
Abstract
We examined the phospholipids (Phls) and the membrane fatty acid (FA) composition in Salmonella enterica serovar Typhimurium dam and/or seqA mutants. Phosphatidylglycerol, phosphatidylethanolamine (PE), and cardiolipin (CL) are the major Phls present in all the strains and accounted for greater than 95% of the total lipid phosphorus. Phosphatidic acid and phosphatidylserine are the minor ones. The seqA mutant showed a decrease in PE and an increase in CL and phosphatidylglycerol proportion compared with the wild-type strain. The same changes were observed with the seqA dam double mutant. However, the dam mutation caused an unusual accumulation of CL with a significant decrease in the PE content, compared with the isogenic wild-type strain. FA composition of the total lipids and the different fractions containing Phls have been determined. The major saturated FAs (SFAs) and unsaturated FAs (UFAs) found were C14:0, C16:0 and C16:1w7, C18:1w9, respectively. Cyclic FAs, cyc17:0 and cyc19:0, were also present in appreciable amounts. Moreover, dam and/or seqA mutations caused a decrease in UFA/SFA ratio and there was a progressive reduction in the content of C16:1w7 and C18:1w9, going through the order seqA, dam/seqA, and dam mutants. This decrease in UFA content was compensated for in all strains by an increase in the corresponding C17− and C19− cyclic FAs. So these UFAs were converted to their cyclopropane derivatives, which resulted in a low UFA/SFA ratio. SeqA and Dam proteins might regulate FA biosynthesis and Phls composition of Salmonella enterica serovar Typhimurium.
Introduction
S
Materials and Methods
Bacterial strains
The strains of S. enterica used in this study belong to serovar Typhimurium and are derived from the mouse-virulent strain American Type Culture Collection 14028. Four isogenic strains were used and their relevant genotypes are listed in Table 1. The strains were kindly provided by Dr. Fransisco Ramos-Morales (Departamento de Genética Facultad de Biología Universidad de Sevilla, Spain).
WT, wild type.
Culture conditions
Pure cultures of bacterial strains were maintained on nutrient agar plates at 4°C. Precultures were prepared by transferring an isolated colony from a plate into a test tube containing 10 mL of sterile NB followed by aerobic incubation in a shaker water bath at 37°C for 24 h. Erlenmeyer flasks containing 50 mL of sterile NB were inoculated with the subculture to a final concentration of 103 cells/mL. When necessary, liquid medium was supplemented with kanamycin to a final concentration of 50 μg/mL. The optical density of the cultures and the dry-weight contents were assayed as described by Ames (1968).
Extraction of total lipids
The lipid extraction procedure was previously described (Bligh and Dyer, 1959). The method consists of extraction of the lipids in a monophasic system in which methanol, chloroform, and water were in the proportions 2:1:0.8 (vol:vol:vol). The lipids were separated from the water-soluble material by diluting the extraction mixture with one volume of chloroform followed by one volume of water. After centrifugation, the chloroform layer was removed completely by gently inserting a pipette through the water–methanol phase and through the pellet that was formed at the interphase. The extractions were performed at 0°C. Chloroform layers so obtained were evaporated to dryness in a rotary evaporator at 40°C or less. The lipid residue was immediately dissolved in a small volume of hexane and stored at −18°C under a nitrogen atmosphere until chromatographic analyses were carried out.
Thin-layer chromatography of lipids
The extracted lipids were separated on precoated thin-layer chromatography (TLC) silica gel plates (Sigma, St. Louis, MO). The plates were impregnated and developed in methanol shortly before sample application and then activated by heat treatment at 110°C for 30 min. The chromatographic tanks were lined with filter paper that was thoroughly impregnated with the freshly made solvent immediately before starting development. A precise amount of lipid could be applied, chromatographed, and identified by cochromatography with various commercial authentic standards (Sigma). A first one-dimensional ascending development was seen in the following solvent system: chloroform–acetone–methanol–glacial acetic acid–water (50:20:10:10:5, vol:vol:vol:vol:vol) (Tremoliers and Lepage, 1971). The plates were allowed to dry at room temperature for about 5 min and used for a second one-dimensional ascending development (for a best resolution). Phls were unspecifically visualized with iodine vapors and their rate of flow (R F) were calculated and compared with the known standards. The corresponding Phl bands were marked and scrapped off the plates; the Phls were eluted and their transmethylated FAs were analyzed using a GC system.
Analysis of FA in total lipids and Phls
These analyses were carried out from 1 L of each bacterial culture obtained as previously described. The different bacterial strains were grown to OD600 of 0.5 at 37°C, cells were centrifuged in exponential phase at 4°C for 10 min at 4000 g (Beckman coulter), washed with 1% NaCl buffer, and total lipids (TLs) and Phls were extracted as described earlier. Methylation of the FAs to yield their FA methyl esters (FAMEs) was achieved according to the method described by Cecchi et al. (1985). An aliquot of the solution was evaporated; then 2 mL of hexane, a known quantity of heneicosanoic acid methyl ester [C21:0] as an internal injection standard, and 0.5 mL of sodium methylate (1%) were added. After stirring for 1 min and allowing it to stand for 2 min, the mixture was neutralized by 0.2 mL of H2SO4 (1 N); then, the methyl esters were washed with 1.5 mL of distilled water. The upper phase solvent containing the FAMEs was removed under vacuum with a speedvac system. The membrane FA composition of the bacterial strains was determined on a GC system (6890NR model [Network GC System]; Agilent Technologies, Wilmington, DE) equipped with a flame ionization detector and an electronic pressure control injector. A polyethylene glycol fused silica capillary column (Innowax, 30 m × 0.25 mm × 0.25 μm film thickness; Agilent Technologies) was used. The column was operated at 150°C for 1 min, the temperature was raised by 15°C/min to 210°C for 5 min and then raised by 5°C/min to 250°C and maintained until the end of analysis (25 min). N2 was used as the carrier gas at a flow rate of 150 kPa, H2 at a flow rate of 25 mL/min, and the split ratio used was 60:1. Peak areas were quantified using chromatography software (ChemStation FamilyR data analysis; Agilent Technologies). The FAMEs were first identified by comparing their retention times with those of known standards, and results were expressed as relative percentages of each FA, which were calculated as the ratio of the surface area of the considered peak to the total area of all peaks. Statistical analysis was carried out using Student's t-test (p < 0,05).
Results
Phl components of S. enterica serovar Typhimurium
The Phls were separated and identified by TLC. The content of each bacterial strain Phl was calculated from the FA contents measured by the capillary GC method and is reported in the following section. Figure 1 shows representative one-dimensional separations of the crude lipid extract. The neutral lipids (spot NL) run with the front in the solvent system. The Phls found were phosphatidylethanolamine (PE), PG, and CL on the basis of the following criteria: (i) identity of chromatographic behavior in TLC with synthetic and purified commercial Phls from various sources; (ii) the R F is the same as that of commercial Phls; and (iii) the Phls are the same as those reported by several authors and works (Ames, 1968).

One-dimensional thin-layer chromatogram of total lipids from isogenic Salmonella enterica serovar Typhimurium strains. Development was in the vertical direction in the following solvent system: chloroform–acetone–methanol–glacial acetic acid–water (50:20:10:10:5, vol/vol/vol/vol/vol), with authentic standards. Approximately 1 mg of lipid was applied at the origin. The spot named NL is the neutral lipid. Spot PE was identified as phosphatidylethanolamine, spot PG was identified as phosphatidylglycerol, and spot CL was identified as cardiolipin.
Phl composition of S. enterica serovar Typhimurium wild-type and mutants' membrane
We analyzed Phl composition of the bacterial membrane with the aim of correlating the membrane structure variation in Phls and in FAs with seqA and/or dam gene mutations.
Phl composition of S. enterica serovar Typhimurium wild-type membrane
The major Phls present in S. enterica serovar Typhimurium wild-type (WT) strain membrane were PE, accounting for about 75.2%, followed by PG and CL (19.4% and 5.3%, respectively) (Fig. 2a). These Phl distributions agreed very closely to those reported in the literature for S. enterica serovar Typhimurium (Ames, 1968).
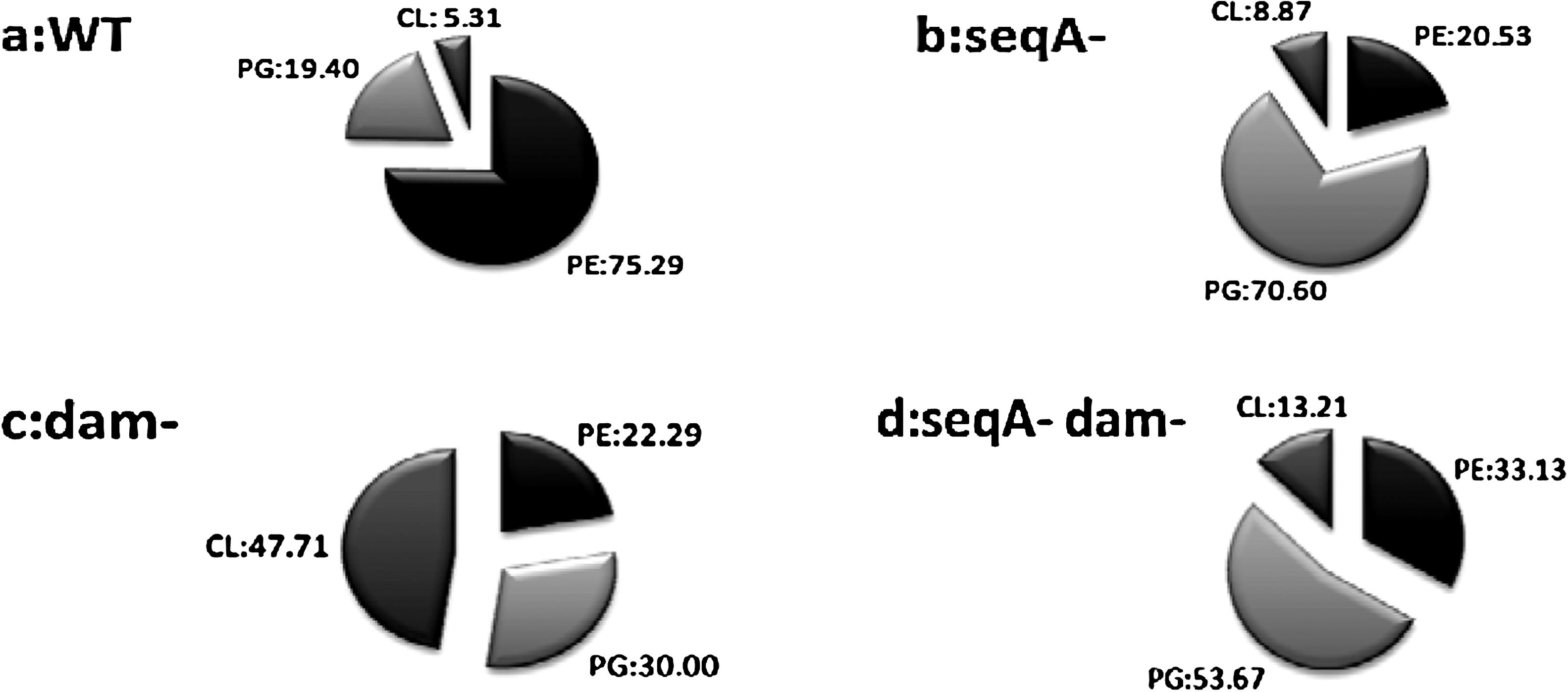
Comparative analysis of Phl levels in four isogenic strains for S. enterica serovar Typhimurium. Analysis of percentage of Phls in (
Phl composition of the seqA mutant membrane
PE, PG, and CL proportions were affected by the seqA mutation while comparing them with the WT strain. In the seqA mutant, the zwitterionic PE fraction decreased from 75.2% to 20.53%. However, the acidic Phl fraction (PG and CL) becomes a majority of total Phls with 79.47%, distributed in 70.6% of PG and 8.8% of CL (Fig. 2b).
Phl composition of the dam mutant membrane
Compared with the isogenic WT strain, the most dramatic change in Phl composition occurred with the dam mutation. In fact, the acidic Phl fractions (PG and CL) showed a great increase with 77.7% of total Phls distributed in 30% of PG and 47.7% of CL. A significant decrease in the PE fraction (22.2%) was observed (Fig. 2c).
Phl membrane composition of the seqA dam double mutant
To evaluate the combined effects of the two mutations on the bacterial membrane integrity, we compared the Phl composition of the double mutant with the isogenic WT strain. Compared with the S. enterica serovar Typhimurium WT strain, the acidic Phls (PG and CL) showed a great increase with 53.6% and 13.2%, respectively. However, PE proportion decreased from 75.2% to 33.1% (Fig. 2d).
Effect of seqA and/or dam mutations on membrane FA composition of S. enterica serovar Typhimurium
The membrane FA composition (molar percent) of the WT, SeqA−, Dam−, and SeqA− Dam− strains is shown in Table 2.
Exponentially growing Salmonella enterica serovar Typhimurium WT, seqA mutant, dam mutant, and seqA dam double mutant were incubated at 37°C, centrifuged when the optical density at 600 nm reached 0.5. Extraction of the total phospholipids (Phls) and purification of each Phl were performed as described in the experimental procedures. The contents of fatty acids (FAs) in total lipids, phosphatidylethanolamine (PE), phosphatidylglycerol (PG), and cardiolipin (CL) were determined as described in the experimental procedures. The composition of each FA was shown as the relative value to the total content of FA. Average values of triplicates were given, and the deviation was less than 5% of each value (the mean value ± the standard deviation of replicate value). Significance was assessed using the Student's t-test. FAs: tetradecanoic (myristic) acid (C14:0), hexadecanoic (palmitic) acid (C16:0), hexadecenoic (palmitoleic) acid (C16:1w7), cis-9,10-methylenehexadecanoic acid (cyc17), octadecanoic (stearic) acid (C18:0), octadecenoic (oleic or vaccenic) acid (C18:1w9), cis-9,10-methyleneoctadecanoic (dihydrosterculic or lactobacillic) acid (cyc19); minor FAs: minor fatty acids; ∑SFA: total saturated fatty acids; ∑UFA: total unsaturated fatty acids; ∑CFA: total cyclic fatty acids.
In FA designations: 0, saturated FA; 1, monounsaturated FA; cy, cyclopropane FA.
Membrane FA composition of S. enterica serovar Typhimurium WT strain
The membrane FA composition of the S. enterica serovar Typhimurium WT strain was determined (Table 2). Many FAs were found and seven main peaks were identified by comparing their retention times with those of known standards. Three saturated FAs (SFAs) were tetradecanoic (myristic) acid (C14:0), hexadecanoic (palmitic) acid (C16:0), and octadecanoic (stearic) acid (C18:0), two monounsaturated FAs (UFAs) were hexadecenoic (palmitoleic) acid (C16:1w7) and octadecenoic (oleic or vaccenic) acid (C18:1w9), and two cyclic FAs (CFAs) were the cis-9,10-methylenehexadecanoic acid (cyc17) and the cis-9,10-methyleneoctadecanoic (dihydrosterculic or lactobacillic) acid (cyc19). Their relative percentages were between 2% and 46% corresponding to more than 96% of all FAs observed. Some other minor FAs were also detected at lower relative concentrations: C17:0, C18:2w6, C18:3w6, C18:3w3, C19:0, and C20:0.
In the WT strain, C16:0, C16:1w7, and C18:0 were the main constituents, representing about 60% of total FAs (Table 2). The proportion of TL CFAs obtained was about 25.79%. However, minimum CFA levels were observed for PE and CL (6.43% and 5.08%, respectively) and higher one for PG (38.35%). The UFA/SFA ratio was in the majority with respect to between the Phl fractions (Table 2).
Membrane FA composition of S. enterica serovar Typhimurium mutant strains
To determine whether the mutations in the seqA and/or dam genes affected membrane lipid components, FA composition was quantified.
Membrane FA composition of S. enterica serovar Typhimurium seqA mutant
Our results indicated that the FA composition of the TLs appeared to be unaffected by the seqA mutation (Table 2). The loss of CL and PE was accompanied with a decrease in the proportion of C14:0, C16:0, and C16:1w7 and an increase in the proportion of C18:0 especially for the CL Phls (from 5.9% to 41.93%). Compared with the isogenic WT strain, CL and PE Phls showed an increase in the percentages of cyc19 and a decrease in their C18:1w9, which resulted in low level of acyl chain unsaturation of FAs (Table 2). The PG fraction showed a great increase of both C16:0 and cyc17 and a decrease in C16:1w7, which resulted in a low UFA/SFA ratio (Table 2).
Membrane FA composition of S. enterica serovar Typhimurium dam mutant
The FA composition of the TL was highly affected by the dam mutation (Table 2). The FAs were characterized by high level of CFA, representing about 40% of total content, and low level of UFA, representing about 2% (25% and 10%, respectively, for the WT strain). These changes were due to a concomitant decrease in C16:1w7 and C18:1w9 and an increase in their cyclopropane derivatives C17− and C19− CFA (Fig. 3) and resulted in a low UFA/SFA ratio (Table 2). The accumulation of the CL fraction was accompanied with a relative stability in the FA composition, which appeared to be unaffected by the dam mutation (Table 2). In the PG fraction, data confirmed that FA C16:0 and C18:0 are the most prominent species accounting for about 61% (43% for the WT PG fraction). Finally, we noticed that the PE Phls were characterized by a reduction in UFAs C16:1w7 and C18:1w9 to the profile of their CFA derivatives cyc17 and cyc19. These Phl changes resulted in a low UFA/SFA ratio (Table 2).
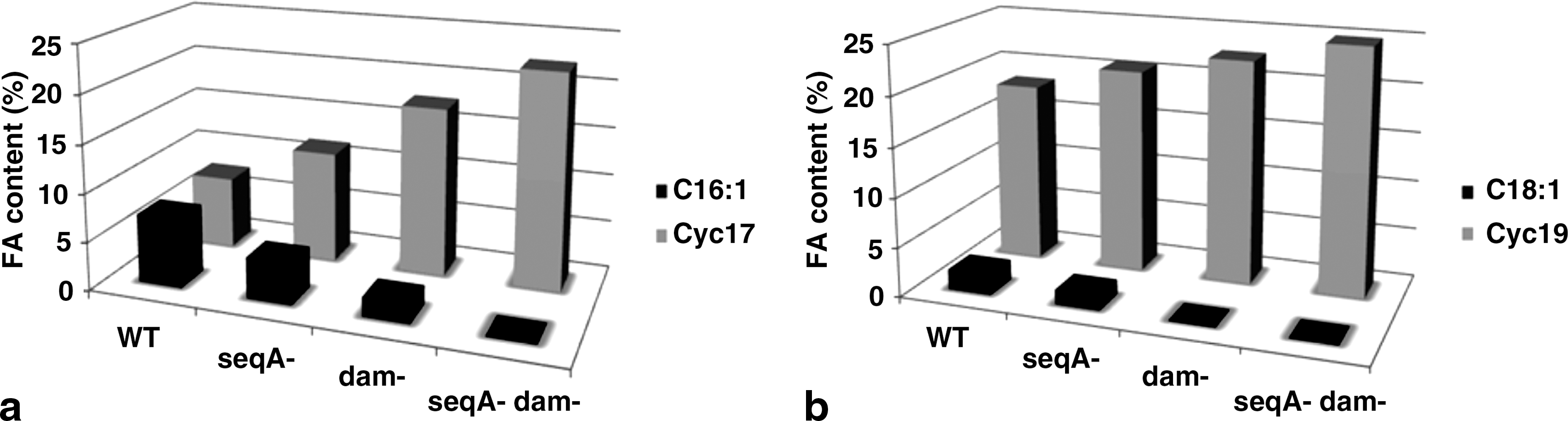
Effect of seqA and/or dam mutations on the membrane content in both C16:1w7, cyc17 (
Membrane FA composition of S. enterica serovar Typhimurium seqA dam double mutant
Our data also showed that the double mutation enhanced CFA levels (cyc17 and cyc19) from 25.79% to 47.18% (about two times higher than control cells), but decreased those of UFAs (C16:1w7 and C18:1w9) from 9.23% to 0.08% (Fig. 3). These changes resulted in very low level of acyl chain unsaturation of FAs (0.001; Table 2). For the other FAs, no significant changes were observed. In CL and PE Phls, FAs were characterized by high levels of cyc17 and cyc19 and low levels of C16:1w7 and C18:1w9. Other FA species such as myristic (C14:0) and palmitic (C16:0) were less abundant. The PG FA composition appeared to be unaffected by the seqA dam mutation with an UFA/SFA ratio, in the majority, with respect to that of WT strain (Table 2).
Discussion
Various physiological and biochemical changes took place as a consequence of many gene mutations, which can lead to numerous damages in the structure and function of the membrane cells (Taylor and Cronan, 1976; Shibuya et al., 1985; Daghfous et al., 2006). The purpose of the work presented here was to investigate a possible connection between both the seqA gene (coding for the sequestration protein SeqA) and the dam gene (coding for the Dam-methyltransferase enzyme) and some membrane components in S. enterica serovar Typhimurium. The Phls and FAs were the object of attention because the membrane delimiting the cell, and presumably conferring to it its characteristic of impermeability, is supposed to be constituted largely of lipids. Interactions of Dam and SeqA proteins with cellular membranes have been previously reported (Boye and Lobner-Olesen, 1990; Landoulsi et al., 1990; Slater et al., 1995; Wegrzyn et al., 1999; Oshima et al., 2002). However, although regulation of the activities of these proteins by membranes or their components was reported (Oshima et al., 2002) or suggested (Slater et al., 1995; Wegrzyn et al., 1999), little is known about the influence of Dam and SeqA on the composition of S. enterica serovar Typhimurium cell membranes. In this report, we have shown that zwitterionic (PE) and acidic Phl (PG and CL) proportions as well as their FAs and their acyl chain unsaturation levels were strongly affected by mutations.
Membrane Phl composition of S. enterica serovar Typhimurium: effects of seqA and dam mutations
The main classes of lipids were extracted and identified, after complete separation on a monodimensional TLC as PE, PG, and CL. The lipid extraction procedure was as described by Bligh and Dyer (1959). This method was compared with other extracting procedures reported in the literature (Folch et al., 1957; Kanfer and Kennedy, 1963; Chang and Kennedy, 1967) and was found more convenient than any of them. It is less time consuming and gives a better yield of extracted lipid. Our results agreed with the preliminary data on the major lipid classes of S. enterica serovar Typhimurium (Macfarlane, 1962; Ames, 1968) and S. newington (Dankert et al., 1966). Introduction of single or double mutations in the structural genes coding for the Dam enzyme and the SeqA protein yielded strains with Phl compositions remarkably different from the WT and from each other (Fig. 2). This depends on their genotypes, suggesting that the new phenotypes of these strains were the direct consequences of abnormalities in membrane Phl composition. As well as the increase in CL and PG synthesis, the dam mutant showed other differences in the PE content compared with the WT strain, suggesting a cross-regulation of Dam enzyme in membrane lipid pathways, crucial for the maintenance of membrane functionality and integrity. Thus, we might assume that the expression of genes coding for components of the membranes (like PE, PG, and CL synthase) or for proteins involved in synthesis of such components might be impaired in the dam mutant. This hypothesis is reinforced by the fact that Dam-mediated methylation is known to be responsible for regulating metabolism, replication, and mismatch repair (Marinus, 1996; Oshima et al., 2002; Aloui et al., 2007). As the Dam methyltransferase protein has also been proposed to form and/or maintain chromosome structure (Lobner-Olesen et al., 2003), it is likely that this protein could affect the expression of many genes, not only by direct activation or repression of particular promoters via interaction with their GATC sequences, but also by changing DNA topology. Dam and SeqA proteins participate in the movement of oriC region of the S. enterica serovar Typhimurium chromosome into the membrane and may interact with its Phls. To examine this hypothesis, we have extracted and analyzed the Phl composition of the seqA mutant membrane. Our results showed that in the absence of the SeqA protein, the bacterial membrane decreased the zwitterionic PE synthesis, which was accompanied with compensatory increase in the acidic Phls (CL and PG) content (79.47% distributed in 70.6% of PG and 8.8% of CL), and this might benefit as far as the membrane fluidity is concerned. These Phl distributions agreed very closely with those reported in the literature for E. coli (Daghfous et al., 2006) and corroborated the cross-feedback model proposed for E. coli (Shibuya, 1992). So, we suggest that in addition to its direct role in the sequestration of oriC region of the S. enterica serovar Typhimurium chromosome on the membrane, SeqA could activate or impair the expression of some genes (e.g., STM1329: putative inner membrane protein and yijP: putative integral membrane protein, respectively) (Prieto et al., 2007) that interact with lipid metabolism and regulate acidic Phl synthesis. In the seqA dam double mutant, we observed an accumulation of acidic Phls (CL and PG) compared with the S. enterica serovar Typhimurium WT strain. We can also note that PG and CL acidic Phls have intermediate proportions in relation to the seqA and dam single mutants. This suggests that Dam methyltransferase and SeqA proteins might play direct roles in Phls' metabolisms and that impairment of functions of both proteins results in less dramatic alterations in these metabolisms. All these results, led us to think that bacterial cells have developed efficient protection systems to cope with dam and/or seqA mutations. In particular, fundamental for the microbial cells is to maintain membrane integrity and functionality in response to the new phenotypes. It is well known that dam mutant initiates chromosome replication asynchronously (Boye and Lobner-Olesen, 1990; Aloui et al., 2009) and this asynchrony mainly results from an inability to inactivate newly replicated origins that have been sequestrated in the membrane (Landoulsi et al., 1990). We proposed that the initiation of the replication might be controlled by the Dam methyltransferase directly by affecting the methylation status of GATC sequences of oriC region of the S. enterica serovar Typhimurium chromosome and indirectly by regulating the DnaA activation via the modification of the acidic Phl composition. Like the Dam methyltransferase, it has been demonstrated that SeqA also affects DNA topology and inhibits open complex formation at the replication origin (Torheim and Skarstad, 1999). Indeed, Weito et al. (2000) have shown that the seqA mutation increases negative superhelicity of chromosomal and plasmid DNA and affects profoundly the transcription of various genes (Higgins et al., 1988; Wegrzyn et al., 1999; Slominska et al., 2001). In addition, SeqA is involved in translocating oriC into the membrane after initiation of DNA replication (Lu et al., 1994). How a seqA mutation can impair membrane stability is an interesting enigma. In E. coli, SeqA has been shown to associate with the E. coli outer membrane in a replication cycle-dependent manner (Slater et al., 1995). Thus, a structural role of SeqA in envelope stability cannot be discounted (Wegrzyn et al., 1999). An alternative explanation is that SeqA might regulate the expression of genes involved in the stability of the cell membranes, a possibility also considered in E. coli (Strzelczyk et al., 2003). In the seqA dam double mutant, we observed an accumulation of acyl chain UFAs in the PG fractions, which might be advantageous to cellular functions. However, Makise et al. (2002) have reported that UFAs increase membrane fluidity, which may be important for the control of the DnaA binding to oriC in vitro and can be related with the partial reestablishment of the replication asynchrony observed at every seqA and dam mutant (Lobner-Olesen and Von Freiesleben, 1996).
Membrane FA composition of S. enterica serovar Typhimurium: effects of seqA and dam mutations
The membrane FA composition of the WT, seqA −, dam −, and seqA − dam − strains is shown in Table 2. The main FAs observed for S. enterica serovar Typhimurium cells were similar to those previously described for S. enteritidis (Sampathkumar et al., 2004) and other Gram-negative bacteria such as E. coli (Casadei et al., 2002). Total SFAs, total UFAs, and total CFAs were used to determine the differences among membrane FAs of S. enterica serovar Typhimurium cells. It has been previously reported that membranes with high UFA/SFA ratio show high fluidity (Casadei et al., 2002).
The most dramatic changes in FA composition occurred in the TLs. A membrane response to seqA and/or dam mutations resulted in a relative stability in SFA levels, a slight decrease in the MFA and UFA levels, and an increase in CFA levels. This last observation resulted in a progressive decrease in the UFA/SFA ratio (Fig. 4) and membrane fluidity, going in the order from WT to the double mutant strain. Thus, the changes in the composition of the cell membrane in response to these mutations serve the purpose of maintaining a degree of fluidity, which is compatible with life. Moreover, the double mutant cells showed CFA levels about two times higher than the WT cells (Table 2). The effect of these mutations on UFA (C16:1w7, C18:1w9) and CFA (cyc17, cyc19) relative concentrations is shown in Figure 3a and b. These FAs were greatly affected by the seqA and dam mutations, which rendered a significant progressive decrease in C16:1w7 and C18:1w9 contents and a concomitant progressive increase in cyc17 and cyc19 levels probably due to the fact that a significant proportion of the UFAs (C16:1w7, C18:1w9) was converted to CFAs (cyc17, cyc19, respectively) during responses to these mutations. This conversion was done by the addition of a methylene group, derived from the methyl group of S-adenosylmethionine (SAM), across the carbon–carbon double bond of the UFA moiety of the Phls (Brown et al., 1997; Grogan and Cronan, 1997). This SAM is synthesized from methionine and adenosine tri-phosphate (ATP) by the enzyme SAM synthetase (Cantoni, 1951) and is the major methyl donor in metabolism. S. enterica serovar Typhimurium WT strain has the Dam enzyme, which catalyzes DNA methylation (Hattman et al., 1978; Herman and Mordrich, 1981; Ostendorf et al., 1999), a reaction in which SAM is the methyl group donor (Jeltsch, 2002) and also an allosteric effector (Bergerat and Guschlbauer, 1990). However, dam − and seqA − dam − strains lacking the Dam enzyme cannot use this substrate for DNA methylation, which in turn results in its accumulation and then it is used to convert UFAs to CFAs by a specific enzyme. This might explain the total transformation of dam − and seqA − dam − UFAs to their cyclopropane derivatives and the high accumulation of CFAs when compared with the WT or seqA mutant cells. These changes result in low UFA/SFA ratio, which has previously been linked to less membrane fluidity (Casadei et al., 2002; Wang et al., 2005). It is generally admitted that cells regulate their lipid composition to achieve a degree of fluidity compatible with life (Teixeira et al., 2002). Our results show that the synthesis of CFAs, which are the major components of the Phls (Grogan and Cronan, 1997) and a postsynthetic modification of the lipid bilayer that occurs as cultures of many other bacteria enter stationary phase (Chang and Cronan, 1999), plays a key role in membrane integrity against both dam and seqA mutations and many external and internal biological challenges. In addition, this synthesis of CFAs has significant implications for the survival of pathogenic bacteria. This hypothesis agreed very closely with the results of Brown et al. (1997), who showed that during acid habituation, mono-UFAs (C16:1w7, C18:1w9) present in the Phls of E. coli are either converted to their cyclopropane derivatives (cyc17 and cyc19), or replaced by SFAs. Moreover, they showed that the acid tolerance of individual strains of E. coli appeared to be correlated with membrane CFAs, and thus, they postulated that increased levels of CFAs may enhance the survival of microbial cells exposed to low pH. So, mutation in seqA and/or dam genes might result in a marked increase in transcription of some SeqA-Dam-regulated genes. It has been reported recently that the transcription of genes implicated in lipid metabolism was increased in dam mutant; Oshima et al. (2002) showed that Dam methylase protein regulates the expression of various genes implicated in lipid (accC, fabB) and phosphatidic acid (gpsA) biosynthesis. cfa and rpoS are two of these genes. These genes code for CFA synthase (Grogan and Cronan, 1984; Wang et al., 1992) and the sigma factor, which are involved in the synthesis of CFAs (Chang and Cronan, 1999). Other results that confirmed the key role of CFAs in membrane integrity was also postulated by Chang and Cronan (1999). They showed that cultures of strains lacking CFAs (as a result of a null mutation in the cfa gene) are abnormally sensitive to killing by a rapid shift from neutral pH to pH 3. This sensitivity is dependent on CFAs itself because resistance to acid shock is restored to cfa mutant strains by incorporation of CFAs from the growth medium or by introduction of a functional cfa gene on a plasmid (Chang and Cronan, 1999). However, CFA contribution to membrane properties is not yet understood, especially concerning the modifications of membrane fluidity in response to environmental stress. Duforc et al. (1984) have indicated that the presence of a cyclopropane ring within membrane FAs increases the stability of the structural and dynamic properties of biological membranes. Other studies have suggested that an increase in CFA content could cause a decrease in UFA/SFA ratio and in membrane fluidity (Yatvin et al., 1986; Magnuson et al., 1993; Annous et al., 1999), which could explain the increase in thermal resistance of S. enterica serovar Typhimurium cells with high CFA levels. On the other hand, changes observed in the membrane FA composition are not enough to explain the influence of seqA and/or dam mutations on the membrane components of S. enterica serovar Typhimurium. Therefore, other mechanisms must be responsible for these changes. Thus, these results need to be complemented by an analysis of the proteomic responses of cells to these seqA and/or dam mutations.

Effect of seqA and/or dam mutations on the membrane content in unsaturated FA (UFA) and cyclic FA (CFA) and in the ratio of acyl chain-unsaturated FAs for S. enterica serovar Typhimurium cells grown at 37°C. The ratio (mol/mol) was calculated using the data in Table 2.
Many analogies between E. coli and Salmonella mutants (including SOS induction, requirement for the SOS regulon for viability, envelope instability, and sensitivity to bile salts and other envelope-damaging agents) were observed (Prieto et al., 2007). Given these analogies, the observation that a seqA and/or dam mutations in S. enterica serovar Typhimurium does not give the same FA alteration as observed with E. coli is anecdotal. Although their genetic maps are very similar (Sanderson and Hartman, 1978), for example, the 87% amino acid identity found between the E. coli and Salmonella SeqA proteins (Prieto et al., 2007), S. enterica serovar Typhimurium and E. coli are not very related members of the family Enterobacteriaceae (Sanderson, 1976). Because of the divergence of these two species, studies on the effects of seqA and dam mutations on membrane FA composition likely yield differences. Currently, we are studying lipids and enzymatic activities from isolated membranes of S. enterica serovar Typhimurium, which can be useful to more fully understand the significance of our results and explain the differences observed with E. coli. Finally, our knowledge on the effects of SeqA and Dam proteins in S. enterica serovar Typhimurium has considerably improved in the last decade. This fundamental research has several implications that will prove to be useful for the development of novel therapeutic approaches. But, to date, therapeutic applications are still in their early experimental phases, but several recent studies provide promising results for future clinical developments. Over the last few years, many studies have demonstrated that Salmonella species dam mutants are highly attenuated for virulence in mice and have been proposed as live vaccines (Mahan et al., 2000; Low et al., 2001). Recently, it has been demonstrated that S. enterica serovar Typhimurium lacking seqA or dam gene exhibit a decrease of virulence in mice (Chatti et al., 2007, 2008; Chatti and Landoulsi 2008a), suggesting that these mutants may be used as a live vaccine. In addition, it has been reported that several bacteria with altered DNA methylation show excellent vaccine strain properties (Heithoff et al., 2001). These results prove that GATC-binding sites might have a role in regulating virulence of Salmonella and related bacteria (Chatti and Landoulsi 2008b). In conclusion, our results demonstrate that our mutant strains have different membrane composition of Phls and FA and these alterations observed in either dam and/or SeqA mutants could be a factor in the attenuation for virulence that is known for these mutants.
Footnotes
Acknowledgments
The authors are extremely grateful to Dr. Francisco Ramos-Morales (Departamento de Genética, Facultad de Biología, Universidad de Sevilla, Spain) for the generous donation of the Salmonella Typhimurium WT and disrupted strains. The authors thank also Mr. Bchini Zin for proofreading and correcting this manuscript. This work was supported by the Tunisian Ministry of Higher Education, Scientific Research, and Technology.
Disclosure Statement
No competing financial interests exist.