
Abstract
Cellular plasticity, which refers to the capacity of cells to alter their identity or potency in response to a variety of stimuli, is emerging as an essential component in tissue repair. Despite the fact that stem cells have historically been considered to be the major agents of plasticity, new research has demonstrated that even differentiated cells in organs including the stomach, pancreas, and lungs are capable of displaying plasticity under specific physiological conditions, such as during injury and repair. One element essential for many physiological processes is vitamin B12 (VB12). Beyond its well-known roles in red blood cell production and nervous system maintenance, VB12 is critical for one-carbon metabolism and DNA synthesis and repair, processes indispensable for cellular health and tissue integrity. With its wide spectrum of actions, VB12 may have the potential to significantly influence tissue plasticity and repair, paving the way for new therapeutic interventions. Investigating fundamental processes and considering consequences for illness and aging, this perspective contemplates the junction of VB12, cell plasticity, and tissue repair.
Cell Plasticity and Tissue Repair
One of the most extreme instances of cell plasticity, allowing fully differentiated cells to return to full embryonic pluripotency, is reprogramming by the so-called Yamanaka transcription factors Octamer-binding transcription factor 4 (Oct4), Sex-determining region Y (SRY) box 2 (Sox2), Krüppel-like factor 4 (Klf4), and cellular Myelocytomatosis oncogene (c-Myc), Oct4, Sox2, Klf4, and c-Myc (OSKM). Partial reprogramming in situ/in vivo by OSKM—whereby OSKM expression is terminated prior to acquisition of full embryonic pluripotency—can promote tissue repair. This has been demonstrated in various organs including the intestine (Kim et al., 2023), pancreas (Chondronasiou et al., 2022), skin (Ocampo et al., 2016), and heart (Chen et al., 2021), providing proof-of-concept evidence that promoting cell plasticity is an avenue for regenerative medicine/tissue repair.
Interestingly, physiological parallels of cell plasticity in the context of injury response have been observed across different organs, whereby novel cell states emerge, which are necessary for repair. The gastrointestinal (GI) tract provides compelling examples of cellular plasticity, in the form of reprogramming events that allow these tissues to regenerate and maintain their critical functions. In the stomach, following an injury such as ulceration, the cells in the gastric pit can undergo transdifferentiation into spasmolytic polypeptide-expressing metaplasia (SPEM) cells (Goldenring et al., 2010). SPEM cells can play a key in repairing damage, but they’re also linked to precancerous changes, showing how cellular plasticity can be both a healing mechanism and a potential risk. Further along the GI tract, the intestine also demonstrates plasticity, with many epithelial cell types dedifferentiating into fetal-like progenitor states to enable rapid repair after injury (Ayyaz et al., 2019; Yu et al., 2018). Plasticity is observed in other organs, including the liver, where hepatocytes can transdifferentiate to aid regeneration (He et al., 2014), and the skin, where specialized epidermal cells can dedifferentiate and acquire stem cell properties to contribute to tissue repair after wound injury (Donati et al., 2017). Similarly, pancreatic acinar cells can transdifferentiate into ductal cells following injury (Houbracken et al., 2011), and in the lungs, alveolar cells can change identity to replace damaged tissue (Liberti et al., 2021; Shiraishi et al., 2024).
By understanding the molecular mechanisms behind plasticity, researchers may develop new therapies that enhance tissue repair in a wide range of tissues and pathologies. Interestingly, vitamin B12 (VB12) supplementation has recently been shown to enhance reprogramming in mouse models with inducible OSKM expression, and in the context of physiological plasticity upon intestinal damage (Kovatcheva et al., 2023), leading us to review what is currently known about the role of VB12 in plasticity and repair.
VB12 in Metabolism and Tissue Plasticity
Many metabolic events depend on VB12. It acts as a cofactor for methionine synthase (MS) and methylmalonyl-CoA mutase (MMUT), enzymes crucial for one-carbon metabolism, nucleotide biosynthesis, and energy production (Shipton and Thachil, 2015). One-carbon metabolism involves the transfer of one-carbon units between molecules, a process that is crucial for DNA methylation and synthesis. MS facilitates the conversion of homocysteine (Hcy) to methionine, a process that concurrently regenerates tetrahydrofolate, a crucial precursor in nucleotide biosynthesis. This mechanism guarantees that cells possess an adequate supply of nucleotides for DNA replication, which is crucial for cell division and tissue repair (Halczuk et al., 2023). MMUT is involved in the conversion of methylmalonyl-CoA to succinyl-CoA, which is an important step in the degradation of certain amino acids and fatty acids. This process is part of the tricarboxylic acid cycle, a major metabolic route that provides energy for cells. These pathways are not only critical for DNA synthesis but also have important implications for cellular plasticity and repair.
Low VB12 levels can result in a deficit in nucleotide pools, causing growth arrest and mutations. This is because DNA replication is highly sensitive to the supply of nucleotides and any imbalance in these building blocks could induce mistakes during replication (Halczuk et al., 2023). Such errors can lead to genomic instability, which is a hallmark of cancer and other diseases. Moreover, it has been shown that VB12 deficiency in rat liver can also impair the function of MMUT, leading to the accumulation of methylmalonic acid, a metabolite that can be toxic to cells (Toyoshima et al., 1996). This is crucial to consider, as most adult epithelial tissues have low proliferation rates; however, injury and regeneration necessitate a significant increase in cell proliferation. Any disruption in nucleotide pools, such as that caused by VB12 deficiency, could severely impair the regenerative capacity of these tissues. VB12 also plays a crucial role in nucleotide biosynthesis by indirectly supporting the synthesis of deoxythymidine monophosphate (dTMP), essential for DNA replication. It does this through folate metabolism, as VB12 is required to convert methyltetrahydrofolate (5-methyl-THF) to THF, a precursor for dTMP synthesis (Palmer et al., 2017). S-adenosylmethionine sensor upstream of mTORC1 (SAMTOR), a sensor of SAM levels, has been shown to integrate with mTOR signaling (Gu et al., 2017). As VB12 is involved in metabolism and especially SAM synthesis, it could influence SAMTOR activity and, by extension, mTOR signaling. Moreover, SAM levels are known to play a crucial role in epigenetic modifications (Liu et al., 2020; Mentch and Locasale, 2016), including histone and DNA methylation, essential for the maintenance of cellular identity. Intriguingly, a study demonstrated that VB12 supplementation resulted in alterations in DNA methylation of specific genes (Kok et al., 2015), including DIRAS family GTPase 3 (DIRAS3), Armadillo repeat-containing protein 8 (ARMC8), and Nodal growth differentiation factor (NODAL). These genes are implicated in carcinogenesis and early embryonic development, processes closely linked to cellular plasticity. VB12 supplementation also enhances Trimethylation of lysine 36 on histone H3 (H3K36me3) levels in a SAM-dependent manner (Kovatcheva et al., 2023), which has been shown to correlate with increased reprogramming efficiency in mouse embryonic fibroblasts and adult mammalian stem cell function (McCauley et al., 2021). This finding suggests a potential epigenetic mechanism through which VB12 may influence cellular plasticity and repair.
As we have shown, VB12 plays a critical role in several metabolic processes that are essential for cellular plasticity and repair. Nevertheless, how VB12 influences these mechanisms in these various settings remains largely unexplored, and warrants future investigation.
VB12 in Inflammation and Anti-Inflammatory Responses
Apart from its function in metabolism, studies indicate an inverse correlation between circulating VB12 levels and inflammatory markers such as interleukin 6 and C-reactive protein in both high-risk cardiovascular patients and aging models (Domínguez‐López et al., 2024; Mahalle et al., 2013), consistent with a broad anti-inflammatory role for VB12. One of the primary pathways in which VB12 modulates inflammation is through its participation in Hcy metabolism. VB12 is an essential cofactor for the conversion of Hcy to methionine, reducing toxic protein modifications and restoring cellular homeostasis. But, in the context of VB12 insufficiency, Hcy levels can increase, resulting in hyperhomocysteinemia (Hhcy) (Moretti and Caruso, 2019). Hhcy is a marker of aging, which is linked to increased inflammation and oxidative stress, both of which can impair tissue repair. High levels of Hhcy have been associated with a variety of inflammatory disorders (Li et al., 2015), including cardiovascular (Mahalle et al., 2013) and chronic renal diseases (Dierkes et al., 1999). In the context of acute pancreatitis, it has been shown that VB12 supplementation helps reduce pancreatic tissue necrosis by increasing ATP levels and decreasing T cell infiltration. This mechanism helps prevent tissue damage and promotes recovery (Chen et al., 2024). Furthermore, in non-alcoholic steatohepatitis, VB12 has been effective in reducing inflammation and fibrosis by preventing the homocysteinylation of critical proteins (Tripathi et al., 2022).
The anti-inflammatory effects of VB12 could be mediated by epigenetic mechanisms. VB12 enhances the sulfur amino acid pathway and increases the production of SAM, which can potentiate DNA methylation and silence gene expression. This mechanism has been shown to occur during COVID-19 infection, whereby patients receiving VB12 supplementation had strong downregulation of chemokine (C-C motif) ligand 3 (CCL3) and interleukin 1 beta (ILB1), associated with hypermethylation in regulatory regions of these genes (Cassiano et al., 2023). However, while inflammation is necessary to initiate tissue plasticity and repair, it can act as a “double-edged sword.” Inflammation, even if it plays a protective role in the short term, can hinder recovery and lead to more severe conditions, such as cancerous lesions. This highlights the importance of managing the timing and dosage of VB12, which could be crucial for maintaining a balanced inflammatory response. These considerations justify further research to better understand the potential applications of VB12 in regenerative medicine.
It is known that vitamins have antioxidant effects, and in certain organisms such as C. elegans, a deficiency in VB12 leads to a high increase in oxidative stress (Bito et al., 2017). However, there is not much evidence regarding the antioxidant role of VB12 in cellular plasticity and repair. Nevertheless, VB12 acts as a key antioxidant, protecting cells from oxidative stress by maintaining glutathione levels (Girish et al., 2022), reducing reactive oxygen species (Yuan et al., 2021), and improving mitochondrial and protein function in colon cancer tissue (Taranikanti et al., 2020). In an in vivo experiment, VB12 demonstrated radioprotective effects, notably by reducing oxidative stress and DNA damage caused by a UV source (Attia et al., 2023). Nevertheless, future investigations will be necessary to interrogate how these functions of VB12 might directly influence cellular plasticity and repair.
VB12 and Aging
Aging correlates with a reduction in tissue repair capabilities, and VB12 deficiency is common in the older population (Stover, 2010). This deficit often results from malabsorption in the stomach and intestine, where alterations in the mucosa and reduced intrinsic factor synthesis hinder VB12 absorption (Guéant et al., 2022). Considering VB12’s functions in DNA repair, inflammatory modulation, and metabolic processes, it is tempting to speculate that its systemic deficiency may contribute to impaired tissue repair observed in aging. Low VB12 levels have been linked to increased inflammation, oxidative stress, and genomic instability, all of which are hallmarks of aging (López-Otín et al., 2023). VB12 supplementation could be one potential approach to addressing these issues. Several studies suggest that supplementing with VB12 may improve cognitive function, reduce inflammation, and enhance overall health in elderly individuals (Dangour et al., 2015; Oliai Araghi et al., 2021). Nevertheless, the overall results of such studies are currently inconclusive. This might be explained by the fact that supplementing only VB12 may not be enough. A study demonstrated that SAM depletion in aged muscle stem cells leads to a loss of heterochromatin, resulting in genomic instability, DNA damage accumulation, and impaired regenerative capacity (Kang et al., 2024). Interestingly, restoring SAM levels in these cells rejuvenates heterochromatin formation and improves cell function. Since both folate and VB12 are necessary for SAM synthesis, VB12 supplementation alone may yield limited benefits. A combination of several vitamins supporting one-carbon metabolism might be required to achieve significant improvement. Further research is needed to fully understand the complex interplay of factors in aging.
Currently, the effects of VB12 supplementation on tissue repair—alone or in combination with other metabolic interventions—in aging populations have not been extensively studied, and this remains an important area for future research. Since aging is multifactorial, it would be worthwhile to examine and integrate the consequences on both cell-autonomous (i.e., parenchymal cell) and systemic (eg., immune) functions.
Conclusion
VB12 is a complex nutrient that is essential for metabolism, DNA synthesis, cellular health, and inflammation control (Fig. 1). These processes are necessary to maintain tissue resilience and promote tissue repair. The prevalence of VB12 deficiency increases with age, which may compromise tissue repair and exacerbate age-related degeneration. While much is known about VB12’s role in these processes, many questions remain unanswered. Mechanistically, how does VB12 deficiency impact tissue plasticity in different organs? Can VB12 supplementation improve tissue repair outcomes in aging populations? These are pertinent questions to be addressed in future studies.
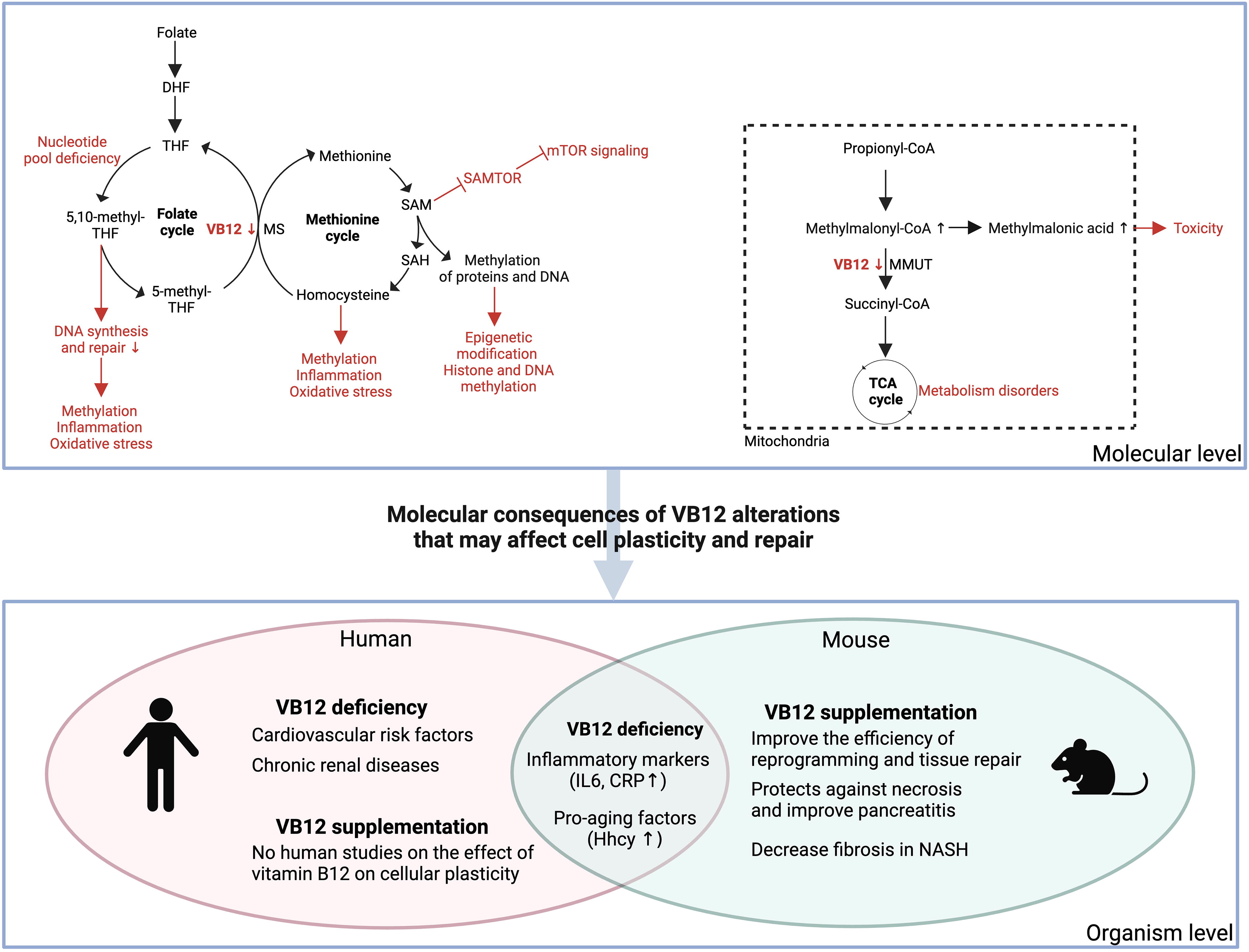
Vitamin B12 deficiency at the molecular level and its organismal consequences that may affect cell plasticity and repair in humans and mice. CRP, C-reactive protein; DHF, dihydrofolate; Hhcy, hyperhomocysteinemia; IL6, interleukin 6; MMUT, methylmalonyl-CoA mutase; MS, methionine synthase; NASH, Nonalcoholic steatohepatitis; SAH, S-adenosylhomocysteine; SAM, S-adenosyl methionine; SAMTOR, S-adenosylmethionine sensor for the mTORC1; TCA, tricarboxylic acid; THF, tetrahydrofolate; VB12, vitamin B12.
Finally, it is interesting to consider that VB12 supplementation can mimic some of the beneficial effects of reprogramming OSKM (Kovatcheva et al., 2023). Transient OSKM expression have shown promise in promoting tissue repair, but may not be feasible for general clinical use. If VB12 or other metabolic interventions could reproduce some of these effects by enhancing or fine-tuning the natural cell plasticity that occurs during tissue repair, it could offer a more accessible approach to enhancing tissue plasticity and repair.
Footnotes
Authors’ Contributions
C.F. and M.K.: Both conceptualized, co-wrote, and revised the article.
Author Disclosure Statement
All authors have no conflicts of interest to disclose.
Funding Information
Authors of this work were supported by Impetus Longevity Grants, The European Association for Cancer Research (EACR), The Mark Foundation, and The Pezcoller Foundation.