
Abstract
Outbreaks of Salmonella linked to fresh produce emphasize the need for rapid detection methods to curb the spread of foodborne pathogens. Reverse-transcriptase–polymerase chain reaction (RT-PCR) detects the presence of mRNA (shorter half-life than DNA), with greater potential of detecting viable pathogens. Real-time RT-PCR eliminates the need for gel electrophoresis and significantly enhances the speed of detection (<1 day) compared with traditional methods (>5 days). The objectives of this research were to apply real-time SYBR Green I-based RT-PCR to detect Salmonella from jalapeño and serrano peppers spiked with low and high inocula of Salmonella. Inoculated and uninoculated peppers were rinsed with water and dried under ultraviolet light for 10 min. Approximately 25 g peppers was inoculated with 108 to 101 colony forming units (CFU) of Salmonella enterica serovar Typhimurium in a stomacher bag and hand massaged in sterile 0.05 M glycine–0.14 M saline buffer (0.05% Tween, 3% beef extract) for optimal recovery of bacteria. A short preenrichment step of 6 h in buffered peptone water was needed for the detection of low inocula (104 CFU/25 g). One-milliliter portions of the extracts were serially diluted, plated on XLT4 agar, and used for RNA extraction with the Qiagen RNeasy® Mini Kit. RT-PCR was carried out using SYBR Green I one-step RT-PCR with previously described invA gene primers and an internal amplification control. Detection limits were 104 CFU/25 g (∼102 CFU/g) and 107 CFU/25 g (∼105 CFU/g) Salmonella from enriched and unenriched inoculated peppers, respectively. Even though this method included a 6-h incubation period, the results were still obtainable in 1 day. This method shows promise for applications in routine surveillance and during outbreaks.
Introduction
T
In recent years, the use of molecular technologies such as polymerase chain reaction (PCR) and real-time PCR (rt-PCR) have enabled the rapid detection of foodborne pathogens to enhance food safety. PCR uses gel electrophoresis to visualize the presence of the target product, whereas rt-PCR uses fluorescence. Previous studies involving rt-PCR or PCR for detection of Salmonella from produce reported detection of 1–10 colony forming units (CFU)/mL and 100 CFU/mL from mixed salad rinse waters (Bhagwat, 2004); 1 CFU/mL, 10 CFU/mL, and 100 CFU/mL from leaf lettuce and apples (Shearer et al., 2001); and 1 CFU/mL from mixed salad, cilantro, broccoli, cauliflower, and cabbage (Bhagwat, 2003). Much of these lower limits of detection were aided by a preenrichment process of around 24 h. Incubating poultry meat for 24 h allowed a detection limit of 103 CFU/mL by PCR (Kanki et al., 2009; De Medici et al., 2003). A detection limit of 103 CFU/g was demonstrated for buffalo meat trimmings using an 18-h incubation period (Biswas et al., 2008). Using rt-PCR, a detection limit of less than 5 CFU/25 g was shown for alfalfa sprouts, cantaloupe, mixed salad, and cilantro when incubated for 24 h (Liming and Bhagwat, 2004).
However, these DNA-based methods do not distinguish between live and dead cells. Reverse transcriptase–PCR (RT-PCR) is a potential method for detecting viable bacteria by targeting mRNA that has a shorter half-life than DNA (Maurer, 2006). Several researchers have investigated the suitability of using invA mRNA for the determination of Salmonella viability (Fey et al., 2004; Jacobsen and Holben, 2007; Gonzalez-Escalona et al., 2009). Real-time RT-PCR (rt-RT-PCR) could allow for the detection of recent contamination rather than cells that have already been killed because of implemented control measures to eliminate bacteria from the food product or environment. Rapid detection by rt-RT-PCR can help the producer recall contaminated fresh produce from the market at a much faster rate than traditional methods that can take ≥5 days.
The purpose of this study was to apply a previously described SYBR Green I-based one-step RT-PCR assay for the detection of Salmonella from jalapeño and serrano peppers within 1 day. Previously reported invA primers that target the chromosomally located invA gene (Galan and Curtiss, 1989) specific for Salmonella species were used in this one-step assay (D'Souza et al., 2009). Peppers were inoculated with high (108–104 CFU/mL) and low (103–101 CFU/mL) inocula and, for the latter, short enrichment times of 6 h were applied.
Materials and Methods
Growth of Salmonella enterica serovar Typhimurium
Salmonella enterica serovar Typhimurium (DT104 strain 2486) was grown in 10 mL of trypticase soy broth (Difco–Becton Dickinson, Sparks, MD) at 35°C for 24 h. The cultures were transferred for two consecutive times at 24-h intervals before use. A 10-fold serial dilution of an overnight culture (1–9 mL) in peptone buffer (Oxoid, Basingstoke Hampshire, England) per tube, was spread plated on trypticase soy agar (Difco–Accumedia, Lansing, MI) and xylose lactose tergitol 4 agar (XLT4), and incubated at 37°C for 24–48 h to determine counts.
Preparation of peppers
Chili peppers (jalapeño and serrano peppers) were purchased at a local grocery store. Before inoculation, the chili peppers were washed for 1 min using tap water, followed by rinsing with sterile deionized water, and placed in an open sterile Petri dish in a BSL-2 hood to dry under ultraviolet light for 10 min at room temperature. The chili peppers (25 g) were inoculated with 0.1 mL Salmonella enterica serovar Typhimurium high inocula (108 to 104) and low inocula (103 to 101) and allowed to dry for 10 min at room temperature under aseptic conditions in a BSL-2 hood. Washed and dried uninoculated peppers were used as controls. Each experiment was run in duplicate and replicated at least twice.
Bacterial elution from peppers
Bacteria were eluted from 25 g peppers by washing with 10 mL of 0.05 M glycine–0.14 M saline buffer (pH 9.0) (Fisher Scientific, Fair Lawn, NJ) and 0.05% Tween-20 (Promega, Madison, WI) to aid in removal of bacterial cells. To improve bacterial recovery, glycine–saline buffer with 3% beef extract and 0.05% Tween-20 was also tested and added to the chili peppers in a sterile stomacher bag. The samples were hand rubbed for 1 min and a 10 mL sample was taken and centrifuged at 8000 g for 10 min at 4°C (Sorvall Legend 23 R; Thermo Electron, Asheville, NC). The supernatant was decanted and the cell pellet was resuspended in 1 mL of 0.1% peptone buffer (Oxoid) before plating and isolation of RNA.
Preenrichment and bacterial enumeration
For low inocula levels, short-enrichment times were needed. Twenty-five grams of peppers (inoculated and uninoculated controls) was placed in 225 mL of sterile buffered peptone water (Oxoid) (Andrews and Hammack, 2007) and incubated for 6 h at 37°C. After the 6-h incubation period, a 10 mL sample was removed, placed in a 15-mL centrifuge tube, and spun at 8000 g for 10 min at 4°C (Sorvall Legend 23 R; Thermo Electron). The supernatant was decanted and the cell pellet was resuspended in 1 mL of 0.1% peptone buffer (Oxoid). Serial dilutions of enriched and unenriched samples were surface plated (0.1 mL) on XLT4 agar and incubated at 37°C for 24–48 h to enumerate typical black colonies for Salmonella detection.
Addition of stressed and autoclaved cells
Portions of 24-h grown Salmonella enterica serovar Typhimurium were also cold stressed at −20°C and 4°C and heat-stressed at ∼45°C for 24 h prior to use for testing the ability of the assay to detect stressed cells. Another portion was heated in an autoclave at 121°C, 15 pounds per square inch (psi), for 15 min. These stressed cells or autoclaved cultures were inoculated on the peppers as described earlier and either directly detected/assayed or preenriched and plated on XLT4 or assayed by real-time RT-PCR.
Isolation of RNA from bacteria
RNAlater Stabilization Reagent (Qiagen, Valencia, CA) (2 mL) was added to 1 mL of overnight Salmonella enterica serovar Typhimurium, enriched and unenriched inoculated pepper samples, uninoculated pepper controls, and peptone water. The combination of stabilization reagent and bacterial culture/extract was incubated at room temperature (25°C) for 10 min and mixed with a vortex for several times. The cells were centrifuged (5417C; Eppendorf, New York, NY) for 10 min at 8000 g at 4°C and the pellet was stored at −80°C or used directly for extraction.
The RNeasy Mini Purification Kit (Qiagen) was used to extract RNA from all samples described above and the manufacturer's instructions were followed. Briefly, 100 μL of Tris-EDTA (10 mM Tris-Cl, 1 mM ethylenediaminetetraacetic acid (EDTA), pH 8.0; Promega) containing 3 mg/mL lysozyme (Sigma–Aldrich, St. Louis, MO) was added to each pellet, mixed with a vortex, and incubated at room temperature for 10 min with mixing. RLT buffer (350 μL) was added, followed by 250 μL of 100% ethanol (Aaper Alcohol, Shelbyville, KY), and transferred to the RNeasy mini column. The column was centrifuged at 8000 g for 15 s and the collection tube was discarded. RW1 buffer (700 μL) was added to the column and centrifuged for 15 s at 8000 g to wash the column. RPE buffer (500 μL) was added and centrifuged at 8000 g for 15 s and repeated once. The RNA was then eluted off the column with 40 μL of RNase-free water and centrifuged at 8000 g for 1 min. The collection tubes containing RNA were capped and stored at −80°C until further analyses.
DNase I treatment
DNase I treatment was carried out using the TURBO DNA-free® Kit (Ambion®; Applied Biosystems, Austin, TX). The procedure consisted of 5 μL of 10 × TURBO DNase buffer, 1 μL of rDNase I DNA-free, and 34 μL of nuclease-free water. The reagents were then mixed with 10 μL of RNA extracts and incubated at 37°C for 30 min. Five microliters of DNase Inactivation Reagent was added and incubated for 2 min. The sample was centrifuged at 10,000 g for 2 min. The supernatant was removed and placed in a new tube.
Nanodrop quantification of RNA
After extraction of RNA (before and after DNAse I treatment), RNA quantification was carried out using a NanoDrop® Spectrophotometer (NanoDrop ND-1000 Spectrophotometer; Thermo Scientific, Wilmington, DE). Nucleic acid concentration was determined using ultraviolet spectrophotometry at 260 nm and purity was determined using the absorbance ratios of 260/280 and 260/230 to detect protein and organic carryover, respectively. Samples with both ratios ≥1.8 were used to provide a good quality of RNA with little to no carryover.
rt-RT-PCR procedure
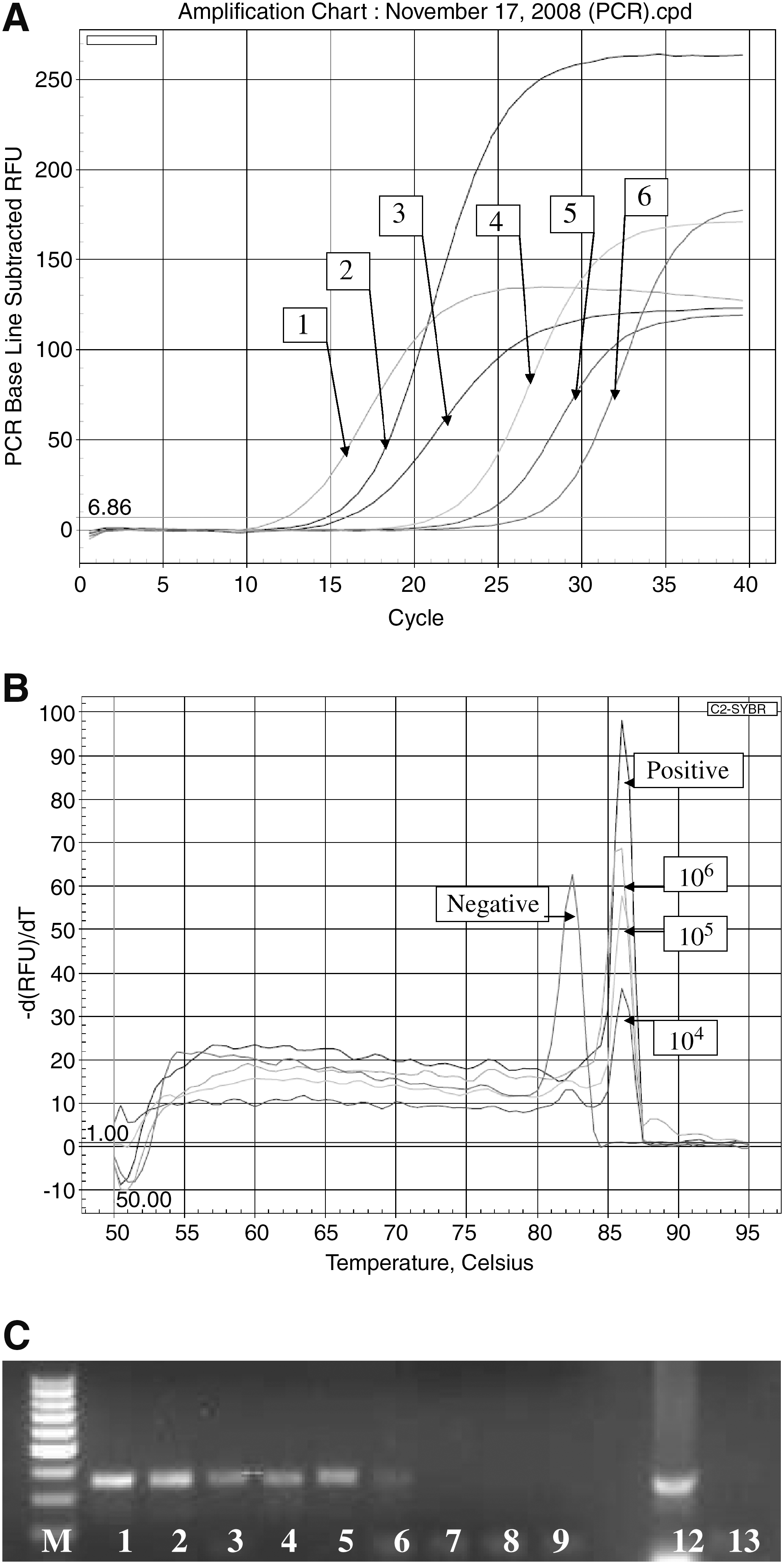
The SuperScriptTM III Platinum® SYBR® Green One-Step qRT-PCR Kit (Invitrogen, Carlsbad, CA) was used in the amplification of mRNA to cDNA, followed by DNA amplification as described by D'Souza et al. (2009). Briefly, 25 μL reactions contained 5 μL RNA extracts in RNAse–DNAse free water, Superscript III (SSIII) one-step RT-PCR kit reagents (Invitrogen) with SYBR Green I, 0.02 μM of each invA primer (F: 5′CACGCTCTTTCGTCTGGCA3′; R2: 5′TACGGTTCCTTTGACGGTGCGA3′), and 1.6 μL bovine serum albumin (Sigma–GenoSys, St. Louis, MO) and an internal amplification control (IAC) (see below). The thermocycler conditions were as follows: RT at 50°C/30 min, denaturation at 95°C/5 min, followed by 45 cycles at 95°C/45 s, 58°C/45 s, 72°C/45 s, and final extension at 72°C/7 min in an iCycler (Bio-Rad, Hercules, CA). After amplification, melt temperature (T m) analysis from 50°C to 95°C with 0.5°C increments was carried out. The software provided by iCycler was used to obtain threshold cycle values, T m, and the standard curve. The expected T m was ∼86°C for the Salmonella-specific invA product and 82°C for the IAC product.
Agarose gel electrophoresis
Products were separated by agarose gel electrophoresis (2% agarose gels) (in 1 × Tris-acetate–EDTA buffer; Fisher Scientific, Pittsburgh, PA) and visualized under ultraviolet light after staining with ethidium bromide using the Gel Doc System/Station (Bio-Rad). A 100-bp DNA marker (Promega) was also run to confirm the presence of the 347-bp target amplified product and the 154-bp IAC product.
Preparation of the IAC
The IAC was added to the SuperScript III Platinum SYBR Green One-Step qRT-PCR mix to eliminate the possibility of false-negatives due to PCR inhibitors, enzyme inactivation, or instrument malfunction as described earlier (D'Souza et al., 2009). The concentration of the amplified IAC RNA product was diluted to reach an optimal concentration of 1.9 fg/μL in the rt-RT-PCR mix.
Results
Bacterial cell recovery from peppers
Extracting Salmonella from the pepper skin proved to be much easier than removal from the stem of the pepper. The hard, smooth surface of the pepper without any crevices left no room for trapping target bacteria (Rusin et al., 2002). Recovery of Salmonella enterica serovar Typhimurium cells, as determined by plating from the surface of the jalapeño and serrano peppers, was about 103 CFU from the initial inocula of 107 CFU/25 g using the optimal glycine–saline buffer containing Tween-20 and beef extract (Table 1). The addition of Tween-20 acted as a surfactant to help lift the cells off the outer surface of peppers and into suspension. When a lower inoculum of 104 CFU/25 g was used, recovery remained at ∼1.08 × 103 CFU/25 g after enrichment as determined by plating on XLT4 agar and observance of typical black colonies when compared with Salmonella controls.
RNA extraction yields from peppers
For detection of Salmonella without enrichment, the 107 CFU inoculum yielded an average of 6 ng/μL nucleic acid from both types of peppers. Purity of the RNA extracts was assessed, which showed little to no protein or organic carryover using the ratios of A260/A280 and A260/A230 that were >1.8. After an enrichment process, with an improved 104 detection limit, 263.34 ± 18.39 ng/μL of RNA was extracted with little or no protein or organic carryover.
rt-RT-PCR detection and specificity
Detection of 107 CFU/25 g from both inoculated jalapeño and serrano peppers without enrichment was achieved using RT-PCR (Fig. 1A). After 6-h preenrichment, an increase in detection by 3 logs to 104 CFU/25 g (Fig. 2A) for both jalapeño and serrano peppers was obtained. Even with preenrichment, this method could still yield results in 1 day. To determine the specificity of the amplified products, T m analysis was carried out. T m curves showed peaks at ∼86°C for the target-amplified invA product and the IAC showed peaks at 82°C. This showed that false-positives were not obtained and that indeed the detection limit was 104 CFU/25 g (Figs. 1B and 2B). Also, the possibility of false-negatives was eliminated, because the IAC peak at 82°C showed that no inhibitors were present and that there was no equipment malfunction or enzyme failure. Further confirmation by gel electrophoresis revealed the 347-bp target product as expected, with similar detection limit to RT-PCR Ct values of 104 CFU/25 g. Gel electrophoresis is generally not required during routine analysis using rt-PCR, but was carried out during this research to confirm the presence of the target products of appropriate size. Negative controls did not show any T m at ∼86°C or any product at 347 bp as expected. Negative controls consisted of water, trypticase soy broth, peptone, or uninoculated peppers, which did show the T m at 82°C for the IAC and the 154-bp IAC product by agarose gel electrophoresis (Figs. 1C and 2C). Again, this showed that false-negatives were not obtained.

(
(
Detection of stressed cells
Cold-stressed Salmonella enterica serovar Typhimurium cells at 4°C and −20°C as well as heat-stressed cells at 45°C overnight (at high and low inoculum levels) from both peppers showed similar detection to nonstressed cells using high and low inocula levels. Preenrichment for 6 h aided in the recovery and detection of the low inoculums of stressed cells from peppers in a manner similar to nonstressed cells (data not shown). Peppers inoculated with autoclaved cells did not result in detection by this real-time RT-PCR method.
DNase I treatment
DNase I treatment was performed to determine that amplification was from RNA without any carryover of DNA. When traditional PCR on the DNase I-treated extracts using the described invA gene primers was carried out, all treated samples did not result in any positive PCR products as expected, demonstrating the absence of any DNA carryover. However, the ability to detect the target RNA from jalapeño and serrano peppers was reduced by 1 log in unenriched peppers and also by 1 log on enriched peppers (data not shown).
Discussion
Salmonella from jalapeño and serrano peppers were recovered by gently massaging the outer surface of the pepper in an optimal glycine–saline buffer solution containing 3% beef extract and 0.05% Tween-20. Previous research suggested that the attachment of bacterial cells to the surface of produce could be due to the roughness or hydrophobic properties of the produce surface (Iturriaga et al., 2003). This study used a buffer solution that contained a surfactant to aid in removal of the Salmonella cells from the surface of peppers to improve recoveries. The similar amounts of RNA recovered by all tested buffers could be attributed to the lower end detection limit of the instrument, even though increased recovery in bacterial count was obtained. Further optimization of elution buffers is needed to improve recovery of bacteria from produce surfaces. Several factors may have played a role in the low recovery, as attachment of bacteria to produce surfaces depends on the serovar of Salmonella (Shi et al., 2007), as some microorganisms bind or attach more strongly than others. In addition, depending on the drying time on the surface of produce, cell injury or death may have occurred (Lang et al., 2004). Preenrichment or enrichment steps can help aid in the recovery/resuscitation of these injured cells as well as increase cell numbers to improve detection probability. Therefore, this study used short preenrichment times of 6 h to overcome these limitations of recovery and improve detection of target bacteria to obtain rapid and sensitive detection within two 8-h working shifts.
Several studies have shown that an 18–24-h enrichment procedure is necessary to determine the presence of low numbers of bacteria (Perelle et al., 2004; Cheng et al., 2008). An 18-h enrichment process was needed to obtain a detection limit of 103 CFU/mL by PCR for Salmonella in raw milk (Perelle et al., 2004). Cheng et al. (2008) showed that a 24-h enrichment process prior to RT-PCR was needed to obtain a detection limit of 103 CFU/mL for Salmonella in chili powder. Bhagwat (2004) showed a lower Salmonella detection limit of 100 CFU/mL from mixed salad rinse after enrichment for 5 h. Others showed detection of 1 CFU/mL from leaf lettuce and apples (Shearer et al., 2001) for Salmonella after a 24-h enrichment; and 1 CFU/mL from mixed salad, cilantro, broccoli, cauliflower, and cabbage for Salmonella enterica serovar Typhimurium using rt-PCR (Bhagwat, 2003), after a 20-h enrichment. In this study, a shorter 6-h enrichment process allowed the detection of lower inoculum levels at ∼104 CFU/25 g within two 8-h working shifts.
Although the major advantage to rt-PCR is the speed of detection (Feng, 2007), both PCR and rt-PCR target DNA that does not distinguish between dead and viable cells. An advantage of using RT-PCR is that it targets mRNA with a shorter half life than DNA, and therefore, has greater potential for detecting primarily viable cells that could cause infection (Maurer, 2006). As most quality control analyses are done on finished products, this is an excellent way to determine if the critical control points are working and that no viable cells are present.
Studies have been carried out to determine the suitability of using RNA to detect the viability of cells. Researchers have shown that invA mRNA from Salmonella enterica serovar Typhimurium can degrade rapidly after 50 h in drinking and pond water (Fey et al., 2004). These researchers quantified S. Typhimurium mRNA in drinking water using invA and 16s RNA genes that detected 2–20 and 100–1000 copies, respectively. Real-time RT-PCR with invA gene primers showed a detection limit of 5 × 104 seeded S. Typhimurium cells per gram of soil (Jacobsen and Holben, 2007), and pasteurizing the samples at 65°C for 30 min resulted in the inability to detect invA mRNA. These results indicate that invA mRNA appears to be a suitable target for cell viability determination for Salmonella.
Recently, using the same target invA mRNA, a TaqMan-based RT-PCR method has shown a detection limit of 5 CFU/25 g for Salmonella Saintpaul and Salmonella Newport from spinach, tomatoes, and jalapeno and serrano peppers; however, a longer preenrichment of 24 ± 2 h was used (Gonzalez-Escalona et al., 2009). Even though our study is similar as it is based on the invA mRNA for RT-PCR detection, it differs from the work described by these researchers in that the less-expensive SYBR Green I fluorescent dye was used in the real-time assay when compared with TaqMan® fluorescent probes. Also, our study successfully used a shorter preenrichment time of 6 h for Salmonella enterica serovar Typhimurium detection from jalapeno and serrano peppers.
An IAC was applied to determine if there were any false-negatives that may have been caused by PCR inhibitors (Moreira et al., 2009), enzyme failure, or instrument malfunction. PCR inhibitors from food matrices include lipids, proteins, enzymes, chemical additives, fiber, other bacteria, and acids (Lee and Fairchild, 2006). The target and IAC were both amplified using the same primer sets to enable the use of only one set of PCR reaction conditions as has been suggested by other researchers (Hoorfar et al., 2004).
A DNAse I treatment was performed to eliminate genomic DNA that may have been present (Werbrouck et al., 2007). Harper et al. (2003) suggested that all genomic DNA may not be completely removed by the treatment process. In this study, traditional PCR run on DNAse I-treated samples did not show any product on agarose gels, indicating that DNA was indeed eliminated (data not shown). Importance must be placed on removal of any carryover DNA that could lead to erroneous results. The lower yield of RT-PCR after DNAse I treatment showed that caution must be used while extracting RNA. It is also crucial to ensure that the DNAse I does not contain any RNAses that could destroy or degrade the total RNA. The RNAses, if any present, could result in decreased yield (Werbrouck et al., 2007). When DNAse I was applied to our study, a 1 log reduction in detection was observed. It is possible that the loss of 1 log could be due to the DNAse I containing some carryover RNAses. This could have resulted in the degradation of some of the RNA present, leading to lower yields.
In nonhost environments, Salmonella more than likely persists in starved or stressed states (Gonzalez-Escalona et al., 2009); therefore, stressed cells were used to inoculate the peppers for detection by this newly developed SYBR Green I-based assay. Salmonella enterica serovar Typhimurium cells that were cold-stressed at 4°C and − 20°C as well as heat-stressed at ∼45°C overnight (at high and low inoculum levels) were used to inoculate peppers and also to simulate conditions during transport or storage of peppers. Preenrichment for 6 h aided in the recovery and detection of these stressed cells, yielding similar detection limits compared with nonstressed cells. Also, previous studies using inoculation of a mixture of microorganisms (Escherichia coli, Enterococcus faecalis, Lactobacillus, etc.) associated with background flora on food commodities followed by enrichment did not show any cross-reactivity by this rt-RT-PCR assay (Techathuvanan et al., 2009a). This assay also did not show cross-reactivity against a panel of various non-Salmonella bacteria in pure culture as previously reported (D'Souza et al., 2009). Amplified products were not obtained/detected either by T m analysis or gel electrophoresis from the extracts of the background flora (either pure culture or when inoculated on food matrices), indicating the specificity of this assay.
Detection limits of around 400 CFU/g (104 CFU/25 g) after a 6-h enrichment step using RT-PCR was similar to results obtained by other researchers using molecular detection methods for produce. Yuk et al. (2006) evaluated Salmonella on the surface of tomatoes with a detection limit of 102 CFU/tomato, with an enrichment period of 4 h. Even though Yuk et al. (2006) used only a 4-h enrichment process, the additional cost of using the flow-through immunocapture system may be a limiting factor in using this method. This newly developed rt-RT-PCR assay requires approximately similar time for detection when compared with nucleic acid sequence-based amplification assays (D'Souza and Jaykus, 2003) or novel loop-mediated isothermal assays (Techathuvanan et al., 2009b) to obtain similar results. However, as suggested by Gonzalez-Escalona et al. (2009), collaborative studies to determine interlaboratory reproducibility are needed before the method can be used for routine diagnostic purposes, without excluding the need for the physical isolation of bacteria for regulatory purposes.
In conclusion, this method rapidly detected Salmonella within two 8-h workshifts and was much faster than traditional plating. Rapid Salmonella detection methods will ultimately save the food industry time and money by avoiding costly recalls and protecting public health. Future work should focus on improving bacterial recovery, RNA yields, and purity to increase detection sensitivity to ≤1 log CFU/g of peppers within 1 work day or at the most, two workshifts, to allow for timely product recall or prevention of release of contaminated products for sale.
Footnotes
Acknowledgments
The authors gratefully acknowledge the support of the Tennessee Agricultural Experiment Station for this research. This research was carried out by Nathan Miller as part of the requirements for the fulfillment of his Master's Degree. The authors are grateful to Dr. David A. Golden for critically reviewing and editing this manuscript. The use of trade names in this manuscript does not imply endorsement by the University of Tennessee nor criticism of similar ones not mentioned.
Disclosure Statement
No competing financial interests exist.