
Abstract
Campylobacters are susceptible to environmental conditions such as starvation, temperature, and oxidative stress. Species such as Campylobacter jejuni have developed a number of mechanisms for responding to these conditions. We conducted a study to investigate whether survival of C. jejuni and pathogen–host cell interactions such as adherence, invasiveness, and intraepithelial survival in pig small-intestinal (PSI) epithelial cells are altered in response to starvation, changes in temperature, and atmospheric oxygen concentration. We assessed the ability of C. jejuni to translocate across polarized intestinal epithelial cell monolayers by measuring transepithelial electrical resistance (TER). Following heat stress, we observed loss of C. jejuni culturability but not viability. Heat-stressed C. jejuni adhered efficiently to pig intestinal epithelial cells, but their invasiveness was significantly impaired when compared with unstressed C. jejuni. Prolonged exposure to atmospheric oxygen reduced the ability of C. jejuni to adhere to intestinal epithelial cells, whereas brief exposure increased invasiveness and intraepithelial survival. By comparison, nutrient limitation reduced adherence, invasiveness, and intracellular survival of C. jejuni. Adherence of C. jejuni strongly affected the pig intestinal epithelium, as reflected by a significant decrease in TER of polarized intestinal epithelial cells. No correlation between TER and the translocation capacity of C. jejuni was observed. Additionally, campylobacters were detected in the basal chamber of a functional small-intestinal epithelial cell model at 3 hours post infection, without a significant reduction in the TER value, suggesting transcellular transport of C. jejuni into the body.
Introduction
T
We conducted a study to analyze whether exposure to environmental stress modulates pathogen–host cell interactions involving C. jejuni, such as bacterial adherence, invasion, and intraepithelial survival, and gives these organisms a survival advantage in an in vitro cell culture model of PSI epithelial cells. We studied changes in the adhesion, internalization, and intracellular survival of C. jejuni following exposure of these organisms to various stress conditions. We also assessed the ability of C. jejuni to translocate across polarized PSI cl 1 cell monolayers by measuring transepithelial electrical resistance (TER) and detecting translocated bacteria in the basal chamber of a functional small-intestinal cell model.
Materials and Methods
Bacterial growth and stress conditions
C. jejuni K49/4, a poultry meat isolate, was stored at −80°C in 0.6 mL brain–heart infusion broth with 20% blood and 20% glycerol. The organism was grown microaerophilically (5% O2, 10% CO2, 85% N2) on Columbia agar (Oxoid, Hampshire, UK) in Preston broth (Oxoid) at 42°C for 9 hours.
Campylobacters secrete numerous stress protective factors that are present in the exponential growth phase culture (Kelly et al., 2001; Klančnik et al., 2009) and this was the reason for focusing further stress response experiments on exponential phase culture. To produce heat shock, the temperature was shifted from 42°C to 55°C for 3 minutes, and the cells were then cooled on ice before analysis. For multinutrient starvation, bacterial cells were harvested by centrifugation (12,000 g at 4°C for 5 minutes), washed, resuspended in Ringer's solution supplemented with KH2PO4 (5 mM; Kemika, Zagreb, Croatia), and incubated microaerobically for either 5 or 15 hours at 42°C. To produce oxidative stress, bacterial cells were exposed to oxygen at atmospheric concentration for 5 or 15 hours. Untreated C. jejuni, taken from culture at the same time as the stressed organisms, were used as simultaneous controls in all three stress analyses.
Culturability and viability assay
Culturability, measured as colony-forming units per milliliter (CFU/mL), was assessed on Karmali agar plates (Oxoid) after serial sample dilution and microaerobic incubation at 42°C for 24 or 48 hours. Viability was determined with a LIVE/DEAD® BacLightTM viability kit (L-7012; Molecular Probes, Eugene, OR) and examined under an Eclipse TE300 microscope (Nikon, Tokyo, Japan), using a super high-pressure mercury lamp power supply (model HB-101040AF, no. 8701816; Nikon) and X-60 oil-immersion fluorescent objective with blue excitation light (B2A; Nikon) as previously described (Klančnik et al., 2006). A digital camera (DXM 1200; Nikon) and LUCIA 4.60 software (Nikon Programming Equipment Laboratory Imaging, Tokyo, Japan) were used for imaging. All experiments were independently repeated three times and the results are expressed as the number of culturable or viable cells found after a particular treatment (stress condition) when compared with the number of cells in the respective untreated control.
PSI cell line
The normal PSI epithelial-derived cell line PSI cl 1 was obtained from an adult pig at slaughter as previously described (Gradišnik et al., 2006) and was further characterized. Cells were positive for cytokeratins 5–18 as epithelial markers and weakly positive for both alkaline phosphatase and periodic acid-Schiff staining, but negative for cytokeratin-19 and all markers characteristic of myeloid cells, myofibroblasts, or mesenchymal cells. The PSI cl 1 cells were able to form a tightly packed epithelial barrier when grown on microporous inserts with or without collagen. Cells were classified as cryptic and incompletely differentiated enterocytes.
Cells supplemented with 5% fetal calf serum (BioWhittaker Europe, Essen, Germany),
Adhesion, invasion, and intracellular survival
At confluence, the PSI cl 1 cells were trypsinized, and for each assay, 96-well plates were seeded at a concentration of 6 × 104 cells/well and incubated for 24 hours at 37°C in a 5% humidified CO2 atmosphere to obtain confluence at 12 hours. The numbers of bacteria in each well were determined spectrophotometrically at 600 nm before infection of the PSI cl 1 cells. Infection was produced by adding C. jejuni at an approximate multiplicity of infection of 100 to the confluent PSI cl 1 cell monolayer. Infected monolayers were further incubated for 2 hours to allow adhesion and invasion as described previously (Rubeša Mihaljević et al., 2007). The PSI cl 1 monolayers were then washed with DMEM without antibiotics to remove unbound bacteria, after which DMEM containing gentamicin at 100 μg/mL was added for the enumeration of intracellular bacteria. Monolayers were lysed with cold distilled water, and the numbers of intracellular bacteria released by lysis were determined through culturability assays at 24, 72, and 96 hours postinfection. The total number of adherent and internalized bacteria was determined simultaneously with an invasion assay performed without gentamicin treatment (Šikić Pogačar et al., 2009). The difference between the numbers of total and intracellular bacteria was calculated to obtain the number of adherent C. jejuni cells. All experiments were repeated three times and the results for the adherence and invasion analyses were expressed as the number of adherent or internalized bacteria found under the specific treatment conditions (the stress conditions) when compared with the number of adherent or internalized bacteria found under the respective untreated control conditions. The log10 CFU/mL of internalized bacteria was used to represent intracellular bacterial survival.
Functional epithelial cell model
To obtain polarized epithelial cell monolayers, we seeded PSI cl 1 cells onto Transwell® filter inserts (0.4 μm pore size, 12 mm diameter; Corning, Corning, NY) placed into 12-well plates (Corning) at a density of 1 × 105 cells/cm2. A volume of 500 μL of cell growth medium (DMEM with antibiotics) and 2% (or 10 μL) fetal bovine serum was added to the inner chamber (apical compartment) of this epithelial cell model and 1500 μL of cell growth medium was added to the outer chamber (basal compartment) of the model. Both the apical and basal media were changed every 2 days until functional polarity was achieved. Functional polarity was defined as having been achieved when the electrical resistance between the apical and basal surfaces of the PSI cell monolayers exceeded 1600 Ω/cm2. At this point we performed the C. jejuni infection assay, which was followed by a 2-hour incubation at 37°C in a 5% humidified CO2 atmosphere to allow adhesion and invasion.
TER
TER was measured with the Millicell-ERS Electrical Resistance System (Millipore, Bedford, MA). The net value of TER (Ω/cm2) was corrected for background resistance by subtracting the resistance value of the cell-free microporous membrane and that of the medium (108 Ω/cm2) from the resistances measured with the system. TER was measured before the addition of bacteria to the medium, with or without gentamycin, at day 7 and on each subsequent day until 12 days after infection. The TER values of cell monolayers without bacteria were used as the control values for each experiment. To determine the translocation of unstressed C. jejuni across the cell barrier, cell culture medium taken from the basal chamber of the model at different times after infection was inoculated onto agar plates and tested for C. jejuni.
Statistical analysis
We compared the bacterial count data for different experimental groups by using one-way analysis of variance and Tukey's b-tests. All statistical values were considered significant at p ≤ 0.05. Statistical analysis was performed with the Statistica for Windows software system version 6.1 (Statsoft, Tulsa, OK).
Results and Discussion
The interactions between bacteria and intestinal cells create the basis for disease pathology. After assessing the survival of C. jejuni under conditions of starvation, heat, or oxidative stress, we studied the organisms' adhesion to and invasion of PSI epithelial cells (PSI cl 1), the model closest to humans in terms of genome, organ development (Lunney, 2007), anatomy, physiology, and metabolism of the intestinal tract (Baltzell et al., 1987; Brown and Timmermans, 2004), disease progression (Lunney, 2007), and intestine–microbe interactions (Pipenbaher et al., 2009). Moreover, these cell lines are not of tumor origin and are therefore a better in vitro model for studying the mechanisms of pathogen–host interactions than human tumorogenic cell lines (e.g., CaCo2). We also measured the TER of polarized epithelial cells as an indicator of cell monolayer integrity and translocation of C. jejuni across the polarized intestinal epithelial cell monolayer.
Survival and pathogen–host cell interactions of C. jejuni
Kinetic analyses of the attachment of C. jejuni to intestinal epithelial cells indicate that stress in general reduces the ability of these bacteria to attach to the surface of and invade these cells. Exposure to all of the stresses examined in our study, other than exposure to oxygen for 5 hours, reduced the ability of C. jejuni to enter the intestinal epithelial cells as shown by the invasion assay used in our study (Fig. 1). Microaerophilic campylobacters are already exposed to oxidative stress in the environment outside the host, as, for example, in poultry meat processing. In the host, campylobacters are additionally exposed to oxidative stress created by macrophages involved in the host immune response. However, the experimental exposure of C. jejuni to oxygen in our study did not reduce either the organisms' culturability or viability (Fig. 1). The survival of C. jejuni inside the intestinal epithelial cells was not effected if the bacteria were exposed to atmospheric oxygen concentration for only a short period (i.e., 5 hours) (Fig. 2). In our previous work, we observed significant intraepithelial survival of C. jejuni in CaCo2 and murine macrophage cell lines (Rubeša Mihaljević et al., 2007; Pogačar et al., 2009). This work showed that bacterial mechanisms involved in inactivating toxic oxygen products enhanced bacterial persistence within the host (Rubeša Mihaljević et al., 2007; Šikić Pogačar et al., 2009). Our results with pig intestinal cells show once again that the ability of C. jejuni to survive in cells that these organisms have invaded depends upon conditions preceding the presence of stress (Fig. 2). The rate of survival of bacteria shortly starved or exposed to oxidative stress before their invasion of PSI cl 1 cells was not strongly reduced. However, in cells oxidatively stressed for 10–15 hours, the number of invading C. jejuni cells were below the detection limit of the assay (Fig. 2). Although most work to date has focused on the entry of C. jejuni into host cells, the intracellular fate of this pathogen has been largely ignored. Our results are consistent with the study by Day et al. (2000), which confirmed the ability of C. jejuni to survive intracellularly for relatively long periods both in phagocytes (72 hours) and in intestinal epithelial cells (96 hours), persuading some investigators to classify this organism as a facultative intracellular pathogen as suggested by Day et al. (2000).
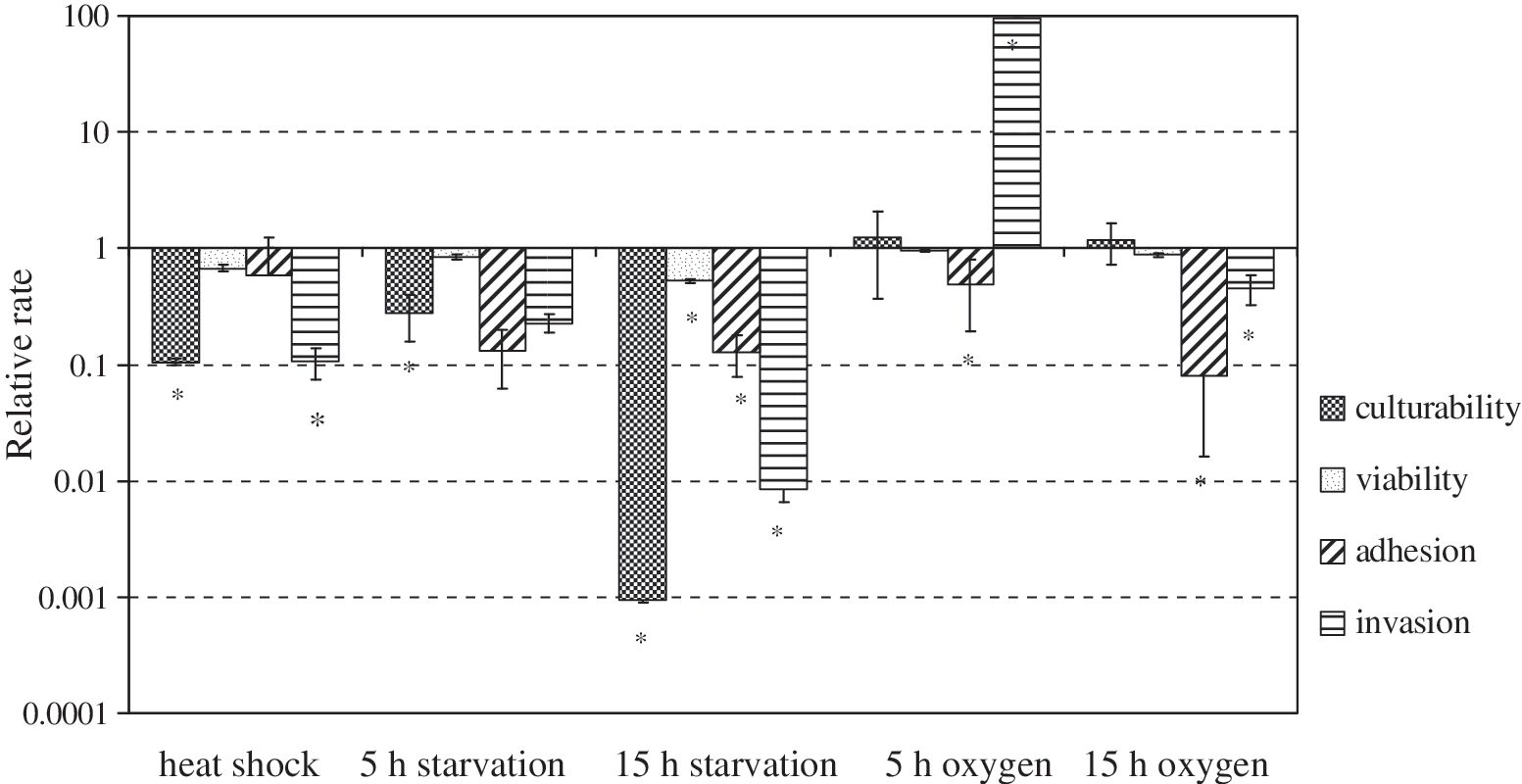
Culturability, viability, adherence, and invasiveness of stressed Campylobacter jejuni in PSI cl 1 cells. The results for each experimental variable are presented as mean ± SD (* p ≤ 0.05). PSI, pig small-intestinal; SD, standard deviation.

Intracellular survival of unstressed (control) and stressed C. jejuni in PSI cl 1 cells at the time intervals indicated. Each point represents the mean ± SD of log10 colony-forming units (CFU)/mL.
Campylobacter spp. encounter dramatic temperature changes during the infection process, which could also influence their survival. As shown in Figure 1, loss of growth of the C. jejuni used in our study was observed after heat shock, although the organisms' viability was not affected. When compared with their attachment and invasiveness in a previous study (Rubeša Mihaljević et al., 2007), the ability of C. jejuni to attach to and invade the CaCo2 cells in this study was strikingly lower than the same properties observed with PSI cl 1 cells. This is not surprising, in that CaCo2 cells originate from a human colon adenocarcinoma. It is well known that cancer cells express modified surface glycoconjugates, among other genotypic and phenotypic differences between these cells and normal cells such as PSI epithelial cells, which make CaCo2 cells inappropriate as a model for pathogen–host cell interactions. Moreover, CaCo2 cells derived from colon tissue differ from epithelial cells of the small intestine (e.g., PSI epithelial cells), where most in vivo host–pathogen interactions occur. As shown in Figure 1, starvation affected the culturability and viability of C. jejuni and reduced the number of adherent and internalized bacteria. The 15-hour starvation, in particular, greatly diminished bacterial attachment and invasiveness and caused a rapid loss of intracellular bacterial growth (Figs. 1 and 2). These findings are consistent with previous studies of C. jejuni done with CaCo2 cells and murine macrophages (Rubeša Mihaljević et al., 2007; Šikić Pogačar et al., 2009), indicating that prolonged nutrient limitation reduced the adherence, invasiveness, and intracellular survival of these organisms. We also previously found that nutrient limitation induced cross protection of C. jejuni against heat stress and increased intracellular oxidative stress and influenced these organisms' adhesion, invasiveness, and intracellular survival both in vitro and in vivo (Klančnik et al., 2008; Vučković et al., 2008).
Intercellular intestinal epithelium integrity and translocation capacity of C. jejuni
Although little is known about the interaction of C. jejuni with cultured epithelial cells, the influence of bacteria–host cell interaction on intestinal epithelial cell polarity has recently been pointed out (Beltinger et al., 2008; Van Deun et al., 2008). The TER of polarized epithelial cells is an indicator of intercellular integrity. Translocation of C. jejuni by a paracellular route would be expected to disrupt the junctional complex between epithelial cells. This would increase monolayer permeability and reduce the TER. The ability of enteric pathogens to translocate across the intestinal cell barrier is a prerequisite for infection (Konkel et al., 2001; Klingberg et al., 2005) and is therefore considered an important virulence attribute. The mechanism by which C. jejuni translocates across polarized epithelial cells remains unclear. To assess the effect of C. jejuni on epithelial intercellular integrity, we measured the TER immediately after infection of polarized PSI cl 1 cells at day 7 (Fig. 3). Following attachment of C. jejuni to the polarized PSI cl 1 cells, we observed a decrease in TER in the first 24 hours after infection and again at 72 hours after infection, reflected by a decrease of almost 1000 Ω in values of TER at day 11 (Fig. 3). Adherent C. jejuni affected the cell monolayer integrity more than did invading bacteria, as shown by TER values (Table 1). This latter effect could be argued to have occurred as a consequence of replication of C. jejuni at the cell surface and translocation of these organisms into the intestinal epithelial cells used in this component of our study. We believe that by increasing the number of bacteria at the cell surface, disturbance of metabolic activity in the gut microenvironment may enhance disruption of the intercellular integrity of the polarized intestinal epithelium. Our results are in accord with those of Bras and Ketley (1999), who concluded that a disturbance in TER during the first 24 hours after entry of C. jejuni into the gut is the result of these bacteria using a transcellular rather than a paracellular route of translocation, as the TER during this interval shows only slight changes. On the other hand, only slight changes in TER could be noticed following invasion by C. jejuni of the epithelial cell monolayer in our study. These results imply that C. jejuni, once inside the host cell, have no effect on intercellular integrity and could disseminate through the intestinal barrier by a transcellular route. We found no correlation between the variations in the resistance values of our model of the intestinal epithelium and the ability of C. jejuni to translocate through the polarized intestinal epithelial cell monolayer in the model. Further, we detected bacteria in the basal chamber of our model at 3 hours after infection of polarized PSI cl 1 cells (Table 1). Similar observations were reported by Konkel et al. (1992), who found a maximum rate of translocation at 4 hours after the infection of polarized CaCo2 cells. Our present report describes the effect of different stress conditions on the course of infection of PSI cl 1 cells by C. jejuni. Further, the mechanisms that C. jejuni uses to enter host intestinal epithelial cells are most likely transcellular, in view of our demonstration that TER was not significantly disrupted after the translocation of these bacteria through the host intestinal epithelial cells in our model.
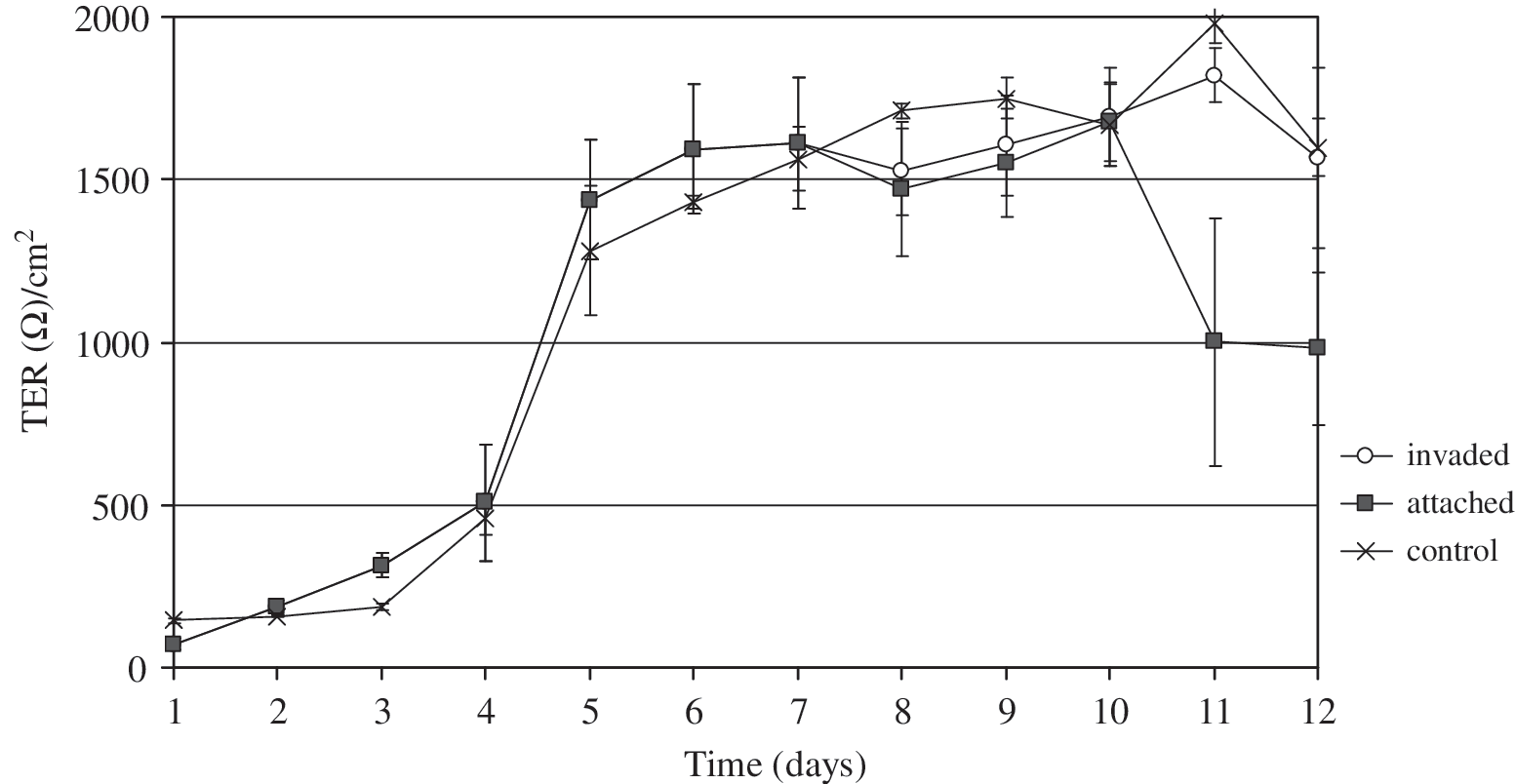
Transepithelial electrical resistance (TER) of polarized PSI cl 1 cell monolayers exposed to 1 × 108 bacteria/mL when compared with the TER with unstressed bacteria (control) in terms of adherent and invasive C. jejuni. Bacteria were added to the apical side of polarized monolayers on day 7, when the maximum TER was measured. Each point represents the mean ± SD of the TER in Ω/cm2.
The TER of pig small-intestinal cells without bacterial inoculation (control) was 1612 ± 207 Ω/cm2 and remained almost unchanged during the 120-hour experimental period.
CFU, colony-forming units; NM, resistance not measured.
Conclusions
C. jejuni can adapt to environmental oxidative stress in host and modulate its intestinal epithelial cell interactions with these bacteria, such as adherence, invasion, and intraepithelial survival, allowing C. jejuni to translocate to the subepithelial mucosa. In our study, exposure to atmospheric oxygen for a short period increased the invasiveness and intraepithelial survival of C. jejuni. On the other hand, heat stress, and more significantly nutrient limitation, reduced the adherence, invasiveness, and intracellular survival of C. jejuni. Adherent C. jejuni had a significant disruptive effect on the TER of the intestinal epithelium, as shown by a marked decrease in TER values in our polarized intestinal epithelial cell model when compared with that for bacterially invaded epithelium. Our new functional model of the intestinal epithelium, which accords with the properties of normal, noncarcinogenic small intestinal epithelial cells (PSI), has proven significantly more accurate for the study of the interactions of foodborne pathogens with the gut epithelium than have models using carcinogenic cells (e.g., CaCo2 cells). This model will allow the additional studies needed to determine the exact mechanism by which C. jejuni establish intestinal infection in vivo.
Footnotes
Acknowledgments
This work was supported by EU-FP6 research project PathogenCombat-FOOD-CT-2005-007081 and by bilateral Croatian–Slovenian project “Resistance and virulence of Campylobacter species.” The authors thank Tanja Botić, a Ph.D. student at the University of Maribor, Faculty of Agriculture and Life Sciences, for her technical assistance and Dr. David Fenn Oldmeadow at the University of Maribor, Faculty of Agriculture and Life Science, for English revision of the manuscript.
Disclosure Statement
No competing financial interests exist.