
Abstract
Liquiritin, a key component extracted from Glycyrrhiza radix, exhibits a variety of physiological effects. This study investigates the role of liquiritin in the progression of breast cancer. This investigation conducted experiments using two breast cancer cell lines treated with varying concentrations of liquiritin, further validating our findings in vivo. Bioinformatics analysis was used to identify the pathways potentially regulated by liquiritin in breast cancer. The results indicated that the epidermal growth factor receptor (EGFR) and mitogen-activated protein kinase 8 (MAPK8) are potential downstream factors regulated by liquiritin in breast cancer. Our findings demonstrated that liquiritin significantly suppressed cell proliferation and induced cell cycle arrest in a dose-dependent manner. In addition, liquiritin triggered apoptosis by inhibiting the phosphatidylinositol-4,5-bisphosphate 3-kinase/protein kinase B signaling pathway. Liquiritin also reduced mitochondrial membrane potential, leading to mitochondrial dysfunction and promoting excessive reactive oxygen species (ROS) production by suppressing the EGFR/MAPK8 signaling pathway. Furthermore, liquiritin treatment resulted in a notable decrease in tumor size in breast cancer models through inhibiting cell proliferation and promoting apoptosis. In conclusion, liquiritin serves as an effective tumor suppressor, suppressing the proliferation and cell cycle progression of breast cancer cells, while inducing apoptosis by regulating mitochondrial function and ROS generation via the EGFR/MAPK8 signaling pathway.
Introduction
Breast cancer is a serious life-threatening disease and is regarded as the most prevalent cancer among women worldwide (Siegel et al., 2023). The incidence and mortality rates of breast cancer remain increasing among women globally (Anastasiadi et al., 2017; Sung et al., 2021). Currently, the primary treatment approaches for breast cancer include surgery, radiotherapy, and chemotherapy, all of which can cause irreversible damage to healthy cells (Arslan et al., 2014; Shi et al., 2016). Furthermore, therapeutic strategies often involve the combination of multiple drugs or targeting various molecular pathways. Natural active ingredients derived from Chinese herbal medicine show promise in preventing breast cancer (Banik et al., 2017). Recent studies have demonstrated that certain components extracted from Chinese herbal medicine can inhibit the carcinogenesis and progression of breast cancer through the epidermal growth factor receptor (EGFR) signaling pathway (Farghadani and Naidu, 2022; Wang et al., 2019). Therefore, it is crucial to explore natural substances that are less toxic and effective against breast cancer cells.
Glycyrrhiza radix (G. radix) is a medicinal plant native to China, known for its diverse pharmacological activities, including antioxidant, anti-inflammatory, and anticancer effects (Nakatani et al., 2017; Wang et al., 2014; Yu et al., 2015). Liquiritin, a flavonoid derivative, is a main bioactive component of G. radix (Cheng et al., 2021) that helps prevent inflammatory responses and cancer progression (Dong et al., 2007; Wang et al., 2008; Yang et al., 2021). A previous study demonstrated that liquiritin induces apoptosis by suppressing mitochondrial membrane potential (MMP), induces cell cycle arrest by regulating p21, p27, cyclin B, and cyclin-dependent kinase (CDK), and triggers reactive oxygen species (ROS) production through the mitogen-activated protein kinase (MAPK)/protein kinase B (AKT) pathway in hepatoma carcinoma cells (Wang et al., 2020). The MAPK pathway generally hyperactivated in diverse types of cancers has been considered to mediate malignant phenotypes in various tumors (Ganesan et al., 2024; Low and Zhang, 2016). The activation of this pathway is an important program to promote multiple processes, such as cell proliferation, metastasis, and survival (Chen et al., 2017). In addition, liquiritin inhibits cell apoptosis by activating caspase-3 and poly ADP-ribose polymerase (PARP) (He et al., 2017). The anticancer effects of liquiritin in gastric cancer have been reported to result from its ability to promote cell apoptosis and ROS generation (Xie et al., 2017). However, the potential antitumor effects of liquiritin in breast cancer have not yet been reported.
We focused on the role and regulatory mechanisms of liquiritin in breast cancer, verifying its effects both in vivo and in vitro. Different concentrations of liquiritin were administered to MDA-MB-231 and MDA-MB-436 cells, which are commonly used to explore the effects of drugs on the development of breast cancer. We found that liquiritin exhibits anticancer effect in breast cancer. Liquiritin inhibits cell proliferation and promotes apoptosis in a dose-dependent manner. Cell cycle arrest, apoptosis, mitochondrial dysfunction, and oxidative stress were induced by liquiritin in breast cancer cells. Based on database analysis and previous literatures, we screened for downstream factors and pathways that may be regulated by liquiritin. The EGFR/MAPK8 signaling pathway was demonstrated to mediate the anticancer effects of liquiritin on breast cancer progression. Our findings provide evidence for the treatment of breast cancer with natural ingredients and open up the possibility of nonsurgical and nonchemotherapeutic approaches to alleviate breast cancer.
Methods and Materials
Data acquisition
Protein targets of liquiritin were predicted using the web tool SwissTargetPrediction (http://swisstargetprediction.ch/index.php). Genes associated with breast cancer were sourced from GeneCards (https://www.genecards.org/). The molecules potentially regulated by liquiritin in breast cancer were identified through Venn analysis and analyzed using Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis. Pathways related to the genesis and progression of breast cancer are presented in bar graph after screening by p < 0.05 and percentage >5%.
Cell culture and treatment
Human breast cancer cell lines, MDA-MB-231 and MDA-MB-436, were incubated in L15 commercial medium (ServiceBio, Wuhan, China) supplemented with 10% fetal bovine serum (Tianhang Biotechnology, Huzhou, China).
The treatment concentrations of liquiritin (Macklin Inc., Shanghai, China) were determined using a Cell Counting Kit-8 (CCK-8) provided by Solarbio (Beijing, China). MDA-MB-231 and MDA-MB-436 cells, planted in 96-well plates, were treated with various doses (0, 25, 50, 100, 150, and 200 μM) of liquiritin for 48 h. Subsequently, 10 μL of CCK-8 solution was added to each well and incubated for 2 h. The optical density values were then measured at 450 nm using an 800 TS microplate reader (BioTek, Winooski, VT, USA). In this study, the treatment concentrations of liquiritin were designed to be approximately 1/8 IC50 (Liquiritin-L), 1/4 IC50 (Liquiritin-M), and 1/2 IC50 (Liquiritin-H). MDA-MB-231 cells were treated with 10, 20, and 40 μM of liquiritin, whereas MDA-MB-436 cells were treated with 5, 10, and 20 μM of liquiritin.
Colony formation assay
Cells (300 per well) were cultured in 60 mm plates for 14 days. Following a wash with phosphate buffer (PBS), cells were fixed with R1 reagent for 1 min. After staining with R2 reagent for 5 min, colonies were photographed using an IX53 microscope (Olympus, Tokyo, Japan).
Cell cycle detection
Cells were collected by centrifugation at 150 g for 5 min. The cell pellet was then fixed with 70% precooled ethanol at 4°C for 2 h. Subsequently, cells were incubated with RNase A for 30 min, followed by incubation with propidium iodide (PI) at 4°C for 30 min in the dark. Cell cycle distribution was then analyzed using a NovoCyte flow cytometer.
Annexin V-FITC/PI staining
Cells were collected by centrifugation, and the supernatant was removed. Cells were then resuspended in 1 mL of precooled PBS to create a cell suspension. One hundred microliter of cell suspension was mixed with 5 μL of Annexin V-FITC and incubated in the dark for 10 min. After adding 10 μL of PI in the mixture for 5 min, flow cytometry was performed immediately.
Western blot
RIPA lysis buffer (Proteintech, Wuhan, China) was used to extract proteins from the cells. Cytoplasmic and mitochondrial proteins were isolated using a mitochondrial isolation and protein extraction kit (Proteintech, Wuhan, China). The concentration of protein samples was determined using a BCA Detection Kit (Proteintech, Wuhan, China). Subsequently, sodium dodecyl sulfate–polyacrylamide gel electrophoresis was performed to separate proteins at 80V for 2.5 h, followed by transferring the proteins to a polyvinylidene fluoride (PVDF) membrane (Thermo Scientific, Pittsburgh, PA, USA) at 85 V for 1.5 h. After blocking the membrane with a blocking buffer (Proteintech, Wuhan, China), the membranes were incubated at 4°C overnight with primary antibodies. Following four washes with washing buffer on a shaker, the PVDF membranes were then incubated with secondary antibodies at 37°C for 40 min. Information on all antibodies used for Western blot assay is listed in Supplementary Table S1. Proteins were visualized using enhanced chemiluminescence (Proteintech, Wuhan, China) reagents. The relative expression of proteins was quantified using blot densitometry with Gel-Pro-Analyzer software.
Hoechst staining
Cells in each group were incubated with Hoechst 33342 staining solution (Biosharp, Hefei, China) for 5 min at room temperature. Cell morphology was imaged under an IX53 fluorescence microscope.
DCFH-DA staining
Cells were harvested by centrifugation, and the supernatant was removed. The cells were incubated with 10 μM of a ROS probe (Biosharp, Hefei, China) in the dark for 30 min at 37°C. After washing by serum-free medium twice, cells were analyzed using a NovoCyte flow cytometer.
JC-1 staining
Breast cancer cells were collected and stained with JC-1 solution (Biosharp, Hefei, China) for 20 min. Following centrifugation, the supernatant was discarded. Cells were washed with JC-1 staining buffer and then resuspended in the staining buffer for analysis using flow cytometry. In addition, cells from each group were mixed with JC-1 staining solution for 20 min. After washing with JC-1 staining buffer, cells were observed under a IX53 fluorescence microscope.
Animal
BALB/c nude mice were obtained from Huachuang Sino Medical Co., LTD (Taizhou, China). A total of 12 female mice aged 6 weeks were housed under standard conditions with free access to food and water. Mice were randomly divided into two groups (n = 6) as follows: Control and Liquiritin. One week after subcutaneous injection of MDA-MB-231 cells (2 × 106) into the mouse mammary fat pad, mice in the liquiritin group were administered with 30 mg/kg/day of liquiritin via intraperitoneal injection. Tumor volume was measured and recorded every 4 days. After 36 days, tumors were excised, photographed, and stored for the subsequent experiments. All animal protocols in the present study were approved by the Ethics Committee in Heilongjiang University of Chinese Medicine (No. 2024042613).
Immunohistochemistry
The collected tumor tissues were fixed, dehydrated, and embedded in paraffin. The paraffin blocks were then sectioned, and the sections were dried. After deparaffinization, sections were incubated with antigen retrieval solution at low temperature for 10 min, followed by a 15-min incubation with 3% hydrogen peroxide (Sinopharm, Shanghai, China). After blocking with bovine serum albumin, sections were incubated with anti-Ki67 (Dilution 1:200, Cat. No. 27309–1-AP, Proteintech, Wuhan, China) at 4°C overnight. Next, sections were treated with horseradish peroxidase-labeled goat anti-rabbit IgG (Dilution 1:100, Cat. No. SE134, Solarbio, Beijing, China) for 45 min. Sections were then incubated with diaminobenzidine solution (Sangon Biotech, Shanghai, China) at 37°C for 10 min. After counterstaining with hematoxylin (Solarbio, Beijing, China) for 3 min, sections were treated with ethanol hydrochloride for 3 s and immediately rinsed with running water. Finally, sections were imaged using a BX53 microscope.
Terminal deoxynucleotidyl transferase-mediated dUTP Nick-End labeling assay
Following deparaffinization, tumor sections were treated with 0.1% Triton X-100 (Beyotime, Shanghai, China); terminal deoxynucleotidyl transferase-mediated dUTP Nick-End labeling (TUNEL) staining solution (Roche, Basel, Switzerland) was then applied to the sections and incubated in the dark for 60 min. Subsequently, tissues were counterstained with 4’,6-diamidino-2-phenylindole (Aladdin, Shanghai, China) in the dark. After mounting with an antifading mounting medium (Solarbio, Beijing, China), sections were imaged using a BX53 microscope.
Statistical analysis
Statistical analysis was performed using GraphPad Prism (version 8.0). Differences among groups were analyzed using a one-way analysis of variance followed by the Tukey’s multiple comparisons test. Statistical significance was defined as p < 0.05.
Results
Downstream factors potentially affected by liquiritin in breast cancer progression were screened
The chemical structure of liquiritin is depicted in Figure 1A. A total of 91 overlapping genes were identified to be associated with breast cancer and were considered potential downstream targets of liquiritin (Fig. 1B). As shown in Figure 1C, these genes were enriched in pathways implicated in tumor progression, suggesting that liquiritin may modulate breast cancer development through these pathways. The Sankey diagram illustrated key factors involved in the Ras signaling pathway, chemical carcinogenesis-ROS production, apoptosis, and metabolic pathways (Fig. 1D). Collectively, our findings suggest that liquiritin potentially regulates breast cancer progression via the EGFR/MAPK8 signaling pathway.
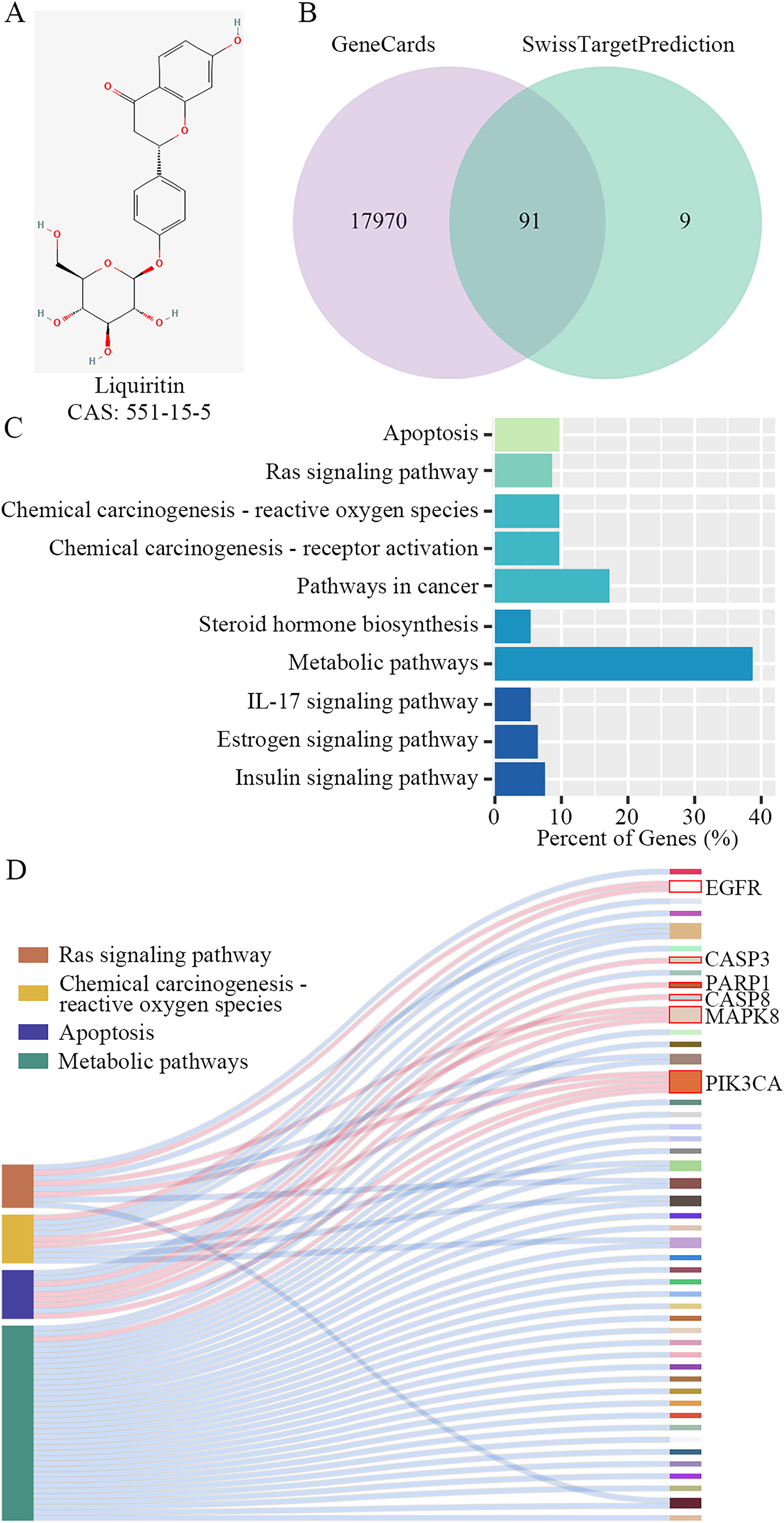
Downstream factors potentially affected by liquiritin in breast cancer progression were screened.
Liquiritin inhibits cell cycle progression and suppresses proliferation in breast cancer cells
Based on the observed inhibition rate, we calculated the IC50 values of liquiritin in two breast cancer cell lines (Fig. 2A). Subsequently, liquiritin was used at concentrations of 10, 20, and 40 μM and 5, 10, and 20 μM for subsequent experiments, respectively. The colony formation assay demonstrated that the proliferation of cancer cells was significantly suppressed by increasing concentrations of liquiritin (Fig. 2B). Compared with the control group, liquiritin treatment resulted in a significant increase in cells in the G1 phase and a concomitant decrease in cells in the S phase (Fig. 2C). The CDK inhibitors p21 and p27 are crucial regulators of cell cycle arrest in numerous human malignancies (Abukhdeir and Park, 2008). We observed a significant upregulation of p27 and p21 expressions in cells treated with increasing doses of liquiritin. Furthermore, the expression of Cyclin B1 and CDK1 was downregulated in liquiritin-treated cells (Fig. 2D). These findings collectively confirm that liquiritin inhibits proliferation and induces cell cycle arrest in breast cancer cells in a dose-dependent manner.
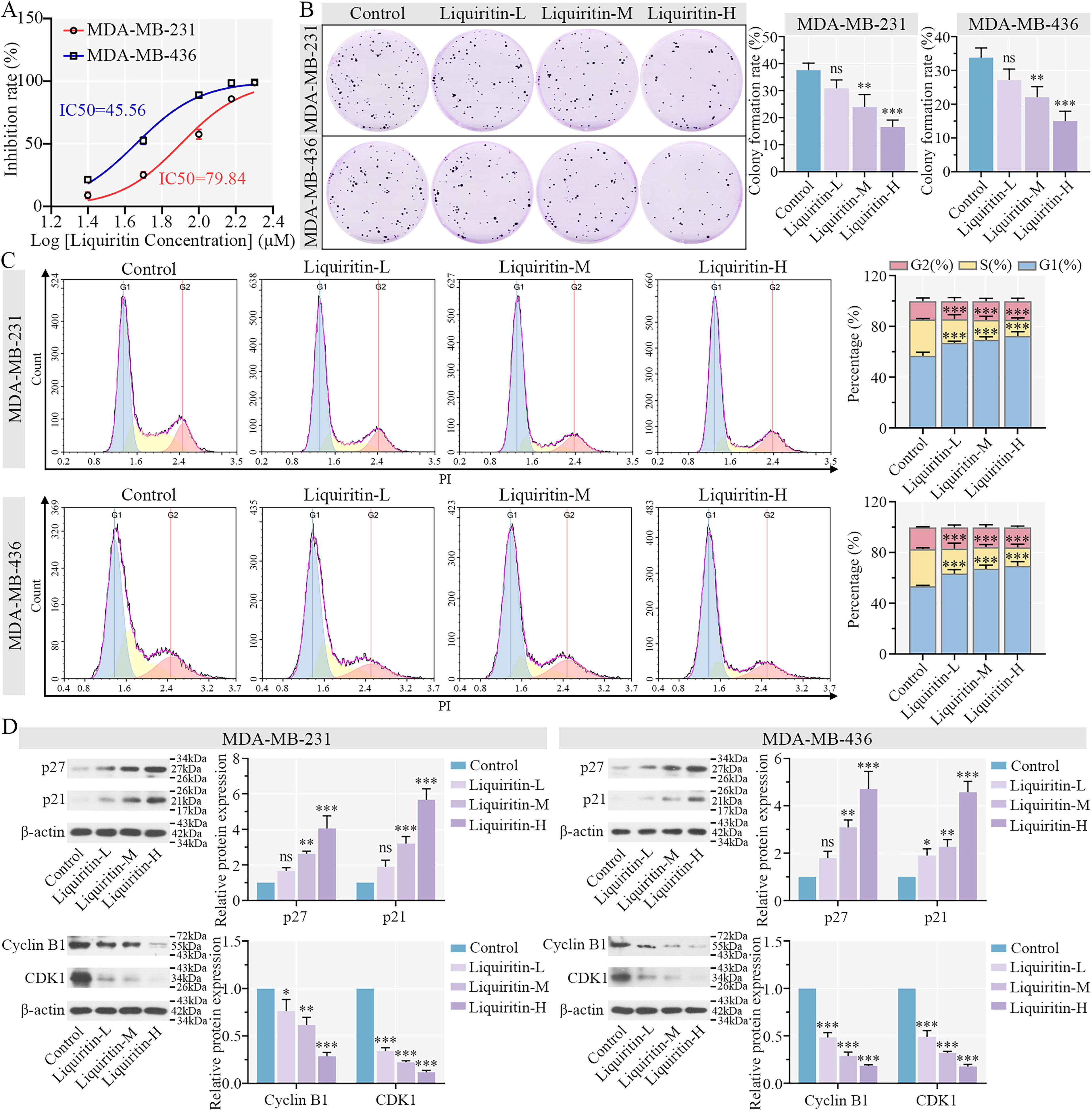
Liquiritin inhibits cell cycle progression and suppresses proliferation in breast cancer cells.
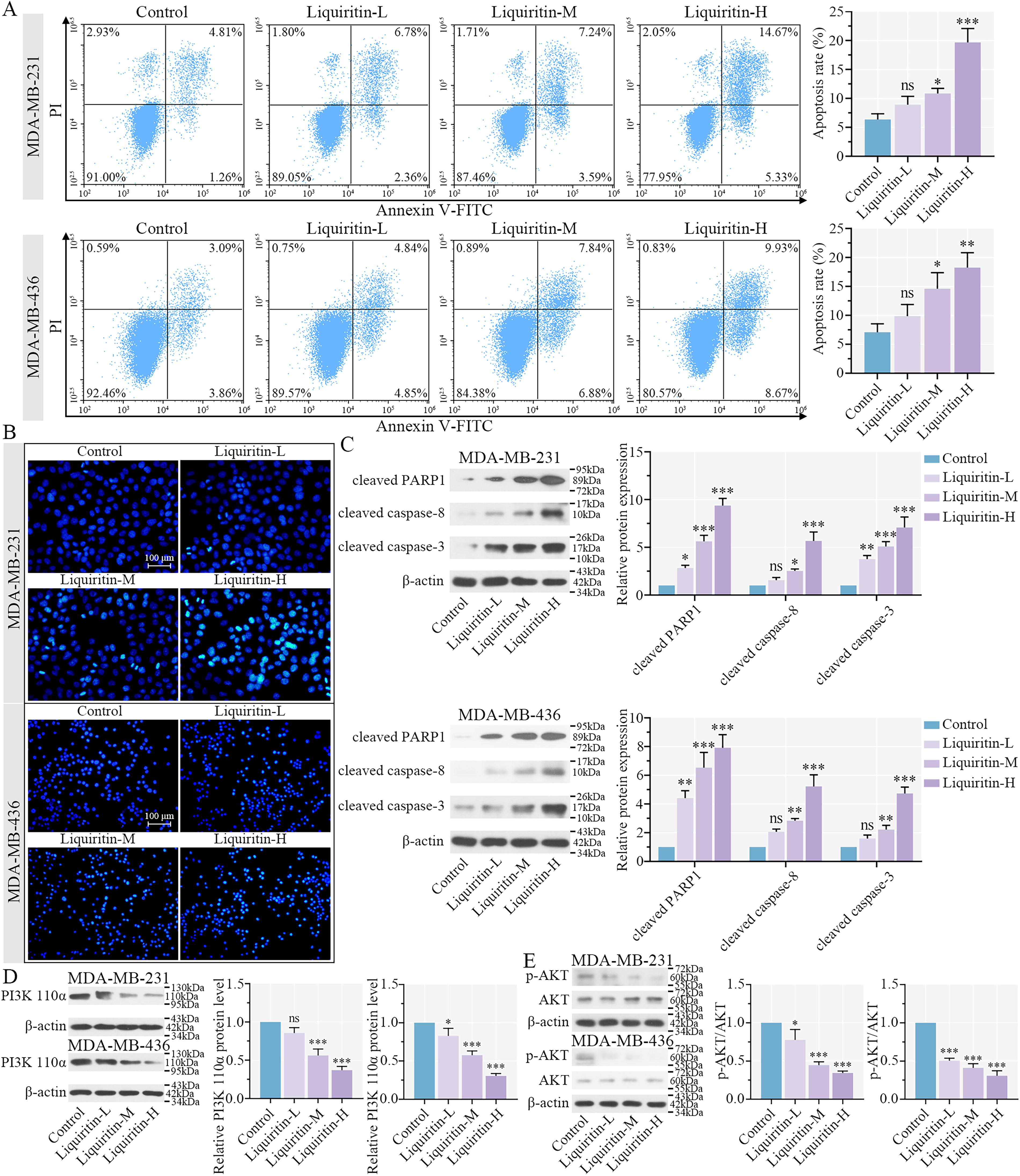
Liquiritin induces cell apoptosis of breast cancer cells through the PI3K/AKT signaling pathway.
Liquiritin induces apoptosis of breast cancer cells through the PI3K/AKT signaling pathway
Annexin V/PI staining revealed that treatment with medium and high concentrations of liquiritin (Liquiritin-M and -H, respectively) significantly enhanced the apoptosis rate, whereas low-dose liquiritin (Liquiritin-L) induced a modest increase of apoptosis in the two cell lines (Fig. 4A). Nuclei were stained with Hoechst solution, and changes in nuclear morphology and fluorescence intensity were assessed to identify apoptotic cells. We observed an increase in bright nuclear staining following liquiritin treatment (Fig. 4B), suggesting that liquiritin promotes apoptosis in breast cancer cells. Furthermore, the relative protein expression levels of cleaved PARP1, cleaved caspase-3, and cleaved caspase-8 were increased by liquiritin in both breast cancer cells (Fig. 4C). These results suggested that liquiritin activated PARP1, caspase-3, and caspase-8 to induce apoptosis. Abnormal activation of the phosphatidylinositol-4,5-bisphosphate 3-kinase (PI3K)/AKT signaling pathway is frequently observed in malignant tumors, including breast cancer (Cerma et al., 2023). Based on the KEGG enrichment analysis, phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit alpha, encoding PI3K 110α, was identified as a potential downstream target of liquiritin. Liquiritin treatment reduced the protein expression of PI3K 110α in the two cell lines (Fig. 4D). Concurrently, the phosphorylation of AKT was decreased by liquiritin, indicating that liquiritin suppressed activation of the PI3K/AKT signaling pathway in breast cancer. In summary, liquiritin promotes apoptosis through upregulating PARP1, caspase-3, and caspase-8 expression and by inhibiting the PI3K/AKT signaling pathway in breast cancer cells.
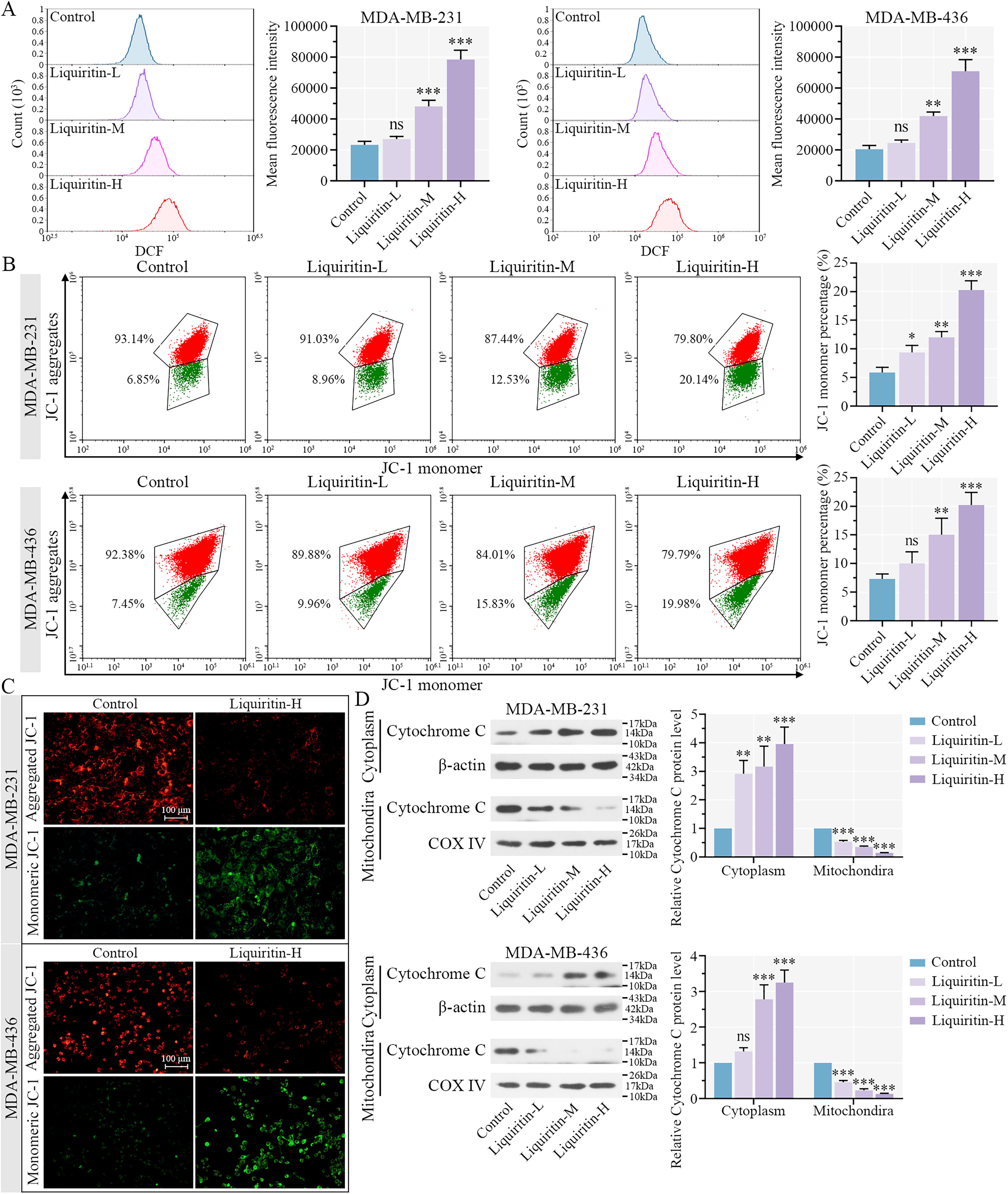
Liquiritin enhances mitochondrial dysfunction and ROS generation of breast cancer cells.
Liquiritin enhances mitochondrial dysfunction and ROS generation via the EGFR/MAPK8 signaling pathway
As shown in Figure 5A, the mean fluorescence intensity was significantly increased in the Liquiritin-M and -H groups compared with the control, which confirmed that liquiritin induced the generation of ROS. A decline in MMP is considered an early indicator of apoptosis (Marchetti et al., 1996). We observed an increase in JC-1 monomers and a decrease in aggregates in both cell lines treated with varying concentrations of liquiritin (Fig. 5B). After liquiritin treatment, the intensity of red fluorescence was reduced, whereas green fluorescence was enhanced in both breast cancer cell lines (Fig. 5C). These results indicated that liquiritin induced a reduction in MMP and promotes ROS generation. The protein expression of cytochrome C was upregulated in the cytoplasm and downregulated in the mitochondria by liquiritin, suggesting that liquiritin triggered the translocation of cytochrome C in breast cancer cells. Thus, liquiritin induces ROS production and MMP reduction, promoting apoptosis in breast cancer cells.
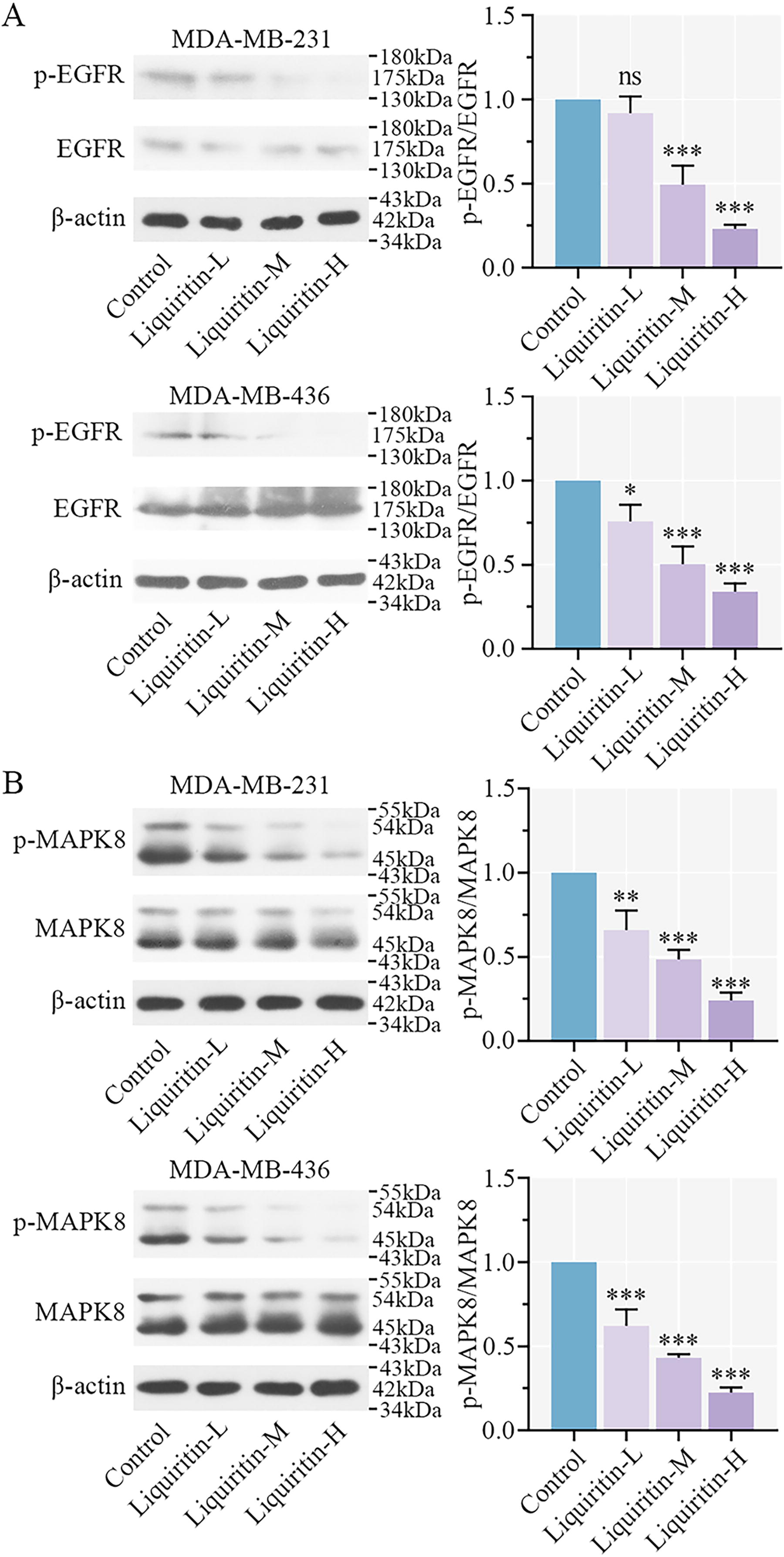
Liquiritin blocks the activation of EGFR/MAPK8 signaling pathway.
EGFR activation is known to contribute to the proliferation and growth of cancer cells (Zandi et al., 2007). As shown in Figure 6A, liquiritin treatment suppressed EGFR phosphorylation in a dose-dependent manner (Fig. 6A). MAPK8 is involved in various cellular events, including apoptosis and oxidative stress. Phosphorylation of MAPK8 is required for cell apoptosis (Li et al., 2015). Following treatment with different concentrations of liquiritin, the phosphorylation of MAPK8 was decreased in cancer cells (Fig. 6B). These findings confirm that liquiritin facilitates oxidative stress and mitochondrial dysfunction by inhibiting the EGFR/MAPK8 signaling pathway.
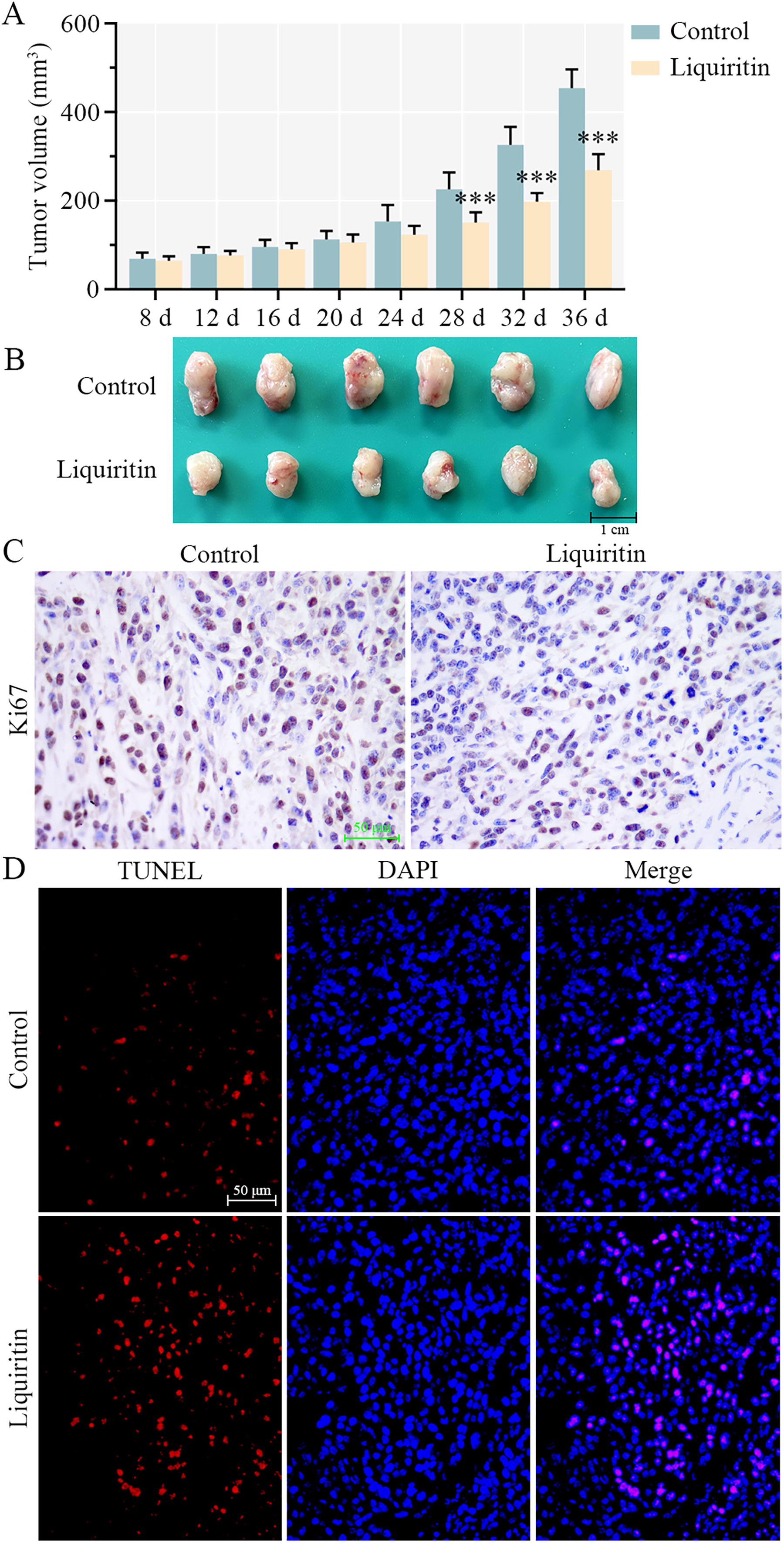
Liquiritin inhibits tumor growth of breast cancer in vivo.
Liquiritin inhibits tumor growth of breast cancer in vivo
Liquiritin has been elucidated to inhibit proliferation and cell cycle progression in vitro and to induce ROS generation and apoptosis. Subsequently, the role of liquiritin in vivo was investigated. One week after MDA-MB-231 cell transplantation, xenograft animals were injected with 30 mg/kg/day of liquiritin for 4 weeks in vivo. Compared with the tumor volume in the control group, liquiritin treatment significantly reduced tumor volume at days 28, 32, and 36 (Fig. 7A). The morphology of tumors is presented in Figure 7B. Tumor size was remarkably decreased by liquiritin administration. In addition, liquiritin treatment suppressed Ki67 expression in tumor tissues (Fig. 7C). Images in Figure 7D demonstrated that the number of TUNEL-positive cells was increased in the tumor tissues of mice receiving liquiritin, suggesting that liquiritin induced apoptosis in vivo. In conclusion, liquiritin effectively suppressed breast cancer tumor growth by affecting both proliferation and apoptosis.
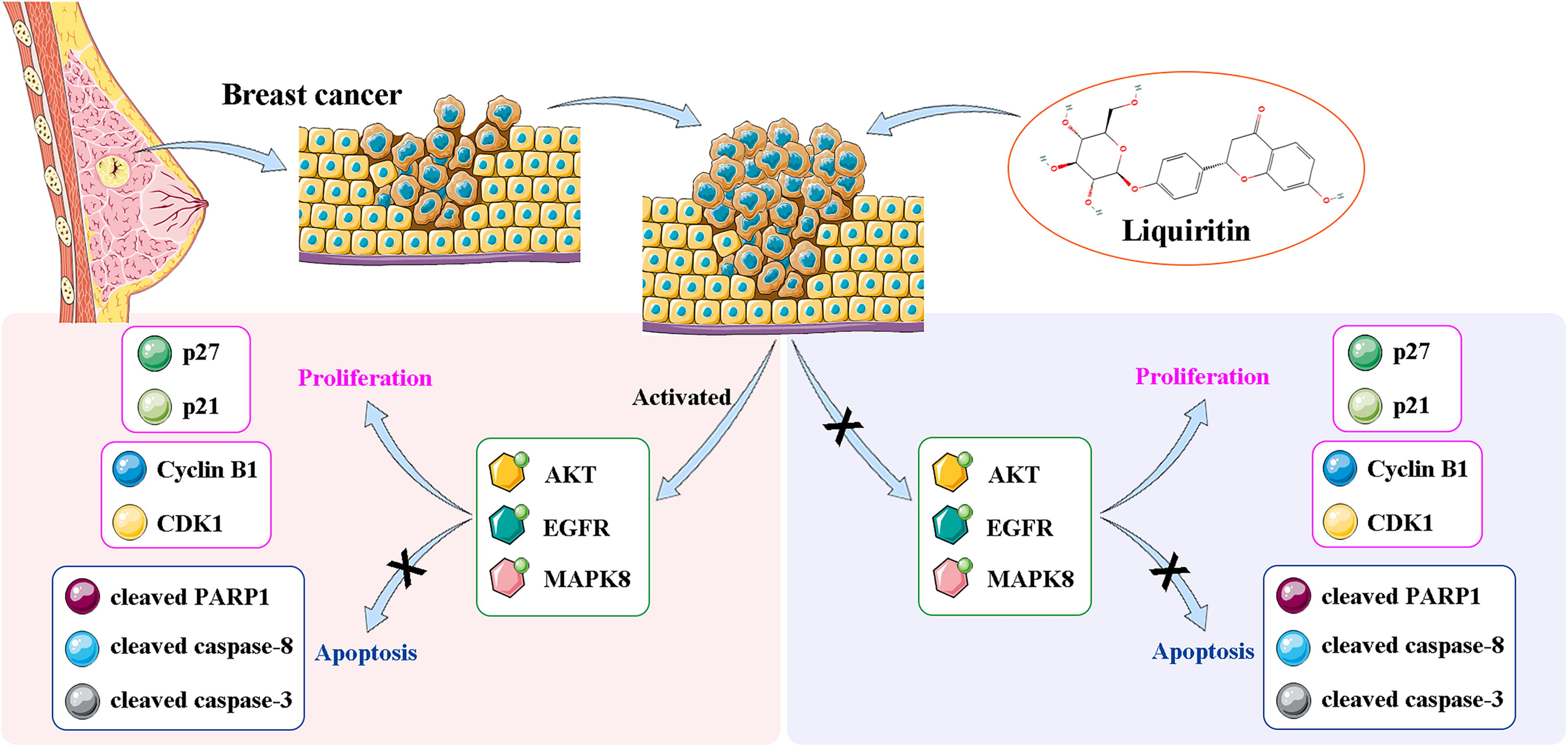
The diagram of liquiritin regulating breast cancer progression. In this study, liquiritin suppresses the cell proliferation and facilitates apoptosis through inhibiting the PI3K/AKT and EGFR/MAPK8 signaling pathways in breast cancer cells, accompanied by the downregulation of Cyclin B1 and CDK1, as well as the upregulation of p27, p21, cleaved PARP1, cleaved caspase-3, and cleaved caspase-8. PI3K, phosphatidylinositol-4,5-bisphosphate 3-kinase; AKT, protein kinase B; EGFR, epidermal growth factor receptor; MAPK8, mitogen-activated protein kinase 8; CDK1, cyclin-dependent kinase 1; PARP1, poly ADP-ribose polymerase 1.
Discussion
Despite improvements in the prognosis of breast cancer due to advancements in therapeutic approaches, the efficiency of cancer cell suppression and clearance still requires enhancement (Dastjerd et al., 2022). Numerous studies have highlighted that liquiritin, an effective and available component derived from G. radix, plays a crucial role in various cancers (He et al., 2017; Wei et al., 2017; Xie et al., 2017). Results from this study indicate that liquiritin suppresses cell proliferation and facilitates apoptosis by leading to cell cycle arrest, mitochondrial dysfunction, and ROS generation via the EGFR/MAPK8 signaling pathway in breast cancer.
The development of new drugs and therapies for cancer prevention and treatment is the key direction of global cancer research. However, the depth and breadth of basic research is still insufficient. The present study provides a new approach for the intervention and remission of tumors by validating the anticancer effect of liquiritin extracted from a natural plant. In this study, liquiritin has been confirmed to obstruct proliferation and contribute to cell cycle arrest in breast cancer via activating p27 and p21, while decreasing cyclin B1 and CDK1. A previous report has confirmed that flavonoids extracted from G. radix exhibits anti-inflammatory and anticancer effects. The flavonoids significantly inhibit the tumorigenic ability of breast cancer cell MDA-MB-231 (Jiang et al., 2018). Liquiritin, one of these flavonoids, has been indicated as a tumor suppressor to suppress the progression of breast cancer and induce apoptosis and oxidative stress in breast cancer. Our findings further validated the conclusion in the previous report. In addition, Zhou et al. have reported that liquiritin, isoliquiritin, and isoliquirigenin are crucial active components from G. radix. These components induced apoptosis through increasing p21 expression, inhibiting the AKT signaling pathway and arresting cell cycle in nonsmall lung cancer cells (Zhou and Ho, 2014). The anticancer effect of liquiritin has been demonstrated in lung cancer cells. Our study confirmed that liquiritin led to an increase in p21 and p27 and a decrease in cyclin B1 and CDK1 in breast cancer cells, as well as inhibiting cell cycle. Meanwhile, liquiritin upregulated the expressions of cleaved PARP1, cleaved caspase-3, and cleaved caspase-8, while also inducing the deactivation of the PI3K/AKT signaling pathway. Our findings further indicated that liquiritin as an effective tumor suppressor impedes proliferation and triggers apoptosis in breast cancer.
According to the results of KEGG enrichment analysis, PARP1, caspase-3, and caspase-8 in apoptosis pathway are potential downstream factors of liquiritin. Liquiritin has been reported to induce apoptosis by activating caspase-3 and PARP in human cervical cancer (He et al., 2017). The activation of caspase-3 and caspase-8 by liquiritin in gastric cancer indicates that liquiritin is an effective drug that activates apoptosis in gastric cancer cells (Xie et al., 2017). The liquiritin-induced upregulation of PARP1, caspase-3, and caspase-8 demonstrates that liquiritin significantly promotes apoptosis in breast cancer cells. Therefore, liquiritin is considered to have anticancer effects on human cancer progression. Our findings show that liquiritin suppresses PI3K 110α expression and the phosphorylation of AKT in two breast cancer cell lines, enhancing cell apoptosis via the PI3K/AKT pathway. Thus, it may serve as a tumor inhibitor with therapeutic effect on breast cancer.
Moreover, liquiritin induced mitochondrial dysfunction by upregulating ROS generation and downregulating MMP. It also inhibited the activation of cytochrome C. The activation of the EGFR/MAPK8 signaling pathway was suppressed by liquiritin, contributing to apoptosis and mitochondrial dysfunction. EGFR is generally upregulated in breast cancer and other cancers (Hagan et al., 2023). Previous studies have indicated that suppressing the EGFR signaling pathway induces cell apoptosis and mediates the anticancer effects of various drugs (Palanivel et al., 2020). The MAPK signaling pathway has been regarded as a key pathway for regulating the progression of breast cancer (Wu et al., 2024). The activation of the MAPK8 signaling pathway mediates the promotive effects of noncoding RNA molecules on the progression of colorectal cancer (Liu et al., 2022). Liquiritin has been reported to exert an antimyocardial fibrosis effect by inhibiting the MAPK8 signaling pathway (Zhang et al., 2016). It also accelerates apoptosis and induces MMP reduction by suppressing the phosphorylation of MAPK8 (Zhai et al., 2019). In addition, the MAPK pathway mediated the inhibitory effect of ononin on tumor bone metastasis in breast cancer (Ganesan et al., 2024). The activation of MAPK signaling pathway serves as an important event to support various cellular processes, including tumor growth and survival (Chen et al., 2017; Engelman, 2009; Machado et al., 2016). Our study has confirmed that liquiritin significantly suppressed the phosphorylation of the MAPK signaling pathway in a dose-dependent manner in breast cancer cells, further suggesting that the MAPK8 signaling pathway is involved in the inhibition of liquiritin on breast cancer progression. Collectively, liquiritin has been indicated to suppress cell proliferation and promote cell apoptosis and oxidative stress in breast cancer through the suppression of the EGFR/MAPK8 signaling pathway.
However, this study is limited by the fact that in vitro studies using cell lines cannot fully mimic the complex tumor microenvironment in vivo. Clinical application of the results of this study will require validation in multiple models. In the future, it is necessary to further study the anticancer properties of liquiritin and explore whether it can be combined with existing anticancer methods and adjuvant therapy to improve the poor prognosis of breast cancer. We will further contribute to the improvement of poor prognosis of breast cancer to broaden the clinical treatment options.
Conclusion
To sum up, liquiritin has been confirmed to be a tumor suppressor contributing to the improvement and therapy of breast cancer. Liquiritin inhibits proliferation through inducing cell cycle arrest in breast cancer and facilitates apoptosis through the PI3K/AKT signaling pathway. The loss of MMP and ROS generation is triggered by liquiritin, indicating that liquiritin promotes oxidative stress and mitochondrial dysfunction to suppress breast cancer development. The inhibitory effect of liquiritin on breast cancer progression was mediated by the EGFR/MAPK8 signaling pathway. Liquiritin is expected to be an anticancer candidate against the development of breast cancer.
Footnotes
Authors’ Contributions
P.L.: Conceptualization, data curation, investigation, methodology, validation, writing—original draft, and writing—review and editing. L.Y.: Data curation, investigation, and validation. Y.J.: Data curation, investigation, and validation. Y.C.: Investigation and visualization. M.Z.: Investigation and visualization. L.J.: Conceptualization, methodology, and writing—review and editing. P.G.: Conceptualization, methodology, and writing—review and editing.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author on special request.
Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This research was supported by the National Natural Science Foundation of China (No. 82304793), the Postdoctoral Foundation of Heilongjiang Province (No. LBH-Z20200), the Joint Guidance Project of Heilongjiang Natural Science Foundation (No. LH2023H072), and the Scientific Research Project of Chinese Medicine of Heilongjiang Province (No. ZHY2023-033).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.