
Abstract
Purpose:
Because of a dismal prognosis for advanced renal-cell carcinoma (RCC), an alternative therapeutic approach, using vitamin K3 (VK3) and D-fraction (DF) was investigated. VK3 is a synthetic VK derivative and DF is a bioactive mushroom extract, and they have been shown to have antitumor activity. We examined if the combination of VK3 and DF would exhibit the improved anticancer effect on RCC in vitro.
Materials and Methods:
Human RCC, ACHN cell line, were treated with varying concentrations of VK3, DF, or a combination of the two. Cell viability was assessed at 72 hours by MTT assay. To explore the possible anticancer mechanism, studies on cell cycle, chromatin modifications, and apoptosis were conducted.
Results:
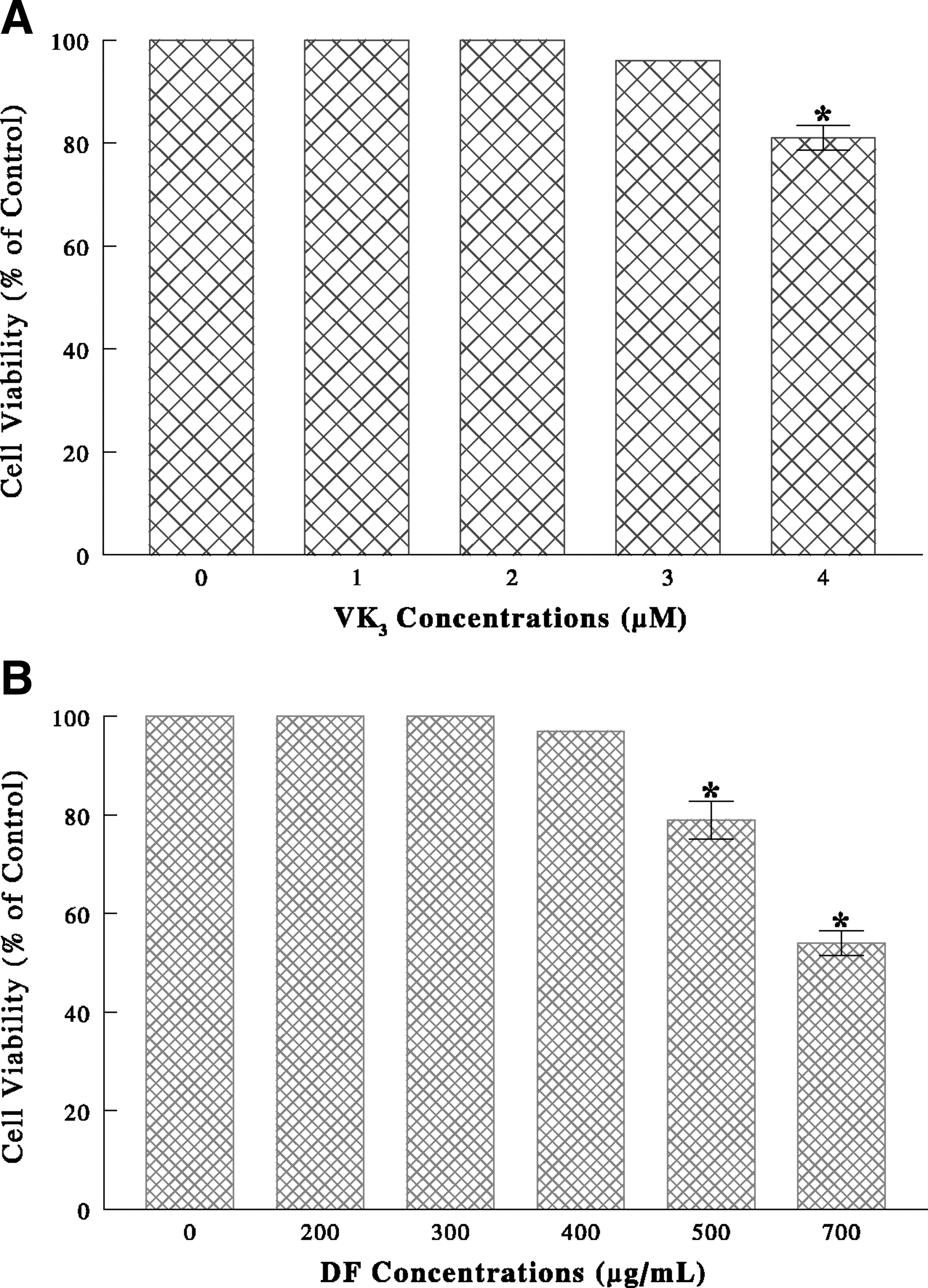
VK3 alone led to a ∼20% reduction in cell viability at 4 μM, while DF alone induced a 20% to 45% viability reduction at ≥500 μg/mL. A combination of VK3 (4 μM) and DF (300 μg/mL) led to a drastic >90% viability reduction, however. Cell cycle analysis indicated that VK3/DF treatment induced a G1 cell cycle arrest, accompanied by the up-regulation of p21WAF1 and p27Kip1. Histone deacetylase (HDAC) was also significantly (∼60%) inactivated, indicating chromatin modifications. In addition, Western blot analysis revealed that the up-regulation of Bax and activation of poly-(ADP-ribose)-polymerase (PARP) were seen in VK3/DF-treated cells, indicating induction of apoptosis.
Conclusions:
The combination of VK3 and DF can lead to a profound reduction in ACHN cell viability, through a p21WAF1-mediated G1 cell cycle arrest, and ultimately induces apoptosis. Therefore, the combination of VK3/DF may have clinical implications as an alternative, improved therapeutic modality for advanced RCC.
Introduction
R
Unfortunately, conventional therapeutic modalities for metastatic RCC, including cytoreductive surgery, radiotherapy, chemotherapy, or immunotherapy, were not very effective. 3,8 Newer, specifically targeted treatment options for distant metastasis, including tyrosine kinase inhibitors (sunitinib, etc.), mammalian target of rapamycin inhibitors (temsirolimus, etc.), and monoclonal antibodies (G250, etc.), are promising, but the survival results are still poor. 9 These reasons thus prompted us to explore a novel treatment for patients with metastatic RCC.
We have been working with vitamins and natural agents/substances as an alternative approach to such metastatic RCC cases. Particularly, we were interested in vitamin K3 (menadione; VK3) as a potential therapeutic agent. VK3 is a synthetic derivative of naturally occurring fat-soluble vitamin K (VK) 10 and has been shown to have an antitumor effect on several cancer cells. 11 –13 Because VK3 requires a relatively high dose to be effective, 11 combinations of VK3 and other agents have been postulated to improve its efficacy at a lower dose.
At the same time, we have also been studying the bioactive extract from maitake (Grifola frondosa), known as D-fraction (DF). DF has been extensively studied for the past 30 years and has been shown to have immunomodulatory and antitumor activities. 14 –16 For instance, antitumor activity of DF has been demonstrated in tumor-bearing mice through activation of various immune effectors. 14,15 In addition, induction of apoptosis by DF was also reported in breast cancer cells. 16 DF thus appears to be an interesting and promising natural agent that could be used in cancer treatment.
The purpose of this study was to investigate whether a combination of VK3 and DF would exhibit the improved and enhanced anticancer effect on RCC in vitro, as an alternative therapeutic approach. These studies were performed focusing mainly on cell viability, cell cycle, chromatin modifications, and apoptosis.
Materials and Methods
Cell culture
The human renal carcinoma ACHN cells (with aggressive grade IV property) were maintained in RPMI 1640 medium containing 10% fetal bovine serum, penicillin (100 units/mL), and streptomycin (100 μg/mL). For experiments, ACHN cells were seeded at the initial cell density of 2×105 cells/mL in six-well plates or T-75 flasks and cultured with varying concentrations of VK3, DF, or their combinations. Cell viability was then assessed at 72 hours by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) assay.
Cell viability assay (MTT assay)
Cell viability was determined by MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl-tetrazolium bromide) assay following the protocol of the vendor (Sigma-Aldrich, St. Louis, MO). Briefly, at the harvest time, MTT reagent (0.5 mg/mL) was added to each well in the six-well plate, followed by 3-hour incubation at 37°C. After removing MTT reagent, dimethyl sulfoxide was added to each well and absorbance of formazan solution was read on a microplate reader. Cell viability was then expressed as percentage (%) of viable cells relative to the control reading (100%).
Cell cycle analysis
A BD FACscan flow cytometer (Franklin Lakes, NJ), equipped with a double discrimination module, was used for cell cycle analysis. Approximately 1×106 cells were resuspended in 500 μL of propidium iodide solution and incubated at room temperature for 1 hour. Ten thousand nuclei were analyzed for each sample, quantified in cell cycle compartments, and estimated as cell cycle phase fractions.
Histone deacetylase (HDAC) assay
HDAC activity was measured using the Epigenase HDAC Assay Kit following the protocol of the manufacturer (Epigentek, Farmingdale, NY). Briefly, 10 μg of nuclear extracts from each sample were added to the coated microplate wells. After 90-minute incubation at 37°C, all wells in the plate were treated with 1st antibody for 60 minutes, followed by 30-minute incubation with 2nd antibody. The plate was then treated with reaction solution for 10 minutes, and the reaction was terminated with stop solution. Absorbance of samples was read on a microplate reader, and HDAC activity was calculated and expressed by the % relative to controls (100%).
Western blot analysis
The detailed procedures are described elsewhere. 17 An equal amount of cell lysates (7 μg), obtained from control and agent-treated cells, was first subjected to 10% SDS gel electrophoresis and transferred to a nitrocellulose membrane (blot). The blot was incubated for 90 minutes with the primary antibodies against p21WAF1, p27Kip1, Bax, or poly-(ADP-ribose)-polymerase (PARP) (Santa Cruz Biotechnology, Santa Cruz, CA), followed by 30-minute incubation with the secondary antibody conjugates. The specific immunoreactive protein bands were then detected by chemiluminescence following the protocol of the manufacturer (KPL, Gaithersburg, MD).
Statistical analysis
All data were presented as mean±standard deviation, and statistical differences between groups were assessed with either one-way analysis of variance or the unpaired Student t test. Values of P<0.05 were considered to indicate statistical significance.
Results
Effects of VK3, DF, or their combinations on ACHN cell growth
ACHN cells were cultured separately with varying concentrations of VK3 (0–4 μM) or DF (0–700 μg/mL). After 72 hours, MTT assay showed that VK3 alone had little effects but led to a ∼20% reduction in cell viability at 4 μM (Fig. 1A), while DF induced a 20% to 45% viability reduction at ≥500 μg/mL (Fig. 1B). Whether the combination of VK3 and DF may potentiate the anticancer effect was also examined. Such study showed that a profound cell viability reduction, from severe (∼90%) cell death, was attained with the specific combination of 4 μM VK3 and 300 μg/mL DF (Fig. 2).

Effects of combination of vitamin K3 (VK3) and D-fraction (DF) on cell viability. After cells were treated with VK3 (4 μM), DF (300 μg/mL), or their combination for 72 hours, cell viability was assessed by MTT assay and expressed by the % relative to controls. The data are mean±standard deviation from three separate experiments (*P<0.05 compared with controls).
Effects of VK3/DF combination on cell cycle
To explore how such VK3/DF combination would induce a viability reduction or cell death, cell cycle analysis was performed. Cells exposed to VK3/DF combination for 6 hours showed a 42% increase in the G1-phase cell number with a 60% decrease in the S-phase cells (Fig. 3). This accumulation of cells in the G1 phase, from a blockade of the cell cycle transition from the G1 to the S phase, is known as a G1 cell cycle arrest. 18 Thus, VK3/DF combination appears to disrupt the cell cycle at a G1 checkpoint.

Cell cycle analysis. Cells treated with or without vitamin K3 (VK3) (4 μM)/ D-fraction (DF) (300 μg/mL) combination for 6 hours were subjected to cell cycle analysis, and the % of cell distribution was plotted against the respective cell cycle phase. The data are mean±standard deviation from three separate experiments (*P<0.01 compared with controls).
Effects of VK3/DF combination on G1 cell cycle regulators
VK3/DF-induced G1 cell cycle arrest was further confirmed by analyzing the specific cell cycle regulators for the G1-S phase transition. Western blots revealed that compared with controls, the expressions of two G1 cyclin-dependent kinase inhibitors, p21WAF1 and p27Kip1, were significantly enhanced or up-regulated with VK3/DF treatment for 72 hours (Fig. 4). Because this up-regulation of p21WAF1 and p27Kip1 is indicative of a G1 cell cycle arrest, 18 the VK3/DF combination appears to induce a G1 arrest, leading to a growth cessation and consequently resulting in the cell viability reduction.
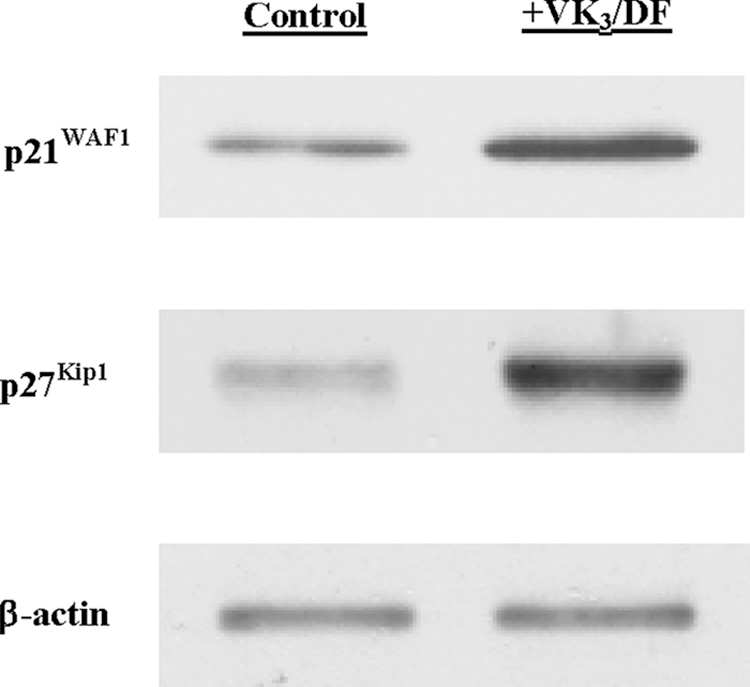
Analysis of G1 cell cycle regulators. Expressions of p21WAF1 and p27Kip1 in cells treated with vitamin K3 (VK3) (4 μM)/D-fraction (DF) (300 μg/mL) combination for 72 hours were analyzed on Western blots. Autoradiographs of p21WAF1 and p27Kip1 in control and VK3/DF-treated cells are shown for comparison.
Effects of VK3/DF combination on HDAC activity
Because p21WAF1 is known to be regulated by chromatin modifications from alterations in the acetylation state of histones, 19,20 such a possibility was tested on HDAC, one of the primary regulators of histone acetylation. 21 Cells were treated with VK3 (4 μM), DF (300 μg/mL), or their combination for 24 hours. Assays showed that VK3 or DF alone had little effects whereas the VK3/DF combination led to a ∼60% reduction in HDAC activity (Table 1). Thus, these results suggest that the VK3/DF combination may significantly inactivate HDAC, inducing hyperacetylation of specific histones 19 that could then up-regulate p21WAF1 as shown in Fig. 4.
Mean±standard deviation of three separate experiments (control 100%=0.91 OD/min/mg).
Values in parentheses are the percent (%) of reduction in HDAC activity relative to controls.
HDAC=histone deacetylase; VK3=vitamin K3; DF=D-fraction; OD=absorbance reading at 450 nm.
Effects of VK3/DF combination on apoptosis regulators
Because it was of interest to also explore how the VK3/DF combination would induce cell death, the possible role of apoptosis was then investigated. Cells treated with the VK3/DF combination for 12 hours were analyzed for expressions of two key apoptosis regulators on Western blots. VK3/DF treatment led to the up-regulation of pro-apoptotic Bax and activation of pro-apoptotic PARP (Fig. 5). Because the up-regulation/activation of Bax and PARP indicates induction of apoptosis, 22,23 it is plausible that VK3/DF-induced cell death may follow apoptosis.

Effects of vitamin K3/D-fraction (VK3/DF) on apoptosis regulators. Expressions of Bax and poly-(ADP-ribose)-polymerase (PARP) in cells exposed to VK3 (4 μM)/DF (300 μg/mL) combination for 12 hours were analyzed on Western blots. Autoradiographs of Bax and PARP in control and VK3/DF-treated cells are shown.
Discussion
To explore an alternative, minimally invasive, and more effective treatment for advanced RCC, we investigated the potential anticancer effect of VK3 and DF on human renal carcinoma ACHN cells. VK3 by itself had some effect with a ∼20% cell viability reduction, while DF needed relatively high concentrations (≥700 μg/mL) to be effective. When two agents with varying concentrations were combined, however, a specific combination of 4 μM VK3 and 300 μg/mL DF led to a drastic ∼90% reduction in cell viability, due to severe cell death (Fig. 2). This finding was somewhat extraordinary and prompted us to explore the cytotoxic mechanism of the VK3/DF combination.
To obtain a better understanding of the VK3/DF cytotoxic mechanism, cell cycle analysis was performed. The results indicated that a G1 cell cycle arrest could be the primary cause of a growth cessation (Fig. 3), resulting in a cell viability reduction. A G1 cell cycle arrest was also confirmed by the up-regulation of both p21WAF1 and p27Kip1 (Fig. 4), which were the inhibitors of the G1-specific cell cycle regulators. 18 These findings suggest that a VK3/DF combination could directly interfere with the cell cycle, particularly at the G1-S phase transition.
It is also plausible that a G1 cell cycle arrest could be a prerequisite for cell death induced by the VK3/DF combination. In particular, p21WAF1 has been shown to be associated with a G1 cell cycle arrest31 and its up-regulation (inducing a G1 arrest) is also known to result from histone modifications in the chromatin. 19 Our study revealed that HDAC was significantly (∼60%) inhibited by the VK3/DF combination (Table 1), presumably inducing hyperacetylation of histones that would then up-regulate p21WAF1. This finding thus suggests that the VK3/DF combination may induce chromatin modifications. The actual acetylation state of specific histones H3 and H4 with other related factors, however, needs to be further examined for confirming such key histone modifications. This study is under way in our laboratory.
Moreover, it was our rational attempt to examine whether the VK3/DF-induced cell death might be associated with apoptosis because it could provide us with valuable information. We then found that two pro-apoptotic regulators, Bax and PARP, were clearly up-regulated with the VK3/DF combination (Fig. 5), indicating induction of apoptosis. 22,23 Thus, the VK3/DF combination appears to ultimately induce apoptotic cell death, accounting for the profound cell viability reduction.
Finally, it is worth discussing the safety profile of DF because of its possible clinical implications. The United States Food and Drug Administration has exempted DF from a phase I study of toxicology and approved it for the Investigational New Drug application for a phase II pilot study on advanced cancer patients. 24 Therefore, the safety of DF has been granted, and it is safe to be used in both normal subjects and a variety of patients. Regarding VK3, it has been studied in several cancer cells, 11 –13 and we believe that the effective concentrations of VK3 (when combined with DF) would be more likely safe and nontoxic. We are going to perform a toxicology test on VK3 for confirmation using animals in our next study, however.
The findings in this study are interesting and encouraging. It is possible that the combination of VK3 and DF could be used as an adjuvant therapy to help reduce the incidence of potential recurrence after surgical treatment of RCC. However, preclinical studies (using animals) are required, however, for assessing the safety and true efficacy of the VK3/DF combination before clinical trials. Accordingly, the next phase of our study will primarily focus on the actual effect of the VK3/DF combination on RCC-bearing animals (athymic nude mice).
Conclusions
The present study demonstrates that the combination of VK3 and DF is highly cytotoxic, inducing a ∼90% cell death in ACHN cells. Such cell death is preceded with a p21-dependent G1 cell cycle arrest presumably through histone modifications. This cell cycle arrest will ultimately lead to apoptosis. Therefore, the VK3/DF combination appears to be promising and may have clinical implications as an alternative and effective therapeutic modality for advanced RCC.
Footnotes
Disclosure Statement
No competing financial interests exist.