
Abstract
Exosome-delivered circular RNAs (circRNAs) are recognized as a key mechanism that regulates osteosarcoma (OS) progression. The purpose of this study is to discover the role of a novel circRNA hsa_circ_0000116 from exosomes in OS progression. Transmission electron microscopy, nanoparticle tracking analysis, and western blotting were used to identify the exosomes isolated from two OS cell lines (HOS and MG-63). After coculturing exosomes with OS cells and transfecting hsa_circ_0000116 knockdown vector into OS cells, cell function experiments, including cell counting kit-8, wound healing, and Transwell experiments, were performed to assess the change of OS cell malignant phenotype. In addition, the levels of PI3K/Akt/mTOR and p38/MAPK pathways-associated proteins were measured using western blotting. Exosomes with around 100 nm in diameter were successfully isolated from HOS and MG-63 cells, and promote OS cells to proliferate, migrate, and invade. hsa_circ_0000116 was upregulated in OS-derived exosomes, and silencing hsa_circ_0000116 declined the exosome-induced OS cell malignancy. In addition, inhibiting hsa_circ_0000116 effectively inhibited exosome-mediated activation of PI3K/Akt/mTOR and p38/MAPK pathways. In conclusion, exosomal hsa_circ_0000116 can facilitate OS cell malignancy by inducing the activation of PI3K/Akt/mTOR and p38/MAPK pathways. The findings of this study may identify novel molecular mechanisms driving OS progression and provide novel therapeutic targets for OS.
Introduction
Osteosarcoma (OS), a primary malignant bone tumor in children and adolescents, has high tendency to disseminate to develop metastasis (Jiang et al., 2023; Sheng et al., 2021). Despite advances in treatment strategies, including surgery, chemotherapy, and radiation, the prognosis for patients with OS remains poor, especially in those with metastasis or recurrence (Kattepur et al., 2021; Miwa et al., 2020). Therefore, a deeper understanding of the molecular mechanisms driving OS progression to find novel therapeutic targets is necessary.
Exosomes secreted by various cell types are small, lipid bilayer-enclosed extracellular vesicles, and have gained significant attention in recent years due to their role in mediating intercellular communication (Ghaffari et al., 2023; Kumar et al., 2024; Moghassemi et al., 2024). It is reported that exosomes carry a diverse cargo, including proteins, lipids, RNAs, and DNA, thereby transferring molecular signals between cells in both normal and pathological conditions (Aheget et al., 2020; Kalluri and LeBleu, 2020). Exosome-mediated transfer of noncoding RNAs with a covalently closed-loop structure called circular RNAs (circRNAs) has been increasingly recognized as a key mechanism that regulates cancer progression (Babin et al., 2021; Geng et al., 2020; Hussen et al., 2023; Xu et al., 2021). For example, exosomal circ-0044516 was reported to promote prostate cancer progression by facilitating cell proliferation and metastasis (Li et al., 2020). Exosome-derived circ_002136 was found to facilitate the progression of hepatocellular carcinoma (Yuan et al., 2022). Exosomal circSHKBP1 played the oncogenic role in gastric cancer (Xie et al., 2020). By microarray analysis, a circRNA hsa_circ_0000116 expression was found to be elevated in exosomes from OS samples. However, whether hsa_circ_0000116 in exosomes can regulate OS progression has not been explored.
PI3K/Akt/mTOR and p38/MAPK pathways are considered crucial regulators of cell growth, survival, and apoptosis, and are often aberrantly activated in cancers (Garcia-Hernandez et al., 2021; Glaviano et al., 2023). Mounting evidence suggests that the dysregulation of these pathways contributes to OS progression. PI3K/AKT/mTOR activated in OS expedites the OS cell malignant phenotype by interacting with noncoding RNAs, including circRNAs (Shao et al., 2024; Yang et al., 2021). Similarly, the p38/MAPK pathway activated in OS can also promote OS progression (Hsu et al., 2019; Yong et al., 2018), although there is no study to reveal its correlation with circRNAs.
This study aimed to investigate the role of exosome-delivered hsa_circ_0000116 in OS progression, focusing on its potential regulation of PI3K/Akt/mTOR and p38/MAPK pathways. By elucidating these mechanisms, we hope to identify novel therapeutic targets that could improve treatment outcomes for patients with OS.
Materials and Methods
Cell culture and transfection
Two OS cell lines, including HOS (CL-0360, Procell, China) and MG-63 (CL-0157, Procell), were kept in Minimum Essential Medium (MEM) medium (Procell) with 10% Fetal Bovine Serum (FBS, Procell) to perform cell culture at 37°C and 5% CO2. When the cell confluence was more than 60%, siRNA targeting hsa_circ_0000116 (si-hsa_circ_0000116, GenScript Biotech Corporation, China) and its negative control (si-NC, GenScript Biotech Corporation) were transfected into two OS cells using the SweTransRNA Transfection Reagent (G1806-0.5ML, Wuhan Servicebio Biotechnology Co., Ltd., China) for 48 h of transfection.
Exosome isolation and coculture
The culture medium supernatant of HOS and MG-63 cells was centrifuged for 15 min at 300 × g, 20 min at 2000 × g, 30 min at 10,000 × g, and 90 min at 100,000 × g. Then, the obtained supernatant after filtration was resuspended with phosphate buffered saline (PBS), and centrifuged at 100,000 × g for 90 min. Finally, the exosome precipitation was collected and resuspended to determine using the BCA Protein Assay Kit (Abcam, USA). Transmission electron microscopy (TEM) was used to observe the exosome morphology, while nanoparticle tracking analysis (NTA) was applied to analyze the size distribution of exosomes.
As for coculture, 10 μg of exosomes isolated from HOS and MG-63 cells were resuspended with MEM medium and added into HOS and MG-63 cells for cell culture, respectively.
Cell counting kit-8 assay
The OS cells were cocultured with exosomes, and transfection was performed with the cell counting kit-8 (CCK8) assay for detecting cell proliferation. Briefly, the OS cells (2 × 104 cells/mL) were seeded in a 96-well plate for cell culture. At different times of coculture and transfection (0, 24, 48, and 72 h), the cells were collected and added with 10 μL of CCK8 solution (G4103-1ML, Wuhan Servicebio Biotechnology Co., Ltd., China) for a 2-h incubation. The optical density value at 450 nm was detected by the YP-96C microplate reader (YouYunPu, China).
Wound healing assay
A total of 1 × 105 cells after coculture and transfection were seeded into a 6-well plate for cell culture until >90% cell confluence. A 200 μL pipette was used to create a straight-line scratch, and then the floating cells were washed using PBS. An inverted microscope was used to image the scratch at 0 h. After cell culture for 24 h, the scratch at 24 h was also imaged by the inverted microscope. The migration rate was calculated to assess the cell migration ability.
Transwell assay
A total of 1 × 105 cells after coculture and transfection in serum-free MEM medium were seeded into the upper chamber of Transwell that was pre-coated with Matrigel (G4130-5ML, Wuhan Servicebio Biotechnology Co., Ltd., China). At the same time, MEM medium with 10% FBS filled in the lower chamber. After a 24-incubation, the invasive cells were fixed, stained, and imaged by an inverted microscope.
Quantitative Real-Time Polymerase Chain Reaction
Total RNA was isolated from OS cells by the RNA Extraction Solution (G3013-100ML, Wuhan Servicebio Biotechnology Co., Ltd., China). Then, 1 μg of isolated RNA and the primers in Table 1 were used to perform qRT-PCR using the SweScript One-Step RT-PCR Kit (G3335-50, Wuhan Servicebio Biotechnology Co., Ltd.). The relative hsa_circ_0000116 expression was measured using the 2-ΔΔCt method and GAPDH as internal control.
Primers Used in This Study
Western blotting
RIPA lysis buffer (G2002-100ML, Wuhan Servicebio Biotechnology Co., Ltd., China) was used to lyse OS cells or exosomes for total protein isolation. After quantifying the isolated proteins using the BCA Protein Quantitative Detection Kit (G2026-200T, Wuhan Servicebio Biotechnology Co., Ltd.), 20 μg of isolated protein sample were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), transferred to polyvinylidene fluoride (PVDF) membranes (Wuhan Servicebio Biotechnology Co., Ltd.), and blocked in skim milk for 3 h. Then, the proteins were incubated by the following antibodies: exosomal markers, including anti-CD9 (GB115697-100, Wuhan Servicebio Biotechnology Co., Ltd.) and anti-CD63 (ab134045, Abcam, USA); PI3K/Akt/mTOR pathway-associated proteins, including phosphorylated anti-PI3K (p-PI3K, abs172917, Absin, China), anti-PI3K (abs173390, Absin), phosphorylated anti-Akt (p-Akt, abs130888, Absin), anti-Akt (abs131788, Absin), phosphorylated anti-mTOR (p-mTOR, abs155636, Absin), and anti-mTOR (abs156124, Absin); p38/MAPK pathway-associated proteins, including phosphorylated anti-p38 (p-p38, AF5884, Beyotime, China) and anti-p38 (AF1111, Beyotime,); and internal control protein anti-GAPDH (GB11002-100, Wuhan Servicebio Biotechnology Co., Ltd.). After incubating primary antibodies overnight, a secondary antibody (GB111738-100, Wuhan Servicebio Biotechnology Co., Ltd.) was added into membranes for further incubation for 2 h. Finally, the proteins in membranes were visualized by the Hypersensitive ECL Chemiluminescence Kit (G2020-50ML, Wuhan Servicebio Biotechnology Co., Ltd.).
Ethics statement
This study does not involve animals or human participants.
Statistical analysis
All data were analyzed by GraphPad Prism with Student’s t-test and one-way analysis of variance (ANOVA) to analyze the differences between two groups and among multiple groups. All data from three biological experiments were shown as mean ± SD. The differences were considered significant if p value was less than 0.05.
Results
OS cell-derived exosomes facilitate OS cell malignancy
Using differential ultracentrifugation, the exosomes were isolated from two OS cell lines (HOS and MG-63). TEM and NTA showed that the exosomes with around 100 nm in diameter displayed a round shape (Fig. 1A). After detecting exosomal markers (CD9 and CD63) by western blotting, it was found that CD9 and CD63 were positively expressed in the exosomes from OS cells (HOS-Exo and MG-63-Exo, Fig. 2B). The results of TEM, NTA, and western blotting proved that the exosomes were successfully isolated from OS cells. Next, the exosomes were cocultured with OS cells to perform the following cell functional experiments. CCK8 assay displayed that exosomes induced OS cell proliferation (Fig. 1C). After performing wound healing assay, it was found that the ability of OS cell migration was enhanced by coculturing with exosomes (Fig. 1D). Similarly, OS cell invasion assessed by Transwell assay also showed enhancement after coculturing with exosomes (Fig. 1E). All results proved that exosomes isolated from OS cells could promote OS cell malignancy.
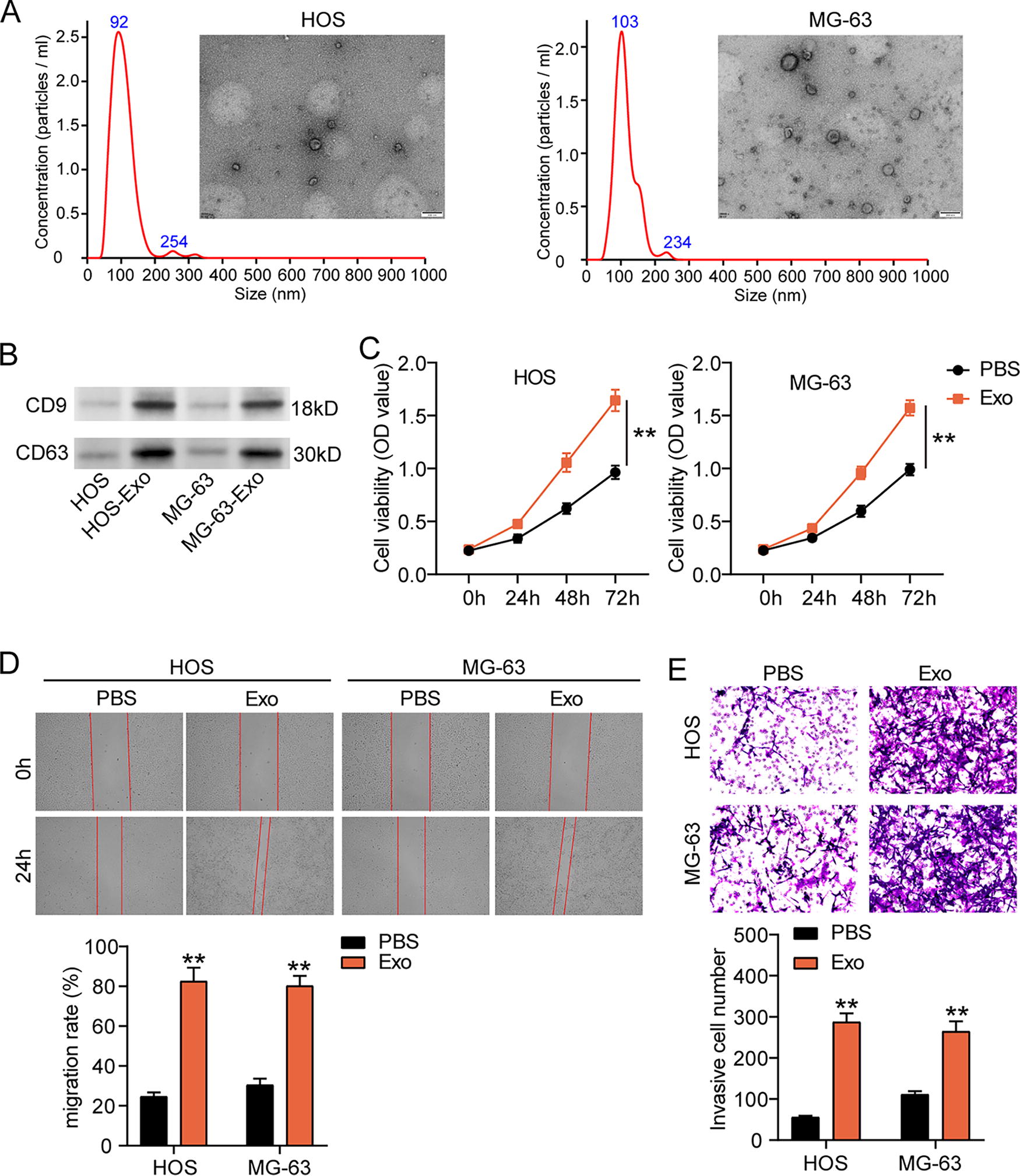
Osteosarcoma (OS) cell-derived exosomes facilitate OS cell malignancy.

Exosomes induce the upregulation of hsa_circ_0000116 in OS.
Hsa_circ_0000116 is upregulated in exosomes from OS
A circRNA microarray GSE218526 included circRNA expression data in plasma exosomes from 10 patients with OS and 5 healthy donors. By GEO2R analysis, the top 10 upregulated circRNAs in exosomes from OS samples were screened (Fig. 2A). Due to hsa_circ_0000116 having the highest expression in exosomes from OS samples, we selected it as our interested circRNA. Then, we detected hsa_circ_0000116 expression in our OS cells cocultured with and without exosomes. qRT-PCR data indicated that hsa_circ_0000116 expression was also upregulated in OS cells cocultured with exosomes (Fig. 2B). Taken together, hsa_circ_0000116 expression could be enhanced in exosomes from OS cells.
Silencing hsa_circ_0000116 suppresses the exosome-induced OS cell malignancy
To explore the function of hsa_circ_0000116 in exosomes, we transfected si-hsa_circ_0000116 to OS cells and further isolated exosomes to coculture with OS cells. CCK8 assay proved that silencing hsa_circ_0000116 relieved the enhancement of OS cell proliferation induced by exosomes (Fig. 3A). Wound healing assay revealed that the elevated migration rate induced by exosomes was declined by silencing hsa_circ_0000116 (Fig. 3B). In Transwell assay, silencing hsa_circ_0000116 reduced the number of invasive cells that was increased by exosomes (Fig. 3C). Hence, silencing hsa_circ_0000116 could effectively inhibit the exosome-induced OS cell malignancy.
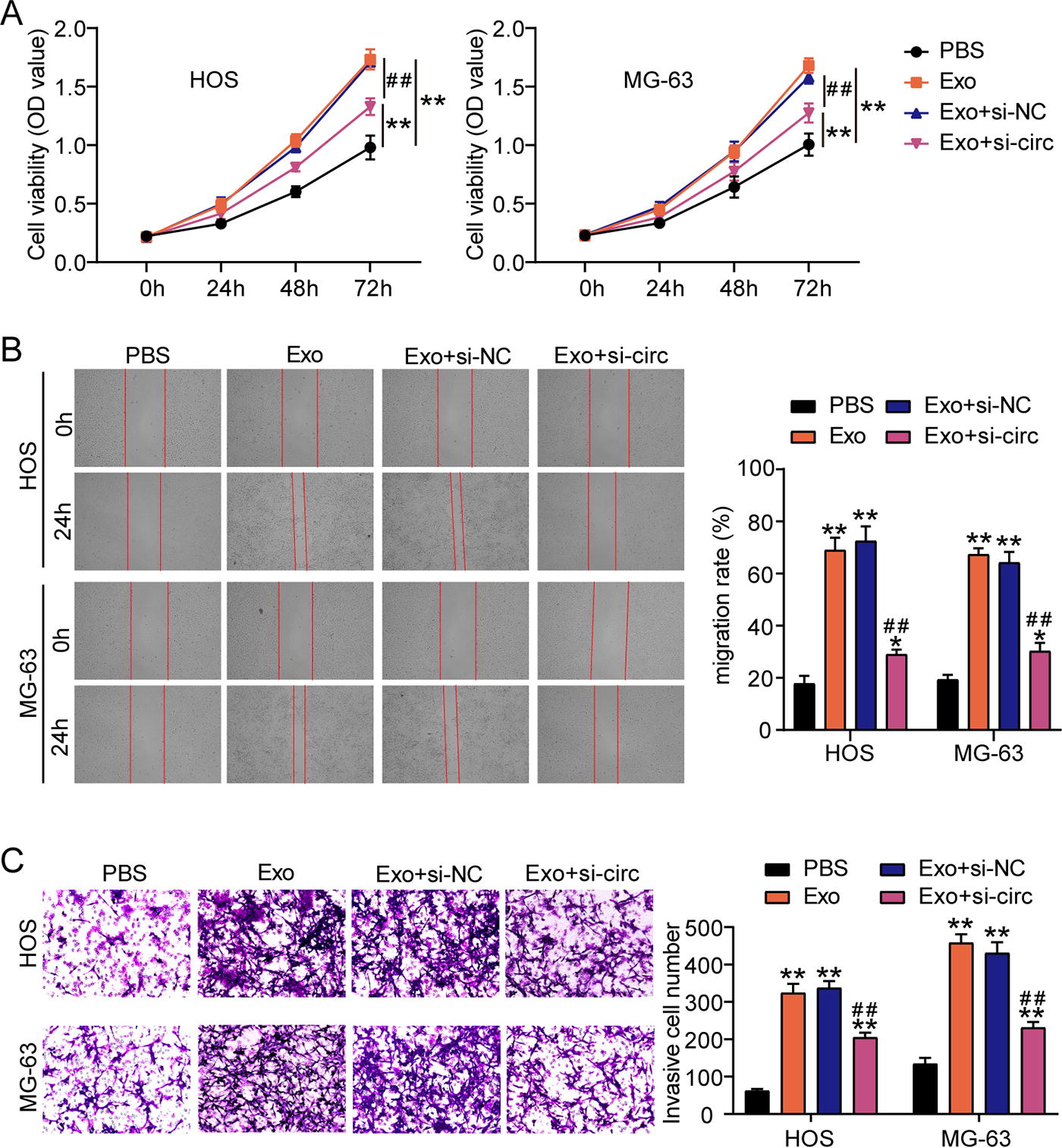
Silencing hsa_circ_0000116 suppresses the exosome-induced OS cell malignancy.
Silencing hsa_circ_0000116 inhibits the activation of PI3K/Akt/mTOR and p38/MAPK pathways induced by exosomes in OS cells
To confirm the downstream signaling pathways of hsa_circ_0000116, western blotting detected the signaling pathway-associated proteins. The results showed that exosomes activated the PI3K/Akt/mTOR pathway by elevating the levels of phosphorylated PI3K, Akt, and mTOR proteins, but silencing hsa_circ_0000116 inhibited the exosome-mediated activation of PI3K/Akt/mTOR pathway owing to reducing the levels of phosphorylated PI3K, Akt, and mTOR proteins (Fig. 4A). In addition, we also found that the phosphorylated p38 expression was enhanced by exosomes and then reduced by silencing hsa_circ_0000116, suggesting that silencing hsa_circ_0000116 could also suppress the activation of p38/MAPK pathway that was induced by exosomes (Fig. 4B). These results confirmed that silencing hsa_circ_0000116 in OS cells could inhibit exosome-induced activation of PI3K/Akt/mTOR and p38/MAPK pathways.
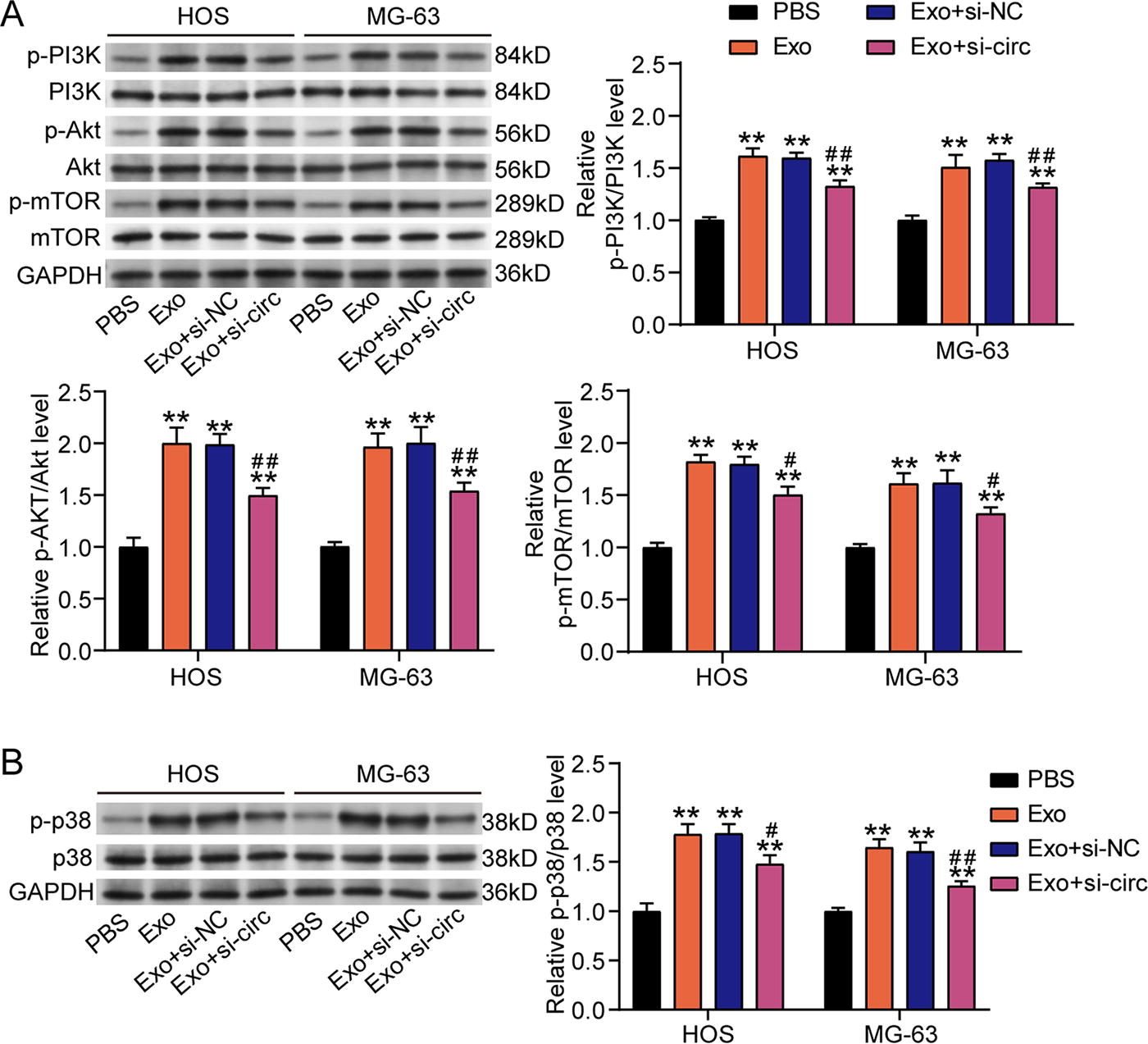
Silencing hsa_circ_0000116 inhibits the activation of PI3K/Akt/mTOR and p38/MAPK pathways induced by exosomes in OS cells.
Discussion
Exosome-delivered circRNAs have been confirmed to play the key roles in regulating tumorigenesis (Chen et al., 2024; Li et al., 2020; Yang et al., 2022). This study investigated the role of exosome-delivered hsa_circ_0000116 in OS progression and its potential molecular mechanisms, focusing on the PI3K/Akt/mTOR and p38/MAPK pathways. Our results demonstrated that exosomes from OS cells accelerated the proliferation, migration, and invasion of OS cells. Notably, hsa_circ_0000116 was upregulated in these OS-derived exosomes, and inhibiting hsa_circ_0000116 relieved exosome-induced cell malignancy in OS by inactivating PI3K/Akt/mTOR and p38/MAPK pathways.
In the context of cancer, exosomes are increasingly recognized as key players in tumorigenesis. In OS, exosomes can promote cancer cell proliferation, migration, and invasion of OS cells, thereby facilitating OS progression (Jiang et al., 2023; Yue et al., 2022; Zhang et al., 2021). Consistent with the previous studies on exosomes in OS, this study also proved that exosomes isolated from OS cells could enhance the abilities of OS cell to proliferate, migration, and invade. However, this study also found the upregulation of a circRNA hsa_circ_0000116 in exosomes from OS samples, which was different from the previous studies. Reviewing the previous studies, only two studies explored that exosomes could transmit circ_0056285 and circ-LMO7 to regulate OS progression (Huo and Dou, 2021; Luo et al., 2024). circ_0056285 promoting OS cell malignancy was reported to be upregulated in serum exosomes of patients with OS (Huo and Dou, 2021). Another study revealed that circ-LMO7 packed in exosomes could attenuate the abilities of OS cell proliferation (Luo et al., 2024). This study confirmed that silencing hsa_circ_0000116 suppressed exosome-induced OS cell malignancy, indicating that hsa_circ_0000116 acts as an oncogenic driver in OS progression. So far, the role of hsa_circ_0000116 has not been reported in any cancer types. Therefore, our study is the first to verify that hsa_circ_0000116 is an oncogenic circRNA to regulate OS cell malignancy.
PI3K/Akt/mTOR and p38/MAPK pathways are well-known for their roles in cell growth, survival, and metastasis, and their aberrant activation has been frequently observed in various cancers, including OS (Hsu et al., 2019; Kang et al., 2018; Liu et al., 2018; Zhang et al., 2022). However, whether circRNA could regulate PI3K/Akt/mTOR and p38/MAPK pathways to affect OS progression is still not fully understood. Here, we found that knockdown of hsa_circ_0000116 reduced the levels of phosphorylated PI3K, Akt, mTOR, and p38 proteins. As we know, the mechanism on the activation of PI3K/Akt/mTOR pathway is that phosphorylated PI3K can recruit and activate Akt, and then, Akt was phosphorylated to activate its downstream effector mTOR, leading to phosphorylation of mTOR. As for the p38/MAPK pathway, once it is activated, it is phosphorylated. Therefore, we could conclude that hsa_circ_0000116 activated the PI3K/Akt/mTOR and p38/MAPK pathways in OS cells.
Despite the significant findings of exosomal hsa_circ_0000116 in OS, this study has several limitations. First, while we demonstrated the role of exosomal hsa_circ_0000116 in promoting OS malignancy, the precise molecular interactions between hsa_circ_0000116 and the specific components of PI3K/Akt/mTOR and p38/MAPK pathways remain unclear. Further mechanistic studies, such as identifying direct binding partners or downstream targets, are necessary to fully elucidate the functional relevance of hsa_circ_0000116 in these pathways. Our experiments were conducted using in vitro cell models, which may not fully recapitulate the complexity of OS progression in vivo. Animal models such as xenograft experiment should be performed in the future to show the role of exosome-delivered hsa_circ_0000116 in OS progression. Clinically, mesenchymal stem cell (MSC) therapy has been reported to prevent cancer progression by inhibiting PI3K/AKT/mTOR pathway, and MSCs are also associated with bone fracture repair (Kangari et al., 2024a; Kangari et al., 2024b). Therefore, further study exploring the correlation between MSCs and hsa_circ_0000116 in OS may provide a new treatment approach for OS. In addition, the clinical significance of hsa_circ_0000116 was not assessed in this study, which should be further explored by collecting the OS clinical samples.
Conclusion
Overall, this study reveals that exosome-delivered hsa_circ_0000116 can facilitate OS cell malignancy by activating the PI3K/Akt/mTOR and p38/MAPK pathways. Our finding suggests that targeting hsa_circ_0000116 may represent a promising therapeutic approach to inhibit OS progression.
Consent for Publication
All participants consented to the publication of this research.
Footnotes
Data Availability Statement
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Disclosure Statement
No conflicts of interest were declared by the authors.
Funding Information
No funding was received for this article.