Q.L.: Conceptualization, methodology, formal analysis, visualization, funding acquisition, project administration, and writing—review and editing.
Disclosure Statement
The authors declare no competing interests.
Funding Information
This work was funded by the Natural Science Foundation of Hunan Province (Grant No: 2021JJ41002 and 2024JJ8182).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.