
Abstract
Thirteen strains of Mucor circinelloides and nine strains of Rhizopus stolonifer were collected from different Saudi fruits. Also, two strains of M. circinelloides and one strain of R. stolonifer isolated from soil were used in this study to analyze the genetic diversity among these 25 strains. The dendrogram constructed from random amplified polymorphic DNA–polymerase chain reaction results using two primers (V6 and M13) showed no correlation between clustering system and sources of isolation for M. circinelloides strains, while Rhizopus strains clustered according to the sources of isolation. The phylogenetic tree based on the unweighted pair group method with arithmetic average of internal transcribed spacer (ITS) rDNA sequence revealed the variation of the ITS region among Mucor strains as well as Rhizopus strains.
Introduction
Mucor and Rhizopus species cause the postharvest decay of a variety of fruits and vegetables. In various instances, the Mucor species have been reported to have caused serious losses in strawberries, pears, apples, peaches, nectarines, guavas, tomatoes, sweet potatoes (Michailides, 1991), mangoes, and various vegetables (Smith et al., 1979). Rhizopus rot caused by R. stolonifer is one of the most destructive diseases of stone fruits (Nishijima et al., 1990; Sholberg et al., 1995; Waller et al., 2001; Howell, 2002; Zhang et al., 2004). Rhizopus soft rot caused by R. stolonifer and R. oryzae is the most serious and widespread postharvest disease of sweet potatoes in the United States and worldwide (Holmes and Stange, 2002).
Molecular genetic approaches are now generally used for filamentous fungi too, and are fundamental for modern taxonomy (Kurtzman et al., 1986; Tamura et al., 1999; Voigt et al., 1999; O'Donnell et al., 2001; Binder and Hibbett, 2002; Moncalvo et al., 2002; Lutzoni et al., 2004). Molecular techniques using genomic targets within the rRNA complex have been shown to be reliable for the species identification of the zygomycetes, including Mucor species (Iwen et al., 2002, 2005; Rakeman et al., 2005; Schwarz et al., 2006). RAPD analysis offers a rapid and sensitive method with which to acquire detailed information about the type and level of genetic variability (Williams et al., 1991). This approach has been used efficiently for the genotypic analysis of several groups within the Zygomycetes (Burmester and Wöstemeyer, 1994; Bidochka et al., 1995; Abbas et al., 1996; Nelson et al., 1998; Vastag et al., 2000; Vágvölgyi et al., 2001). Taxonomic revision of the genus Rhizopus has led to several independently described species made being obsolete (such as R. reflexus, R. circinans, and R. niveus) on morphological and physiological considerations. They are now differentiated as only two varieties within the same species: R. stolonifer var. stolonifer and R. stolonifer var. reflexus (Schipper, 1984). In contrast with this situation, the RAPD analysis of 29 R. stolonifer strains isolated from various locations and substrates revealed a high genetic heterogeneity within the species R. stolonifer. This suggesting practically species-level differences between the isolates of R. stolonifer var. stolonifer and R. stolonifer var. reflexus (Vágvölgyi et al., 2004).
Epidemiological investigations and control of these infections require reproducible methods of distinguishing among isolates and revealing the genetic make-up of a fungal species. On the other hand, little is known about the genetic variability of these organisms, and the compositions of their populations are poorly understood. Therefore, the purpose of the present study is to characterize Mucor circinelloides and R. stolonifer species isolated from some Saudi fruits based on RAPD and ITS region of rDNA sequences to evaluate the validity of the taxonomic classification by morphological characteristics.
Materials and Methods
Fungal strains and cultivation
Twenty-two strains of Zygomycetes (13 M. circinelloides and 9 R. stolonifer) isolated from different Saudi fruits (apricot, date, grapes, and plums) were used in this study. Two strains of M. circinelloides and one strain of R. stolonifer isolated from soil were used to confirm the relation between source of isolation and DNA fingerprinting patterns. Mycelium for DNA isolation was grown in YM broth (0.3% yeast extract, 0.3% malt extract, 0.5% peptone, 2% dextrose; Difco, Detroit, MI) at room temperature for 2 to 5 days. Strains grown on YM agar (2% Difco agar) were examined morphologically according to the method of O'Donnell (1979) to confirm their identity.
DNA extraction
After 5 days mycelium was collected by vacuum filtration and ground to a fine powder in liquid N2. Fifty milligrams of the powder was transferred to 1.5 mL Eppendorf tube and mixed with 700 μL CTAB buffer (0.1 M Tris–HCL, 20 mM EDTA, 1.4 M NaCl, 2% cetyltrimethylammonium bromide, and 0.2% mercaptoethanol, pH 8.0). The tubes were incubated at 65°C for 30 min, and then 700 μL of chloroform was added and the mixture vortexed briefly. The resulting mixture was centrifuged at a maximum speed of 15,000 rpm for 30 min and the cleared supernatant was mixed with 600 μL of isopropanol chilled to 72°C. The mixture was centrifuged at the maximum speed for 5 min and the resulting pellet washed twice with 1 mL of 70% ethanol. The pellet was dried under vacuum and dissolved in 100 μL TE buffer (10 mM Tris, 1 mM EDTA, pH 7.5). The DNA was observed by electrophoresis in 1% agarose gel in Tris Borate EDTA buffer: 89 mM Tris–HCl (pH 8.3), 89 mM boric acid, and 20 mM EDTA (Maniatis et al., 1982). Agarose gels were stained with ethidium bromide and examined under ultraviolet light.
Random amplified polymorphic DNA–polymerase chain reaction
Polymerase chain reaction (PCR) conditions and separation of random amplified polymorphic DNA (RAPD)–PCR fragments were carried out according to the techniques of Messner et al. (1994). PCR was carried out using primer V6 (TGCAGCGTGG; Prillinger et al., 1999) and M13 (GAGGGTGGCGGTTCT; O'Donnell et al., 1999), respectively. The following temperature protocol was used: initial denaturizing step of 5 min at 95°C, 40 cycles of denaturation for 15 sec at 95°C, annealing for 90 sec at 40°C, and extension for 100 sec at 72°C. The PCR products were resolved by electrophoresis by using 1.4% agarose gel in 0.5 × Tris Borate EDTA buffer, at 125 V for 2 h. Gels were stained with ethidium bromide and viewed under UV light using UVP BioImaging CDS 8000 System (UVP, Cambridge, UK). Dendrograms for RAPD results were constructed by the unweighted pair group method with arithmetic average based on Jaccard's Similarity Coefficient by using Phoretix 1D Advanced v5.20 software (Non-Linear Dynamics).
ITS sequencing
Ribosomal DNA including the complete ITS1-5.8S-ITS2 region was amplified with ITS1 (TCCGTAGGTGAACCTGCGG
Sequence analysis
DNA sequences were aligned with CLUSTAL Software. The Treecon program (Van de Peer and De Wachter, 1994) was used to carry out rooted parsimony analysis for investigating the tree topology. Support for the branches based on parsimony criteria estimated by bootstrap analysis of 100 replicates were generated with this program.
Results and Discussion
Twenty-five species of Zygomycetes were collected in this work (Table 1). Fifteen isolates of M. circinelloides and 10 isolates of R. stolonifer were collected from different fruits and from soil as well. Michailides (1991) isolated M. circinelloides from stone fruits from California and Chile. Also, Eseigbe and Bankole (1995) isolated this specie as associated fungus with postharvest rot of black plum (Vitex doniana) in Nigeria. R. stolonifer has a broad host range: many fruits and vegetables are susceptible to this pathogen—peaches, nectarines, sweet cherries, strawberries, and plums are the most susceptible. Some Mucor and Rhizopus species have been described as an important agent of postharvest fungal infections (Smith et al., 1979; Michailides, 1980), causing decay in highwater-content fruits (Lopatecki and Peters, 1972) and vegetables (Moline and Kuti, 1984). These infections can be significant when fruits are kept at temperature between 0°C and 20°C (Michailides, 1980).
RAPD–PCR for M. circinelloides isolates
Using two different primers, each isolate recorded considerable numbers of DNA patterns. The numbers of bands for M13 primer fluctuated from 11 to 17 bands. The recorded bands showed molecular weight variation from 188 to 3135 bp. On the other hand, V6 primer gave numbers of bands ranging from 13 to 19 with molecular weight ranging from 271 to 3163 for its bands. The results of M13 and V6 primers were used to construct dengrogram showing the relationships among Mucor circinelloide isolates (Fig. 1). RAPD analysis of 30 isolates of M. piriformis, collected from infected fruit, revealed 19 composite amplification types (Papp et al., 1997), indicating a much higher degree of variability than found in previous isoenzyme studies (Vágvölgyi et al., 1996). The dendrogram indicated that there is no correlation between clustering system and sources of isolates. For example, TUM2, TUM5, TUM6, TUM7, TUM8, TUM10, and TUM11 clustered together although they were isolated from different fruits (Fig. 1). This results may be attributed to the broad host range of M. circinelloides. Papp et al. (1997) reported that the numerical analysis of M. piriformis revealed three clusters, which correlated with the mating competency of the isolates or their place of origin. Their results demonstrated that RAPD analysis can identify isolates and subspecific populations of M. piriformis. A similar observation has been made with Gilbertella persicaria isolates from infected peaches and nectarines; in this case, reproducible RAPD markers were obtained which correlated with the mating types (plus or minus) of the isolates (Papp et al., 2001).
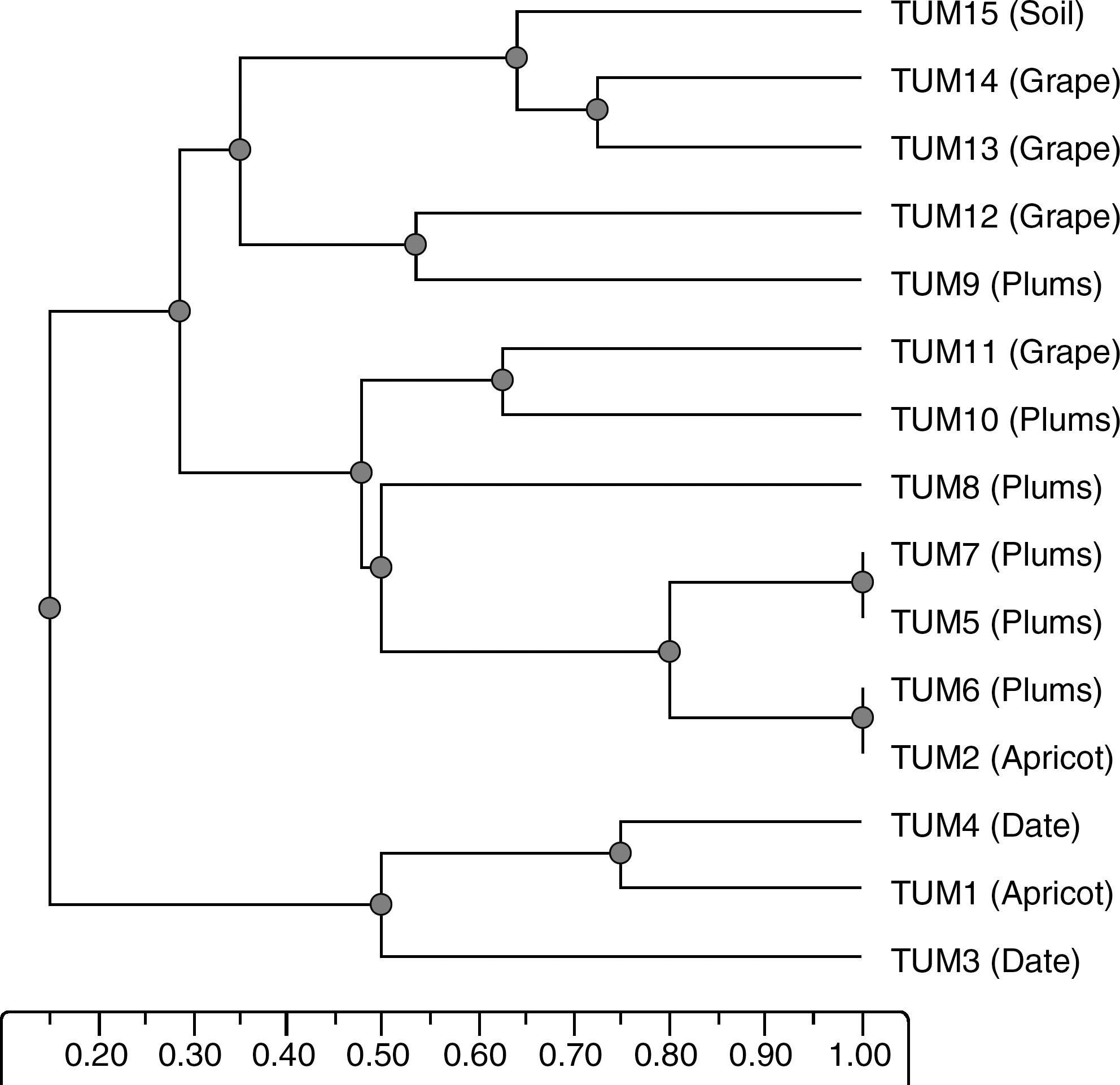
Dendrogram showing relationships among 15 strains of Mucor circinelloides isolated from different fruits (in parentheses) and soil. Genetic distances were obtained by random amplified polymorphic DNA analysis using two different primers (V6 and M13).
RAPD–PCR for R. stolonifer isolates
The number of amplified bands with M13 primer varied between 15 and 18 with amplification product between 300 and 2754 bp. On the other hand the number of amplified bands varied between 12 and 18 and sizes of the detected amplification products were between 297 and 2607 bp with V6 primer. Different primers revealed different degrees of DNA polymorphism among the strains of R. stolonifer. Also, strains TUR1, TUR2, TUR3, and TUR4 isolated from apricot and comprised one cluster. Meanwhile TUR9 and TUR10 (isolated from soil) cluster together as shown in Figure 2. Vágvölgyi et al. (2004) assessed genetic variability in R. stolonifer, by RAPD analysis. They reported that the numerical analysis of the RAPD data revealed four main clusters with extremely high dissimilarity values, but only low or moderate variability was observed within these groups. Their results suggest a high genetic heterogeneity in the case of R. stolonifer: isolates of R. stolonifer var. stolonifer, R. stolonifer var. reflexus, and R. niveus displayed species-level genetic distances, which gives rise to considerations that they might be separate species. A dendrogram was constructed from these data by using unweighted pair group method with arithmetic average method. The R. stolonifer strains formed three different clusters. The clustering system followed the source of isolation. Strains TUR6, TUR7, and TUR8 isolated from grape and constituted one cluster (Fig. 2). Vágvölgyi et al. (2004) reported that three R. stolonifer isolates isolated from infected nectarines and peaches belonged to one cluster. These isolates exhibited similar morphological characteristics and growth requirements as other R. stolonifer isolates, although they displayed a very few common RAPD markers with isolates in the other clusters. The work by Vágvölgyi et al. (2004) supports the need for a reconsideration of the original taxonomic distinction.
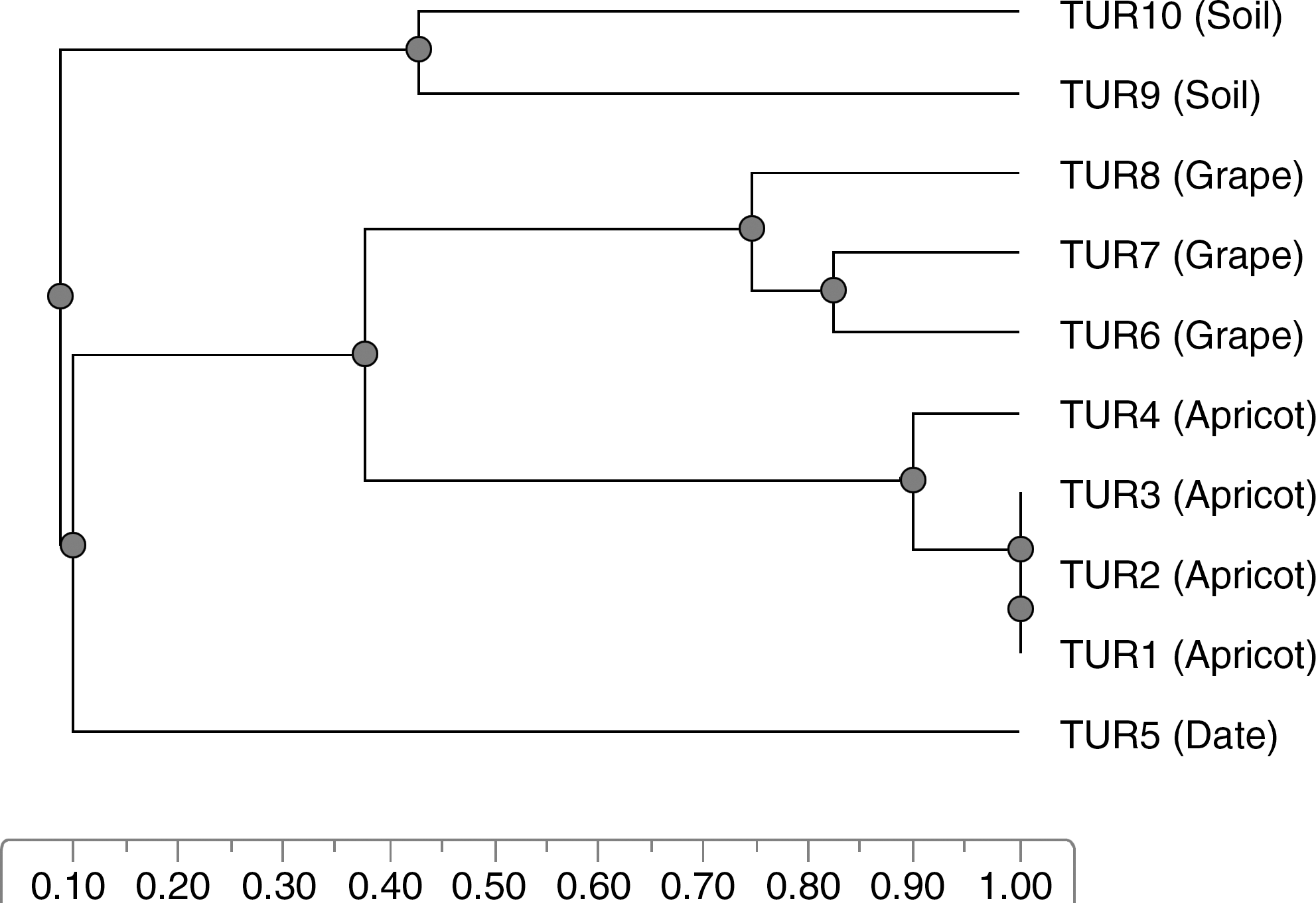
Dendrogram showing relationships among 10 strains of Rhizopus stolonifer isolated from different fruits and soil. Genetic distances were obtained by random amplified polymorphic DNA analysis using two different primers (V6 and M13).
Sequencing
Since TUM2 and TUM6; TUM5 and TUM7; and TUR1, TUR2, and TUR3 were completely identical according to their RAPD results; TUM6, TUM7, and TUR1 were used as representative strains for sequencing. The other isolated strains were applied for sequencing analysis. The phylogenetic tree was made with those data and some known sequences from the databases (Fig. 3). Among these DNA sequences, a major difference that could divide the sequences into two groups was detected (Fig. 3). According to the phylogenetic tree constructed from the sequences results there were two different clades; Mucor clade and Rhizopus clade.
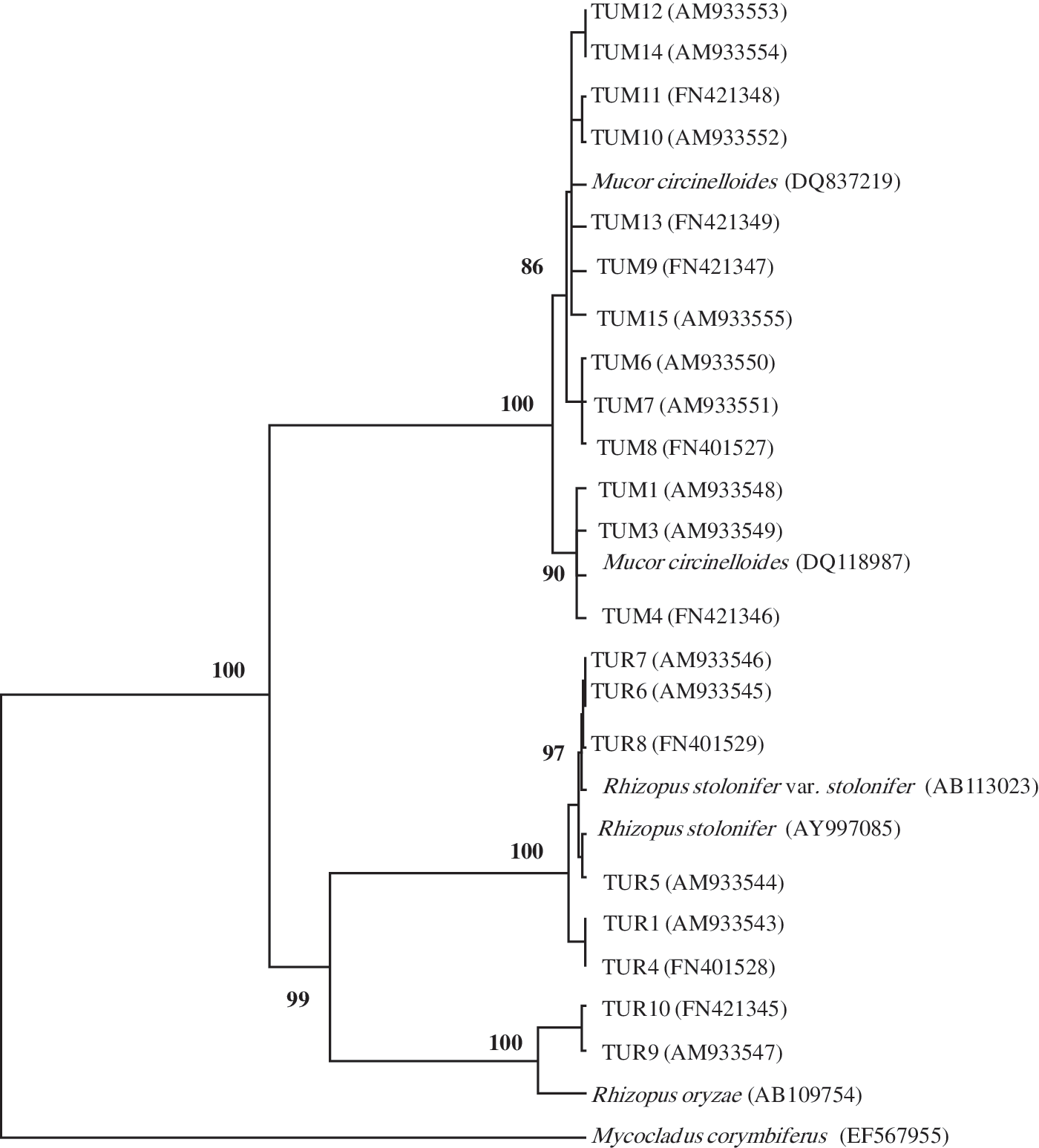
The unweighted pair group method with arithmetic average phylogenetic tree or rDNA ITS sequences of Mucor and Rhizopus strains. Bootstrap percentage over % from 100 replicates are shown in each branch. The number in parentheses indicated their database accession numbers.
Mucor clade was divided into two subclades. The first subclade comprised of seven strains (from TUM9 to TUM15). These strains clustered together with M. circinelloides (DQ837219) from GenBank with 79 bootstrap values. On the other hand, the second subclade included six strains (TUM1, TUM3, TUM4, TUM6, TUM7, and TUM8) and Mucor circinellonides (DQ118987) from GenBank with 80 bootstrap values. These results indicated the variation of ITS rDNA in M. circinelloides strains. Nagy et al. (2000) reported that for the M. circinellonides and Mucor racemosus strains, extensive chromosome-length polymorphism was observed, whereas the karyotypes of the Mucor plumbeus were much more similar. This phenomenon may somehow reflect that both M. circinellonides and M. racemosus species unify several species formae in the current taxonomic approach, while M. plumbeus is a single specie with well-defined morphological characteristics (Schipper, 1976). This explanation is supported by the results of earlier isoenzyme studies on 27 Mucor strains; for M. circinellonides and M. racemosus, much higher levels of intraspecific variability were found than for M. plumbeus (Varga and Vágvölgyi, 1991). Schipper (1976) described four formae (M. circinelloides f. circinelloides, M. circinelloides f. janssenii, M. circinelloides f. griseocyanus, and M. circinelloides f. lusitanicus) based on differences in the shapes of spores and columellae and in thermotolerance. Schwarz et al. (2006), and confirmed by Iwen et al. (2007), showed that M. circinelloides demonstrates variability within the ITS regions, potentially supporting subspecies groups.
Rhizopus clade was divided into two subclades: R. oryzae group subclade and R. stolonifer subclade. The first one comprised of strains TUR9 and TUR10 that had the same sequence of Rhizopus oryzae AB109754. These strains were identified mistakenly (based on morphological criteria) as R. stolonifer but it is clear now that they were R. oryzae according to the sequence results. Vágvölgyi et al. (2004) reported that isolates of R. oryzae from very different origin formed a well-defined group, supporting the unity of this species.
R. stolonifer subclade included strains TUR1 and TUR4 that clustered together with R. stolonifer AY625072 with 69 boostrap values. While TUR5 and TUR8 clustered together with R. stolonifer AY997085 with 89 boostrap values. This clustering system supports the heterogeneity of the ITS region among R. stolonifer strains. Schipper and Staplers (1984) divided the genus Rhizopus into three groups; the stolonifer, oryzae, and microsporus groups based on the general morphology and mating experiments. However, the morphological and physiological characteristics of these species tend to overlap species lines. Our results came in agreement with Abe et al. (2006) findings. They sequenced three molecules of the ribosomal RNA encoding DNA (rDNA), complete 18S, ITS1-5.8S-ITS2, and 28S D1/D2 regions of all the species of the genus Rhizopus. Their phylogenetic trees showed three major clusters corresponding to the three groups in the current morphological taxonomy: microsporus group, stolonifer group, and R. oryzae.
In conclusion our results indicate the variation of ITS region sequence among M. circinelloides strains and also possibility of the molecular identification of the Rhizopus species groups using rDNA sequencing.
Footnotes
Acknowledgment
This work was supported by a grant (Contract No. 51-428-1) from Taif University, Kingdom of Saudi Arabia.
Disclosure Statement
No competing financial interests exist.