
Abstract
Sepsis is a serious systemic inflammatory condition triggered by a variety of pathogens, including bacteria and viruses, that can result in multiple organ failure and a life-threatening situation. Despite advances in medical care, the mortality rate for sepsis remains high even with aggressive treatment strategies such as antibiotic therapy, fluid resuscitation, and respiratory and circulatory support. Extracellular vesicles (EVs), as a novel nanoscale biocarrier, exhibit diverse biological functions including immune modulation and tissue regeneration, suggesting promising applications in the field. This article provides an overview of the diverse therapeutic effects of EVs derived from various sources in the management of sepsis. Furthermore, EVs not only possess intrinsic therapeutic properties, such as immune modulation, but also function as targeted delivery vehicles for a variety of drug molecules, leading to synergistic therapeutic outcomes. In conclusion, extracellular vesicle therapy is poised to emerge as a dynamic and innovative force driving advancements in sepsis treatment.
Introduction
Sepsis is a severe pathological response to infection that can result in life-threatening organ dysfunction (Schlapbach et al., 2024). The pathogenesis of sepsis involves dysregulation of the body’s response and an overwhelming release of inflammatory mediators. Common sources of infection leading to sepsis include the respiratory, urinary, and abdominal tracts (Karalapillai et al., 2020). Initial clinical symptoms may include fever, chills, and systemic symptoms, which can rapidly progress to hemodynamic instability and organ failure. The incidence of sepsis has been observed to increase annually. Sepsis and septic shock represent significant health care challenges, impacting a substantial number of individuals globally each year, with mortality rates ranging from one in three to one in six (Rudd et al., 2020). These conditions are notably more prevalent among the elderly, potentially due to age-related comorbidities and compromised immune function. Additionally, the incidence of sepsis is influenced by the quality of medical care (Gabrielli et al., 2024). Numerous studies have indicated that immune dysfunction plays a crucial role in the pathogenesis of sepsis (Robey et al., 2024), leading to a complex interplay between hyperinflammatory responses and immune suppression in affected patients. The former is intricately linked to initial tissue injury and organ dysfunction, while profound and prolonged immunosuppression can exacerbate a range of life-threatening complications. This significantly heightens mortality rates during the intermediate and advanced stages of sepsis (Pei et al., 2022). These challenging immune status alterations also complicate the management of sepsis. Sepsis imposes substantial burdens on both society and health care systems, as patients with sepsis experience prolonged hospitalizations, necessitate costly intensive care, and frequently require ongoing treatment for enduring complications (Paoli et al., 2018; Prescott et al., 2015). The escalating prevalence and fatality rates of sepsis pose significant risks for severe organ dysfunction and metabolic disturbances, placing substantial burdens on both society and the health care system. Consequently, the development of efficacious treatment strategies for sepsis is imperative in enhancing patient outcomes and diminishing mortality rates.
Extracellular vesicles (EVs) are vesicle carriers released by various cell types into bodily fluids, typically ranging in diameter from 30 to 1000 nm (D’Souza-Schorey and Schorey, 2018). There are three primary classifications of EVs: exosomes, microvesicles, and apoptotic bodies (van Niel et al., 2018). Exosomes, originating from the endoplasmic reticulum, measure approximately 30–150 nm and are produced through endosomal fusion within polycysts before being released by exocytosis (van Niel et al., 2018). Microvesicles are formed from the plasma membrane and typically range from 100 to 1000 nm in size, developing through outward budding processes (Xu et al., 2022). Apoptotic bodies, which range in diameter from 50 to 5000 nm, are generated from cell fragments during apoptosis (Wei et al., 2022). EVs contain a variety of bioactive substances, including proteins, lipids, microRNAs (miRs), and DNA, which facilitate intercellular communication and information exchange (Choi et al., 2020; Wei et al., 2022).
The functionality of EVs depends on their cellular origin and specific vesicle composition, particularly concerning immune responses. EVs exhibit a range of effects from immune activation to inhibition, contingent on their parental cell origin. For instance, exosomes derived from neutrophils enhance inflammatory cytokine expression through miR transduction (Ye et al., 2023). Exosomes from bacteria act as pathogen-associated molecular patterns, mediating host-pathogen interactions, and hold potential as drug delivery systems and vaccine candidates (Laakmann et al., 2023). Conversely, mesenchymal stem cell-derived EVs (MSC-EVs) possess anti-inflammatory properties and promote tissue repair and regeneration, thus protecting tissues and organs (Qin and Zhao, 2020; Zheng et al., 2019). Endothelial cell-derived exosomes exhibit both anti-inflammatory and endothelial-protective properties, protecting against inflammation and maintaining endothelial cell integrity (He et al., 2020). Exosomes from M2 macrophages and tumor cells display immunosuppressive characteristics, potentially moderating excessive immune responses (Chak et al., 2024; Sun et al., 2022). Moreover, exosomes from other sources, such as platelets and fibroblasts, also show therapeutic potential against sepsis, demonstrating the diverse roles of exosomes in addressing this complex condition (Konig et al., 2021; Kunz et al., 2017). Apoptotic bodies primarily act as immunomodulators in vivo (Wen et al., 2023), predominantly exerting their effects by reducing neutrophil activity (Ou et al., 2022). Utilizing these capabilities, EVs are promising as potential therapeutic interventions for conditions such as septic lung injury and kidney injury (Qiu et al., 2021). Additionally, drug incorporation into EVs enhances therapeutic efficacy and specificity compared with their free form (Gangadaran et al., 2018). The surface of EVs can be engineered with specific targeting peptides to facilitate organ- or cell-specific delivery, improving therapeutic outcomes and reducing adverse effects (Zhang et al., 2022b). Furthermore, EVs can carry pathogen antigens and stimulate immune responses, presenting a promising avenue for therapeutic applications (Zhang et al., 2022a).
This article reviews the roles of EVs derived from various cell sources, such as mesenchymal stem cells, immune cells, tumor cells, and plant cells, in managing sepsis and its molecular pathways. By elucidating the molecular mechanisms that underlie the therapeutic effects of EVs on sepsis, this research contributes to the development of targeted EV engineering strategies. The findings of this study provide both theoretical and empirical support for using EVs as a novel therapeutic agent for precise sepsis treatment. Consequently, further investigation into the mechanisms of EV action and the advancement of their clinical application in sepsis treatment is imperative.
MSC-derived EVs
MSC-EVs play crucial roles in anti-inflammatory responses and tissue regeneration, demonstrating significant therapeutic potential in managing sepsis and various other diseases. This potential is attributed to the proteins and miRs carried by the vesicles, which regulate target cells (Dos Santos et al., 2024; Lopes-Pacheco et al., 2020) (Fig. 1). Specifically, MSC-EVs have been shown to suppress inflammatory cytokine production and facilitate the differentiation of M2-type macrophages, thereby exerting anti-inflammatory effects and promoting tissue repair (Hao et al., 2019; Phinney et al., 2015; Song et al., 2017). Concurrently, MSC-EVs serve to safeguard organs through the mitigation of vascular permeability and the suppression of apoptosis (Li et al., 2019). The strategic utilization of MSC-EVs holds promise for the prophylaxis and management of sepsis.

Exosome secretion and structure (By Figdraw. ID: TYWOI4dbb6).
Sepsis is characterized by a systemic inflammatory response syndrome resulting from infection (Rola et al., 2022). MSCs induce granulocyte trained immunity by secreting parasecretory factors, thereby effecting lasting functional changes by reprogramming hematopoietic stem cells to enhance neutrophil-mediated antimicrobial immunity (Ng et al., 2023). In the immunopathological progression of sepsis, MSC-EVs are identified as key modulators of the immune response by regulating various immune cell populations (Hwang et al., 2023) (Fig. 2). Specifically, MSC-EVs induce a shift in macrophage polarization from the pro-inflammatory M1 phenotype to the anti-inflammatory M2 phenotype. This shift is accompanied by a decrease in the production of pro-inflammatory cytokines, such as tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β), mediated by signaling pathways including nuclear factor kappa-B (NF-κB), serine/threonine kinase, and heme oxygenase-1 (HO-1), and an increase in the secretion of anti-inflammatory factors such as IL-10 (Pei et al., 2024; Ti et al., 2015; Xu et al., 2019; Zhao et al., 2022). The regulatory effects of MSC-EVs on macrophage polarization and cytokine production are closely associated with the composition of microRNAs within the MSC-EVs. For example, adipose tissue-derived MSC-EVs (ADSCs) reduce the polarization of macrophages toward the M1 type by modulating the Notch/miR148a-3p signaling axis, thereby reducing lipopolysaccharids (LPS)-induced inflammation and sepsis (Bai et al., 2020). Increased miR-21 content in exosomes released by MSCs induced by IL-1β preconditioning induced M2-like polarization of macrophages in mouse septicemia models. These EVs were more effective in reducing sepsis symptoms and improving the survival rate of mice. miR-21 inhibits macrophage polarization and sepsis in mice by inhibiting its target gene programmed cell death factor 4 (Yao et al., 2021). EVs containing miR-17 released by bone marrow-derived MSCs (BMSCs) have been shown to regulate bromodomain-containing protein 4, suppress LPS-induced inflammation in RAW264.7 cells via the bromodomain and extraterminal domain4/enhancer of zeste homolog/TNF-related apoptosis inducing ligand pathway, enhance cell viability, and demonstrate therapeutic potential in animal models (Su et al., 2021; Yao et al., 2021). Concurrently, the administration of small EVs derived from MSC-EVs through inhalation has been found to enhance the polarization of macrophages towards the M2 phenotype, exert anti-inflammatory and antioxidant effects by modulating the nuclear factor erythroid-derived 2-like 2 (Nrf2) pathway (Zhao et al., 2022), and inhibit cytokine production through mechanisms such as mitochondrial transfer and oxidative phosphorylation (Morrison et al., 2017). MSC-EVs significantly suppressed cytokine release into the systemic circulation and the infiltration of neutrophils and monocytes in the abdominal cavity (Park et al., 2019). ADSC-EVs facilitate M2 polarization of macrophages through the regulation of the long noncoding RNA (lncRNA) deleted in lymphocytic leukemia 2(DLEU2)miR-106a-5p/latexin axis, thereby attenuating inflammatory responses and decreasing apoptosis of lung-injured cells (He et al., 2024). ADSC-EVs exhibit a protective effect in sepsis-induced acute lung injury by delivering miR-150-5p. EVs transport miR-150-5p to macrophages and inhibit the expression of high mobility group protein 2, thereby inhibiting the mitogen-activated protein kinase (MAPK) pathway, promoting M2 polarization, and inhibiting pro-inflammatory cytokines (Zhao et al., 2024). In addition, the production of EVs important for inflammation resolution was significantly reduced in Syndecan 2-silenced MSCs (Han et al., 2022). BMSC-derived apoptotic bodies accumulate through electrostatic interaction with positively charged neutrophil peripheral traps (NETs). Apoptotic vesicles convert NETosis to apoptosis through the Fas ligand-activated Fas pathway (Ou et al., 2022). Through the expression of associated speck-like protein containing caspase recruitment domain, pyroptotic EVs can bind to B cells in the spleen and bone marrow and inhibit cell death, thereby reducing the inflammatory response. Pyroptotic EVs also modulate the immune response by inhibiting toll-like receptor 4 (TLR4) and improve survival in mice with sepsis (Huang et al., 2024).
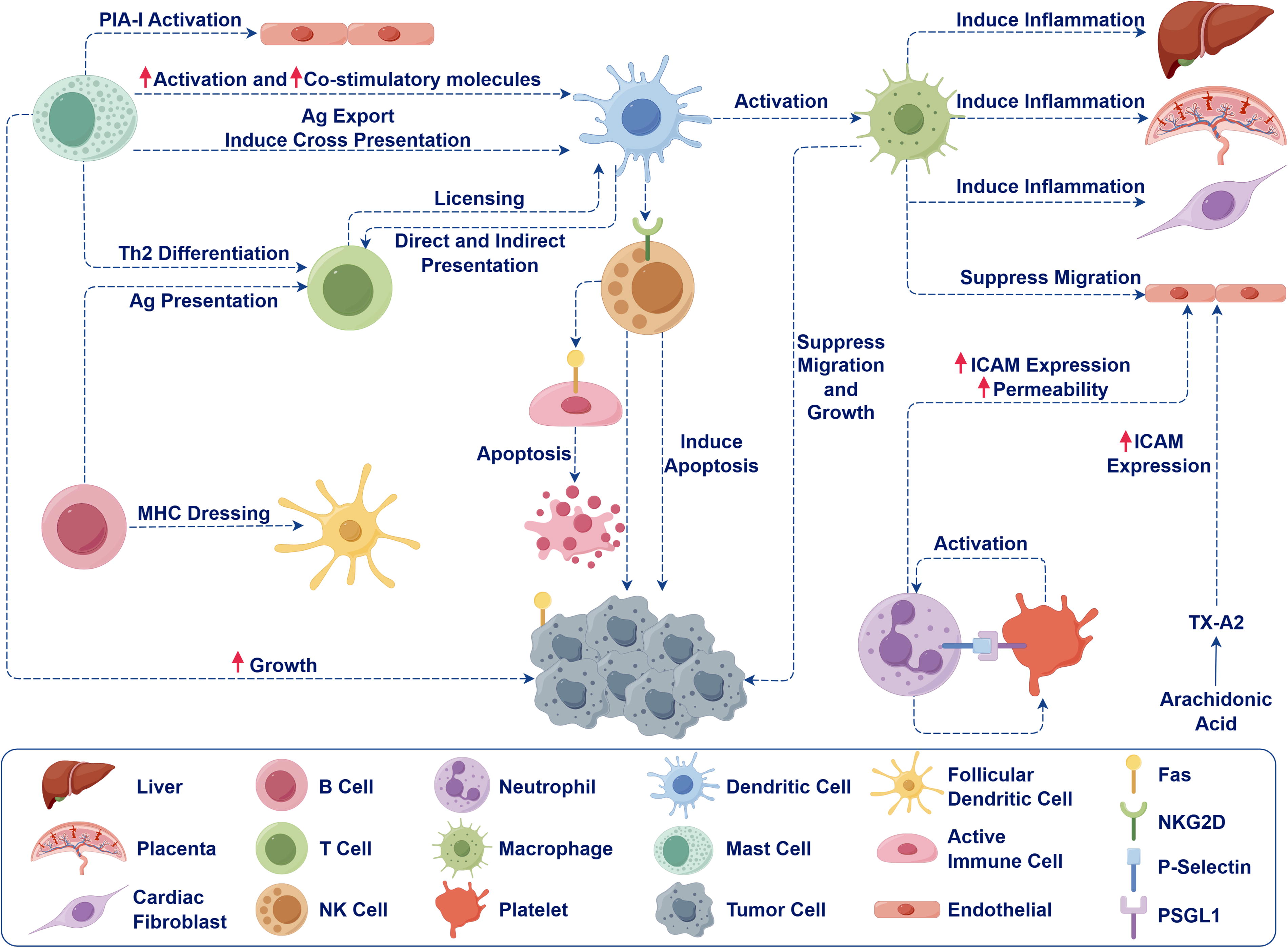
Effects of exosomes on immune cells (By Figdraw. ID: OSRSRa1356).
In addition to their immunomodulatory effects, MSC-EVs have been shown to exert organ-protective effects by modulating cellular oxidative stress and various cell death processes in sepsis (Xu et al., 2021). Specifically, MSC-EVs enhance pulmonary microvascular permeability, mitigate histopathological changes, and suppress neutrophil infiltration in lung tissue. Mechanistic investigations suggest that MSC-EVs may mediate these effects by inhibiting the phosphorylation of the MAPK/NF-κB pathway, as well as by enhancing the activity of Nrf2 and HO-1 (Chen et al., 2021, 2023; Su et al., 2019). ADSC-EVs can deliver lncRNA DLEU2 to lung epithelial cells, promote their viability and proliferation, and inhibit the inflammatory response and apoptosis, thereby improving lung injury induced by sepsis (He et al., 2024). In the context of sepsis-induced lung damage, BMSC-EVs that encapsulate serum amyloid A1 (SAA1) have been shown to mitigate the effects of this condition. Specifically, in murine models, upregulation of SAA1 resulted in a decrease in LPS-induced lung injury, as well as a reduction in levels of endotoxin, TNF-α, and IL-6 (Lv et al., 2022). MSC-EVs regulate the phosphatase and tensin homolog/β-catenin axis by carrying microRNA (miRNA)-141 and reduce myocardial damage induced by abdominal infection (Pei et al., 2021). Additionally, lncRNA-p21 in MSC-EVs inhibited miR-181 levels and reduced epithelial cell apoptosis by upregulating silent information regulator 1 expression, thereby alleviating lung tissue damage (Sui et al., 2021). miR-223 in MSC-EVs play a cardioprotective role in sepsis by regulating Semaphorin 3A and signal transducer and activator of transcription 3, reducing inflammation and death of cardiomyocytes (Wang et al., 2015). MSC-EVs improve the myocardial mitochondrial calcium ion disturbance and cell damage caused by severe infection (Zhou et al., 2021). ADSC-EVs specific delivery of miR-125b-5p increased Nrf2 expression and nuclear translocation and alleviated ferropotosi in pulmonary microvascular endothelial cells (Shen et al., 2023).
In conclusion, MSC-EVs play a significant role in the prevention and treatment of sepsis through diverse mechanisms including immune modulation, antioxidative stress, and inhibition of cellular apoptosis. Through the delivery of specific miRNAs, lncRNAs, noncoding RNAs, and proteins, MSC-EVs modulate crucial signaling pathways in target cells, leading to therapeutic outcomes such as anti-inflammatory responses. Further investigation into the underlying mechanisms and essential components of MSC-EVs is essential for advancing their translational application in the management of sepsis.
Immune cell-derived EVs
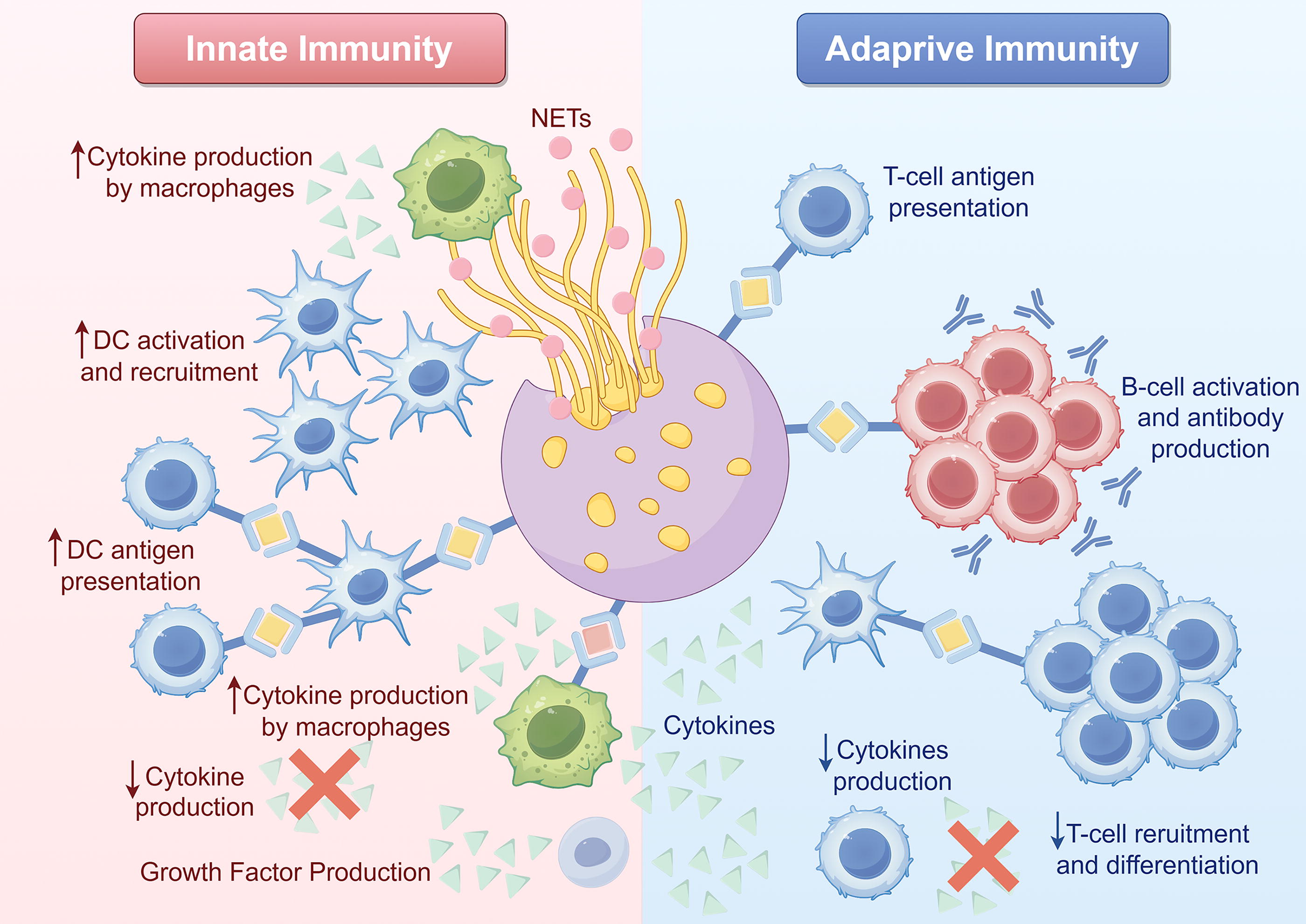
The dysregulation of immune cells is crucial in the pathogenesis of various diseases, with overactivation of neutrophils and mononuclear/macrophages leading to cytokine storms and subsequent tissue and organ damage (Gotts and Matthay, 2016). EVs transport bioactive molecules and facilitate intercellular communication, thereby playing a significant role in the pathophysiology of sepsis and other pathological conditions (Paz-Ares et al., 2019).
Neutrophils, as important effector cells of innate immunity, produce different subtypes of EVs during inflammatory responses (Fig. 3). Neutrophils release neutrophil-derived trails (NDTRs) during migration to the inflammatory site, and release neutrophil-derived EVs (NDEVs) after reaching the inflammatory site (Youn et al., 2021). NDTRs can activate phagocytes, while NDEVs inactivate bystander cells, which have different effects on immune responses (Prakash et al., 2012). Studies have found that the expression of NDEVs’ miR-150-5p in patients with sepsis is significantly increased, and it is involved in pathological processes such as inflammatory responses, myocardial apoptosis, and mitochondrial dysfunction caused by sepsis by targeting a variety of genes. The combination of miR-150-5p with NT-pro BNP, left ventricular ejection fractions, sequential organ failure assessment score (SOFA), and other clinical indicators can independently predict the occurrence and severity of sepsis-induced myocardial lesions (Ye et al., 2023). Additionally, TNF-α stimulates the release of NDEVs, which affect alveolar M1 macrophage activation and death by delivering lncRNA HCG18 (Zhu et al., 2023). NDEVs also have antibacterial and antioxidant functions. In the sepsis mouse model, the exosomes contain lactoferrin and other antibacterial molecules (Kumagai et al., 2020). Circulating NDEVs containing mitochondria, which express superoxide dismutase 2 after exposure to LPS, play an antithrombotic role by clearing endothelial reactive oxygen species (Bao et al., 2022).
Neutrophils are important for maintaining immune homeostasis (By Figdraw. ID: STSRW42456).
Monocytes and macrophages are key innate immune cells involved in both infection and anti-infection responses. Targeting proinflammatory macrophage exosome secretion is an effective treatment strategy. For example, lung epithelial cells can secrete IL-25 to inhibit macrophages from secreting TNF-α-rich exosomes, thereby inhibiting LPS-induced damage (Li et al., 2018). Macrophages modulate the protein secretion of EVs via caspase-4 during LPS-induced nonclassical caspase 4/5 inflammasome activation (Lorey et al., 2017). This process is partially reliant on Nucleotide-binding domain and leucine-rich repeat (NLR) family pyrin domain containing 3 (NLRP3) and caspase-5, with LPS preconditioning amplifying its impact. The regulation of EV-mediated protein secretion is influenced by autocrine interferon signaling triggered by TLR4, potentially leading to paracrine effects (Cypryk et al., 2023). Macrophage-derived EVs also inhibit high mobility group box 1 (HMGB1) release (Liang et al., 2024). High levels of miR-6238 were detected in macrophage-derived EVs, which modulate neutrophil infiltration into the lung alveoli by downregulating C-X-C motif chemokine ligand 3 (Park et al., 2023a). The EVs secreted by M2 macrophages contain a high concentration of miR-24-3p, which mitigates myocardial apoptosis and systemic inflammation following sepsis through the suppression of tumor-necrosis-factor-related apoptosis-inducing ligand (Tnfsf10) expression (Sun et al., 2022). In addition, macrophage-derived EVs inhibited the release of TNF-α and IL-6, enhanced reactive oxygen species (ROS) production in recipient cells, and mediates dysregulation of oxygen homeostasis through cytochrome P450 (Zakeri et al., 2021). natural killer(NK) cells are also involved in the immune response to sepsis. EVs derived from NK cells stimulated by granulocytecolony-stimulating factor (G-CSF) and interferon α (IFN-α) can carry perforin and granzyme B to exert cytotoxic effects (Federici et al., 2020).
Existing studies have shown that EVs derived from immune cells play an important role in the pathogenesis of sepsis, and exert pathogenic or protective roles through the regulation of immune response, inflammatory factors, apoptosis, and other pathological links. Interventions targeting specific components and functions of EVs may provide new insights for the immunotherapy of sepsis.
Endothelial cell-derived EVs
EVs secreted by endothelial cells contains a variety of active substances with anti-inflammatory properties and endothelial protection, showing promising applications in the treatment of sepsis (Lehmann et al., 2018) (Fig. 4). Endothelial cell-derived EVs with heat shock protein A12B downregulate NF-κB activation and nuclear translocation (Tu et al., 2020). MiR-125b-5p in endothelial cell-derived EVs prevents sepsis-induced acute lung injury by inhibiting topoisomerase II α (Jiang et al., 2021). In small airway epithelial cells, miR-126-3p targets phosphoinositide-3-kinase regulatory subunit 2, and miR-126-5p inhibits inflammatory alarms of high mobility group box 1 protein and vascular permeability factor-α, both of which increase tight junction protein expression and reduce LPS-induced lung injury (Zhou et al., 2019). Exosomes containing miR-126-5p and 3p inhibited LPS-induced high mobility group box 1 protein and vascular cell adhesion molecule 1 levels in microvascular endothelial cells, respectively, and prevented microvascular dysfunction (Zhou et al., 2018). Endothelial cell-derived EVs also carry miR-93-5p and play a protective role by regulating the lysine (K)-specific demethylase 6B/histone H3 Lys27 trimethylation/TNF-α axis, which played a therapeutic role in the mouse CLP sepsis model (He et al., 2020). In sepsis, EVs released by intestinal epithelial cells contain miRNAs that suppress the expression of the pro-inflammatory cytokines TNF-α and IL-17A. Through the isolation of EVs from enteric lavages in septic mice, researchers observed the presence of typical exosomal markers (CD63 and CD9) and epithelial markers (EpCAM) on these EVs, which were found to be upregulated during sepsis (Appiah et al., 2020). Furthermore, administration of septic EVs into the inflamed gut led to a significant reduction in the expression of pro-inflammatory cytokines TNF-α and IL-17A. Further study of the biological characteristics and functional regulation mechanisms of endothelial EVs, and development of new diagnostic and therapeutic technologies related to endothelial EVs, are expected to provide new insights and strategies for the precision medicine of sepsis.

Formation of endothelial cell-derived exosomes (By Fig.draw. ID: IITATb0a10).
Tumor cell-derived EVs
Cytokine storm and immunosuppression are the main pathogenic factors (Padovani et al., 2024; Zhong et al., 2024). Tumor cells secrete a variety of immunosuppressive factors, and EVs play a key role in tumor immune escape (Graham et al., 2023; Lin et al., 2023). EVs secreted by tumor cells contain a variety of bioactive substances, such as microRNA, lncRNA, and proteins, which can transmit information between cells and regulate a variety of biological processes. Tumor EVs can induce T cell apoptosis, inhibit NK cell and dendrite cell function, promote the expansion of bone marrow-derived suppressor cells, and create an immunosuppressive microenvironment (Chak et al., 2024; Tankov et al., 2024). Considering the immunosuppressive characteristics of tumor EVs, there is potential for their utilization in managing tissue damage disorders resulting from immune hyperactivity. Teng et al. demonstrated that exosomes originating from H22 liver cancer cells have the ability to trigger lymphocyte apoptosis and mitigate the systemic organ injury induced by LPS in murine models, indicating the potential therapeutic application of tumor exosomes in inflammatory conditions like sepsis (Teng et al., 2012). In addition to direct application in sepsis treatment, EVs can also be used as a new vaccine adjuvant to enhance the immunogenicity and protective efficacy of sepsis vaccines (Li et al., 2022).
Flora-derived EVs
EVs secreted by bacteria are important pathogen-related molecular models, carrying virulence factors and effector molecules, mediating host-pathogen interactions and regulating immune responses (Ashbolt, 2023; Laakmann et al., 2023). Targeting EVs and their pathogenic mechanisms is expected to provide a new strategy for the treatment and prevention of sepsis. In vitro experiments have shown that inhibiting EV production or blocking EV interaction with host cells can reduce the LPS-induced inflammatory response (Gerritzen et al., 2019; Muller et al., 2021). EVs can also be used as a new drug screening system to identify more suitable drugs for the treatment of bacterial sepsis (Yuan et al., 2022). Pathogen EVs can also be used as a new drug delivery system to deliver antibacterial drugs and anti-inflammatory preparations to the infected foci, improve efficacy, and reduce toxic side effects. Because EVs are rich in conserved pathogene-associated molecular patterns, they are ideal vaccine candidate antigens (Gerritzen et al., 2017). The use of inactivated bacterial EV as a vaccine can induce a broad spectrum of neutralizing antibodies and T cell response, and has shown good preventive effect in a variety of animal infection models (Higham et al., 2023; Hou et al., 2005). The polyvalent vaccine prepared by mixing EVs from multiple Gram-negative bacteria, such as four species of Staphylococcus aureus, can produce a synergistic immune response and prevent multidrug-resistant bacterial infections (Konig et al., 2021). Modifying the composition and immunogenicity of EV using synthetic biology techniques, such as fusion expression of host immunomodulators, can further enhance the protective efficacy of the vaccine (Park et al., 2023b).
Other source-derived EVs
Recent research has demonstrated elevated levels of platelet-derived EVs, particularly those containing miR-223-3p, in the circulating blood of sepsis patients, with evidence suggesting a potential protective effect on renal tubule epithelial cells in sepsis-induced acute injury (Wan et al., 2024). Platelet-derived EVs containing miR-223-3p can reduce NLRP3-induced pyroptosis. In vitro experiments confirmed that fibroblast-derived EVs could enhance the phagocytic function of macrophages and promote the bacterial clearance at infection foci (Kunz et al., 2017). Panax ginseng root-derived EVs target the nicotinamide adenine dinucleotide phosphate oxidase 4 (NADPH oxidase 4)/dynamin-related protein 1/NLRP3 inflammasome signaling pathway, alleviating pyroptosis and the inflammatory response in alveolar epithelial cells (Ma et al., 2024).
Conclusions
EVs are small membranous structures released by cells at the nanoscale level, capable of transporting a diverse array of bioactive molecules to facilitate intercellular communication. These EVs exhibit significant promise in the therapeutic management of sepsis. They contain specific miRNAs, lncRNAs, and other molecules that effectively modulate crucial signaling pathways in recipient cells, thereby exerting anti-inflammatory and organ-protective effects through the regulation of immune cell function, suppression of oxidative stress, and inhibition of apoptosis. Further investigation into the mechanisms of action of EVs and the development of innovative engineering strategies hold potential for the advancement of novel and precise treatments for sepsis.
Footnotes
Authors’ Contributions
L.K.: Writing—original draft, and investigation. Q.H.: Investigation, and visualization. J.C.: Investigation, and visualization, and resources. Y.D.: Conceptualization and writing review & editing. B.W.: Conceptualization, supervision, and funding acquisition.
Author Disclosure Statement
The authors report no conflicts of interest in this work.
Funding Information
This study was supported by the Jinhua Science and Technology Plan Public Welfare Project (2022-4-10), and Provincial Medical and Health Science and Technology Plan Project Achievement Promotion Project (2022ZH063).