
Abstract
Background:
The study sets out to either confirm or refute a recent study's findings that chicken meat is an unlikely source of Salmonella enterica subspecies enterica serovar Infantis (Salmonella Infantis) in humans in the Kyushu-Okinawa region, Japan.
Methods:
A total of 74 Salmonella Infantis isolates (30 from human and 44 from other sources), mainly from the Kyushu-Okinawa region in south-western Japan, were analyzed using a molecular-epidemiological approach combining two fingerprinting methods, namely pulsed-field gel electrophoresis and amplified fragment length polymorphism (AFLP), a novel polymerase chain reaction-based technique.
Results:
The resulting pulsed-field profiles showed that 17 of 30 human isolates were similar to those found in chicken meat, whereas there were no common pulsed-field profiles between human and chicken egg isolates. Overall, 3 of 18 AFLP profiles included 7 human isolates and 14 chicken egg isolates. In addition, the combined results of the pulsed-field gel electrophoresis and AFLP analyses showed that 8 human Salmonella Infantis and 13 chicken meat isolates belonged to the same clonal lines.
Conclusions:
These results suggest that chicken meat is an infection source of Salmonella Infantis for humans in the Kyushu-Okinawa region, Japan. The results also showed the relatively high suitability of AFLP for application to epidemiological studies of Salmonella Infantis.
Introduction
For the purpose of food hygiene, it is important to solve the question whether chicken meat represents an infection source of Salmonella Infantis for humans in the Kyushu-Okinawa region, Japan. Kudaka et al. (2006) have reported that the infection source of human salmonellosis with Salmonella Infantis is unlikely to be chicken meat in Okinawa (a southern part of the Kyushu-Okinawa region), Japan. They characterized 108 isolates of Salmonella Infantis including 15 human (taken from two hospitals), 1 layer hen (collected from a farm), 60 chicken meat parts (collected from 15 grocery stores and 2 factories), and 32 broiler isolates (collected from four farms). They classified these isolates into 25 genotypes with the molecular-epidemiological technique of pulsed-field gel electrophoresis (PFGE) using two kinds of enzymes. Subsequently, they reported that human isolates (showing 6 different types) never coincided with other source isolates (showing 19 different types). Their interpretation of their own data suggests that there is no hazard of Salmonella in chicken meat. Their study strongly affects the education of the food handlers and consumers in relation to food hygiene in the Kyushu-Okinawa region, Japan. However, our previous findings of high contamination rates of the pathogen (25.6%) in chicken meat and low contamination rates in other foods (Murakami et al., 2001) suggested that chicken meat was the most likely infection source of Salmonella Infantis. Other studies also suggested that chicken meat is one of the infection sources of Salmonella Infantis (Wilkins et al., 2002; Nógrády et al., 2007). Investigations of chicken meat as infection sources are important for preventing salmonellosis and reducing the prevalence of asymptomatic Salmonella Infantis carriers. The purpose of this study was to evaluate whether chicken meat is actually an infection source of Salmonella Infantis for humans in the Kyushu-Okinawa region, Japan.
Because the combined usage of different methods has been deemed more effective than single-technique usage (Arbeit, 1995), PFGE and amplified fragment length polymorphism (AFLP) were chosen for this study. PFGE is the most widely used genetic fingerprinting method for Salmonella (Murakami et al., 1999a; Janda and Abbott, 2006). AFLP is a novel polymerase chain reaction (PCR)-based technique that has been used for DNA fingerprinting of plant genomes (Savelkoul et al., 1999) and bacteria (Huys et al., 1996). Several AFLP-based studies on Salmonella have indicated that AFLP is a highly suitable technique for epidemiological studies of Salmonella (Lindstedt et al., 2000; Gebreyes and Altier, 2002; Hu et al., 2002; Torpdahl et al., 2005).
Materials and Methods
Bacterial isolates
The 74 isolates of Salmonella Infantis used in this study were all from the collection of Fukuoka Institute of Health and Environmental Sciences (FIHES) between 1996 and 2005 (Table 1). The isolates were from a foodborne disease outbreak sample, food and farm inspection samples, samples from periodic inspections of food handlers, and samples of various sporadic cases (symptomatic patients), with the exception of the swine and cattle isolates. The foodborne disease outbreak isolate (a symptomatic patient's isolates) and the food isolates (25 chicken meat and 13 chicken egg isolates) were isolated from samples in FIHES; the farm-related isolates (three layer hens, a swine, and a cattle isolate) were provided by livestock hygiene service centers; the food handler isolates and nine symptomatic patient isolates were isolated in a clinical laboratory and serotyped in FIHES (Murakami et al., 2007). These isolates were collected from 13 different prefectures in Japan. Although the majority of the samples were obtained from the Kyushu-Okinawa region (which consists of eight prefectures including Japan's third largest island and is located southwest of the main island of Honshu), six isolates (except for a control strain) were obtained from regions outside of Kyushu-Okinawa. The majority of the isolates (95.9%) were obtained between 1996 and 2001. Salmonella Infantis American Type Culture Collection 51741 was included as the control.
AFLP
AFLP analysis was performed as described in a previous report (Noda et al., 2006) as well as in other reports (Keim et al., 1997; Desai et al., 1998), with slight modifications. This method consisted of seven processes, namely DNA extraction, DNA digestion, DNA ligation, preselective PCR, selective PCR, electrophoresis, and fragment interpretation. In the DNA extraction process, genomic DNA (approximately 100 ng) of each isolate was extracted using a commercial kit (ISOPLANT Kit; Nippon Gene, Tokyo, Japan) and the concentration of DNA was estimated with a spectrophotometer (PerkinElmer, Waltham, MA). In the digestion and ligation processes, the genomic DNA from each isolate was digested with the enzymes MseI (Daiichi Pure Chemicals, Tokyo, Japan) and EcoRI (Daiichi Pure Chemicals) and the fragments were ligated to MseI and EcoRI restriction half-site-specific double-stranded adaptors (Applied Biosystems, Carlsbad, CA) in a total volume of 11 μL. The reaction mixture contained 1 U of MseI, 5 U EcoRI, 1.1 μL of 10 × T4 DNA Ligase Buffer with ATP (New England Biolabs, Ipswich, MA), 1.1 μL of 0.5 M NaCl, 10 U of T4 DNA Ligase (New England Biolabs), 0.5 μL of 1 mg/mL bovine serum albumin, 1 μL of MseI adaptor, and 1 μL of EcoRI adaptor. The adaptors were heated at 95°C for 5 minutes before use. The mixture was incubated at 23°C for 8 hours and heated at 65°C for 20 minutes in a PCR Thermal Cycler SP (Takara Shuzo, Kyoto, Japan), before being diluted to 200 μL with 0.1 × TE buffer (10 mM Tris-HCl [pH 8.0], 0.1 mM ethylenediaminetetraacetic acid). The preselective PCR process was carried out in a final volume of 20 μL comprising 4 μL of the diluted restriction–ligation mixture, 0.5 μL of MseI preselective primer (Core Sequence; Applied Biosystems), 0.5 μL of EcoRI preselective primer (Core Sequence; Applied Biosystems), and 15 μL of AFLP Amplification Core Mix (Applied Biosystems). The preselective PCR cycling conditions used for amplification are as follows: initial reaction for 2 minutes at 72°C, followed by 20 cycles of denaturation at 94°C for 20 seconds, annealing at 56°C for 30 seconds, and extension at 72°C for 2 minutes. These cycling steps were carried out with 0.01 ramp times. Preselective PCR was performed using a PE-9600 thermocycler (PerkinElmer). After amplification, the PCR sample was diluted by 10-fold with 0.1 × TE buffer. The selective PCR process was performed in a final volume of 20 μL comprising 1.5 μL of the diluted amplification PCR sample, 0.5 μL of MseI + G selective primer (Applied Biosystems), and 0.5 μL of EcoRI + 0 selective primer (Applied Biosystems). The selective PCR cycling conditions used for amplification are as follows: initial denaturation for 2 minutes at 94°C (1 cycle), followed by 30 cycles of denaturation at 94°C for 20 seconds, annealing for 30 seconds at various temperatures (see below), and extension at 72°C for 2 minutes, and a final extension at 60°C for 30 minutes. The annealing temperature was 66°C, also for the first cycle, and decreased by 1°C for each of the following nine cycles. The annealing temperature for the remaining 20 cycles was 56°C. Selective PCR was performed using the PE-9600 thermocycler. In this study, combinations of MseI selective and EcoRI selective primers were selected from an AFLP microbial fingerprinting kit with 64 possible combinations (Applied Biosystems). In our preliminary experiments, we tried eight combinations of selective primers using some of our isolates of Salmonella Infantis. The eight MseI–EcoRI selective primer combinations evaluated were 0 (nonselective)-A, 0-C, 0-G, 0-T, A-0, C-0, G-0, and T-0. Subsequently, we chose the combination of G-0 (MseI + G − EcoRI + 0) as our selective primers because their performance with respect to the peak height of the fragments and the number of fragments was superior to the other seven combinations. In the electrophoresis process, the amplified products were separated in a 5% denaturing (sequencing) polyacrylamide gel (Applied Biosystems) in an ABI Prism 377 DNA automated sequencer (PerkinElmer). An aliquot (2 μL) of the reaction mixture was added to 2.5 μL of loading dye (a mixture of 1.7 μL of formamide [Amresco, Solon, OH], 0.3 μL of loading solution [Applied Biosystems], and 0.5 μL of internal size markers [Applied Biosystems]). To accurately evaluate the sizes of the EcoRI/MseI fragments, GeneScan-500 markers labeled with the red fluorescent dye 6-carboxy-χ-rhodamine were used as internal size standards. The sample mixture was heated at 95°C for 3 minutes, cooled on ice, and immediately loaded onto the gel. The running buffer was 1 × TBE buffer (89 mM Tris, 89 mM boric acid, 2 mM ethylenediaminetetraacetic acid), and the electrophoresis conditions were 3.0 kV at 51°C for 3.5 hours. The well-to-read distance was 36 cm. In the fragment interpretation process, fragments were sized with GeneScan software version 2.1 (Applied Biosystems). The peak height threshold was set at 50, and any peak heights below this value were excluded from the analysis. Electropherograms of all fragment profiles were visually inspected for polymorphisms. The presence and absence of fragments were scored in a binary matrix and recorded as a text (tab-delimited) file in Excel version 2003 (Microsoft, Redmond, WA). The text file was transferred to Fingerprinting II Japanese Edition software (Bio-Rad Laboratories, Hercules, CA). Similarity and cluster analyses were performed with the Dice coefficients of similarity and an unweighted pair group method using average linkage (UPGMA), respectively. As the reproducibility of AFLP was 85.6% in our preliminary experiments for reproducibility using three isolates, we typed the isolates at a similarity linkage level of 86% as the cutoff value in this study. Simpson's index of diversity (Hunter and Gaston, 1988) was used to express the discriminatory power of AFLP.
PFGE
PFGE was conducted as described in our previous report (Murakami et al., 1999b). The DNA of Salmonella Infantis was digested with 20 U BlnI (Takara Bio, Otsu, Japan) at 37°C for 15 hours. PFGE was performed using a CHEF-Mapper apparatus (Bio-Rad Laboratories) for 24 hours at 200 V and 14°C, with a pulse time ranging from 2 to 43.2 seconds. The DNA fragment patterns were assessed visually. Isolates were considered to be genetically similar or identical if there was complete concordance of the DNA fragment profiles and were considered to be different if there was a difference of one or more of DNA bands in the DNA fragment profiles. Similarity and cluster analyses were performed with the Dice coefficients of similarity and UPGMA, respectively, using Fingerprinting II Japanese Edition software (Bio-Rad Laboratories). Simpson's index of diversity (Hunter and Gaston, 1988) was also used to express the discriminatory power of PFGE.
Results
The results of AFLP
AFLP analysis of the 74 Salmonella Infantis isolates produced approximately 44 fragments per isolate, ranging in size from 35 to 495 bp. The obtained fingerprints and a UPGMA dendrogram of the isolates are shown in Figure 1. The isolates were typed into 18 AFLP profiles at a similarity linkage level of 86% as the cutoff value (Table 2, Fig. 1). The value was based on the reproducibility at 86%. The Simpson's index of AFLP typing results was 0.90. The dominant AFLP profiles were profiles 5, 9, and 11, encompassing 16, 8, and 11 isolates, respectively.
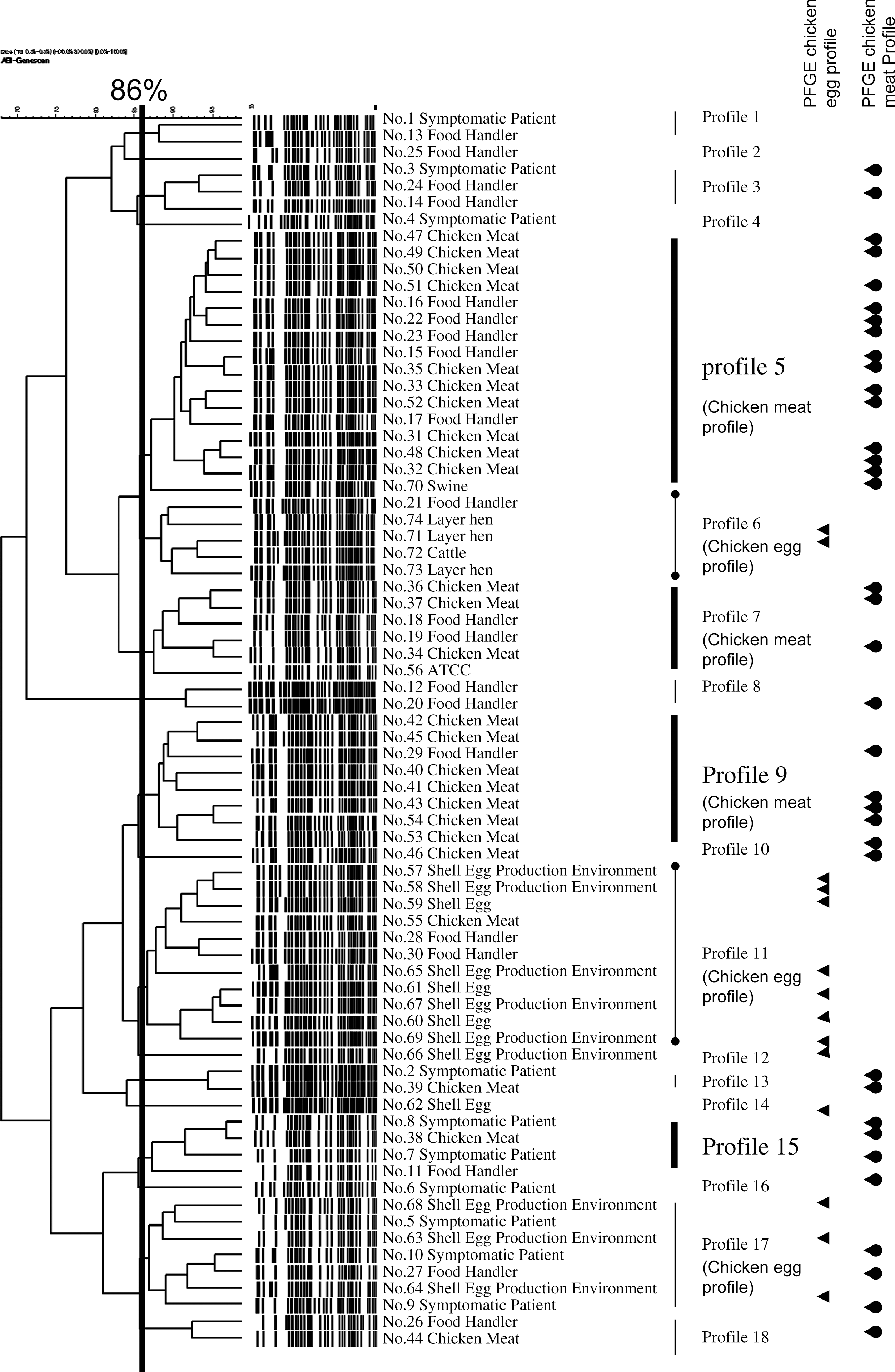
Amplified fragment length polymorphism analysis of fingerprints from genetically defined Salmonella enterica subspecies enterica serovar Infantis (Salmonella Infantis) isolates. The numbers on the right of the dendrogram are isolate numbers (Table 2). The scale indicates the percentages of similarity, as determined by the Dice coefficients. Among the 18 profiles, 7 profiles (5, 7, 9, 11, 13, 15, and 18) include 15 human isolates and 24 chicken meat isolates. Arrows without circles indicate isolates belonging to the “pulsed-field gel electrophoresis (PFGE) chicken egg profiles” by PFGE. Arrows with circles indicate isolates belonging to the “PFGE chicken meat profiles” by PFGE.
Origins were divided into six categories: food handler, symptomatic patient, chicken meat, chicken egg, swine, and cattle (except for ATCC 51741).
Clonal line including isolates from both humans and chicken meat.
Out of Kyushu-Okinawa region isolates.
Chicken egg was included in SEPE, shell-egg at shell-egg processing facility, and layer hen.
SEPE, shell-egg production environment; U, H, and S represent company names.
The chicken egg isolates belonged to profiles 6, 11, 12, 14, and 17, whereas the chicken meat isolates belonged to profiles 5, 7, 9, 10, 11, 13, 15, and 18. Among the 18 AFLP profiles, each of 7 profiles (5, 7, 9, 11, 13, 15, and 18) included human isolate(s) and isolate(s) from chicken meat simultaneously (a total of 15 human and 24 chicken meat isolates), whereas each of 3 profiles (6, 11, and 17) included 7 human and chicken egg isolates simultaneously (a total of 7 human and 14 chicken meat isolates). The swine isolate was in profile 5 with human and chicken meat isolates. The cattle isolate belonged to profile 6 with human and chicken egg isolates.
Some profiles containing the isolates from either chicken egg or chicken meat were defined as specified profiles for further discussion. Profiles 6, 11, and 17 included two or more chicken egg isolates and were defined as “AFLP chicken egg profiles.” A total of 23 isolates belonged to these profiles (Fig. 1). Profiles 5, 7, and 9 included two or more chicken meat isolates and were defined as “AFLP chicken meat profiles.” A total of 30 isolates belonged to these profiles (Fig. 1).
The results of PFGE
The 74 isolates generated 24 different pulsed-field profiles (PFPs) after restriction with BlnI (Fig. 2). (Some PFPs of Salmonella Infantis, PFPs 1, 3, 5–8, 11–14, 16, 19, 21–24, 26–35, 47, 49, and 50, were assigned numbers in a previous study and are thus not consecutively numbered in this study.) The Simpson's index of PFGE typing results was 0.86. The PFPs of the chicken meat isolates were clearly different from those of the chicken egg isolates. The PFPs of the chicken egg isolates showed no bands below approximately 90 kbp, except for PFP 20, whereas the PFPs of the chicken meat isolates revealed some bands in this size range (Fig. 3). Only two PFPs (2 and 4) included isolates from both humans and other sources simultaneously, and these PFPs only included isolates from chicken meat and humans (Figs. 2 and 3). All the human isolate PFPs were clearly different from the chicken egg isolate PFPs (Table 2, Figs. 2 and 3). In contrast, the PFPs of 17 human isolates were indistinguishable from the PFPs of chicken meat isolates. The swine isolate was PFP 4 and therefore belonged to “PFGE chicken meat profiles.” The cattle isolate belonged to a distinct profile, PFP 45.
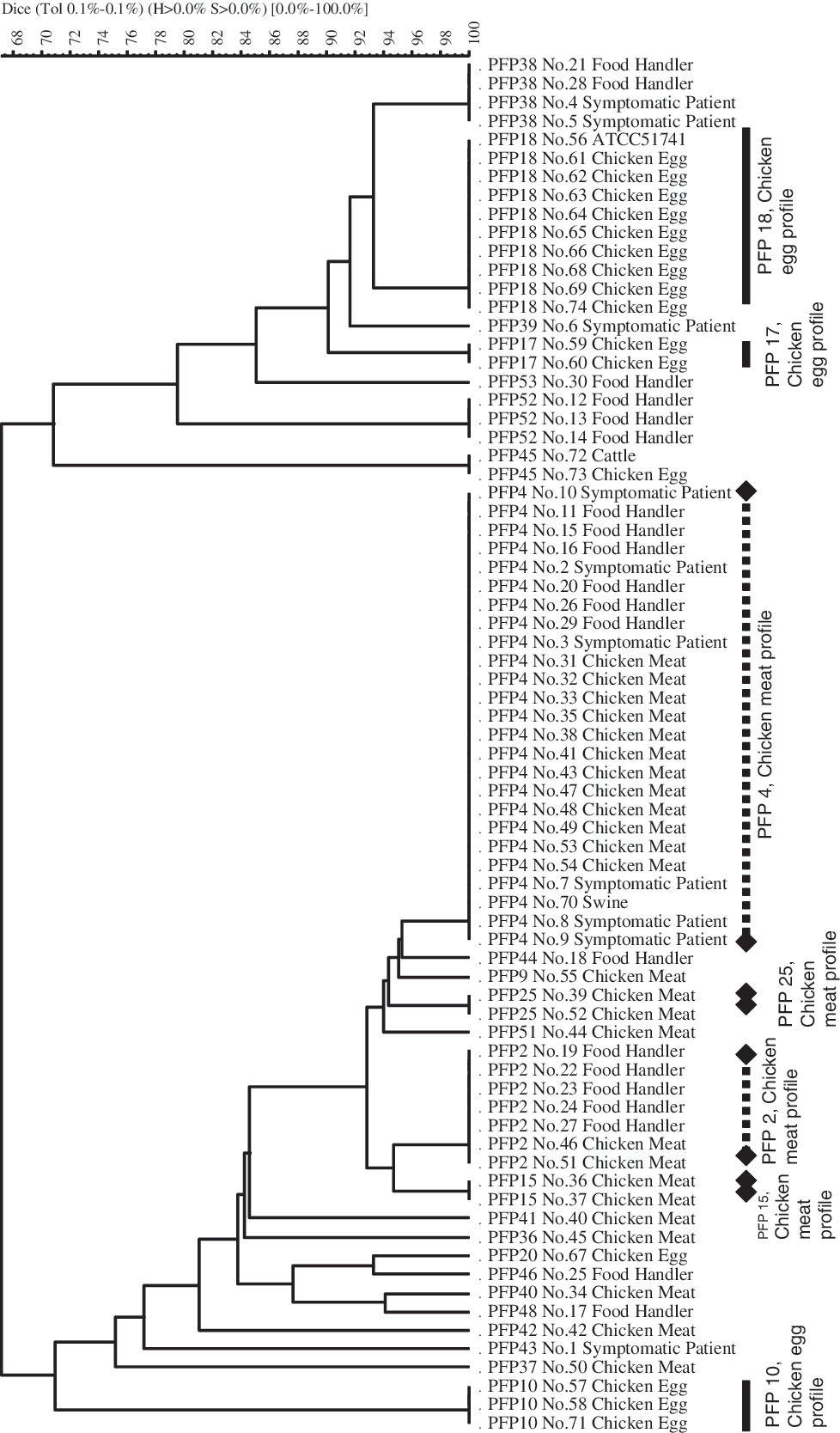
PFGE analysis of fingerprints from genetically defined Salmonella Infantis isolates. The numbers on the right of the dendrogram are pulsed-field profiles (PFPs) and isolate numbers (Table 2). The scale indicates the percentages of similarity, as determined by the Dice coefficients. PFPs 2 and 4 include isolates from humans and chicken meat (Table 2).

Schema of the PFPs of Salmonella Infantis isolates with BlnI digestion. Twenty-four different kinds of PFPs were obtained from the 74 isolates. The numbers on the left are the sizes of the fragments expressed in kbp. Numbers at the top indicate the PFPs. The chicken meat isolates belong to PFPs 2, 4, 9, 15, 25, 36, 37, 40, 41, 42, and 51. The chicken egg isolates belong to PFPs 10, 17, 18, 20, and 45. The human isolates belong to PFPs 2, 4, 38, 39, 43, 44, 46, 48, 52, and 53. There are only two PFPs (2 and 4) that include isolates from both humans and other source, and these profiles only include isolates from chicken meat and humans. Some PFPs of Salmonella Infantis (PFPs 1, 3, 5–8, 11–14, 16, 19, 21–24, 26–35, 47, 49, and 50) were assigned numbers in our previous study and are thus not consecutively numbered in this study.
Some profiles containing the isolates from either chicken egg or chicken meat were defined as specified profiles for further discussion. The chicken meat isolates belonged to PFPs 2, 4, 9, 15, 25, 36, 37, 40, 41, 42, and 51. In particular, PFPs 2, 4, 15, and 25 included two or more chicken meat isolates and were defined as “PFGE chicken meat profiles” for this study. A total of 36 isolates belonged to these profiles (Fig. 2). The chicken egg isolates belonged to PFPs 10, 17, 18, 20, and 45. In particular, PFPs 10, 17, and 18 included two or more chicken egg isolates and were defined as “PFGE chicken egg profiles.” A total of 15 isolates belonged to these profiles.
The results from the combination
The combination of the results of the two genetic fingerprinting methods yielded 47 combination types among the 74 isolates (e.g., PFP 2-AFLP profile 3 was defined as the combination type of 1) (Table 2). The dominant combination types were type 7 (PFP 4–AFLP profile 5), type 9 (PFP 4–AFLP profile 9), type 11 (PFP 4–AFLP profile 15), type 2 (PFP 2–AFLP profile 5), and type 24 (PFP 18–AFLP profile 17) consisting of 10, 5, 4, 3, and 3 isolates, respectively.
Four combination types (numbers 2, 7, 9, and 11) were present in human and chicken meat isolates simultaneously (a total of 8 human and 13 chicken meat isolates) (Table 2). None of the combination types consisted of both human and chicken egg isolates. The swine isolate was in one of the four combination types. There were no differences in the combination types between symptomatic patients and the food handlers.
Overall, 22 of 36 (61.1%) isolates within the PFGE chicken meat profiles belonged to the AFLP chicken meat profiles, and 22 of 30 (73.3%) isolates within the AFLP chicken meat profiles belonged to the PFGE chicken meat profiles. Further, 12 of 15 (80%) isolates within the PFGE chicken egg profiles belonged to the AFLP chicken egg profiles, and 12 of 23 (52.8%) isolates within the AFLP chicken egg profiles belonged to the PFGE chicken egg profiles. However, some isolates (numbers 9, 10, and 27) showed divergent connections; they belonged to both AFLP chicken egg profiles and PFGE chicken egg profiles.
Discussion
This study's results are contradictory to the results of Kudaka et al. (2006). In their study, it was reported that chicken meat is unlikely to be an infection source of Salmonella Infantis for humans. They found that the PFPs of 15 human isolates (taken from two hospitals) differed from those of 62 chicken meat isolates from a single prefecture in the Kyushu-Okinawa region, Japan, using PFGE. The limited geographical spread of the human source isolates from the two hospitals in their study may have contributed to the contradictory outcome. We compared the PFPs of 30 human isolates from numerous origins with those of nonhuman isolates from a wider geographical region comprising 13 prefectures inside and outside of the Kyushu-Okinawa region. In a study in The Netherlands, van Pelt et al. (1999) reported that approximately 21% of human salmonellosis cases were caused by chicken meat, of which the most dominant serovar was Salmonella Infantis. Their report supports our results that chicken meat plays a potentially significant role in human salmonellosis.
Our study had three major findings. First, some Salmonella Infantis isolates in humans were molecular-epidemiologically linked to some Salmonella Infantis isolates in chicken meat. Second, AFLP showed a relatively high suitability for application to epidemiological studies of Salmonella Infantis. Finally, none of the clonal lines (which were identical genotypic traits of the same combination types) consisted of both human and chicken egg isolates with the combined results of the two methods.
Taking the present molecular-epidemiological findings with PFGE and AFLP together with previous reports on the prevalence of Salmonella Infantis among poultry (Murakami et al., 2001) and Japanese eating habits, it is clear that chicken meat is an infection source of Salmonella Infantis in the Kyushu-Okinawa region, Japan. The combined results of the two methods, PFGE and AFLP, showed that some Salmonella Infantis isolates in humans were molecular-epidemiologically linked to some Salmonella Infantis isolates in chicken meat by determining clonal lines among different isolates. In the prevalence of Salmonella Infantis, the pathogen was frequently encountered, being isolated from 42.9% (15 of 35) of broiler farms and from 25.6% (23 of 90) of chicken meat samples in the Kyushu-Okinawa region (Murakami et al., 2001). In addition, some people eat raw chicken meat that is only slightly braised on the outside, “chicken sashimi,” as a delicacy in Japan. Further, according to the Food and Agriculture Organization of the United Nations (
Sources other than chicken meat were not investigated to determine if they might be a potential infection source of Salmonella Infantis for humans in this study. This study showed that there was a lack of common clonal lines presented between human isolates and chicken egg or cattle isolates; although only a small number of isolates were examined (some of the isolates were from outside of the Kyushu-Okinawa region). In sources other than chicken-related isolates, we believe that swine and pork meat are infection sources of Salmonella Infantis for humans because of the presence of common clonal lines presented between swine (an out of the Kyushu-Okinawa region isolate) and human isolates (clonal line no. 7). In fact, another study reported that Salmonella Infantis of swine origin was the cause of a large-scale outbreak of food poisoning in Denmark in 1993 (Wegener and Baggesen, 1996). Food sources other than chicken meat may also be infection sources of Salmonella Infantis, although these sources remain to be analyzed in future studies with more isolates from the same region.
AFLP showed a relatively high suitability for application to epidemiological studies of Salmonella Infantis. In general, determination of infection sources by means of molecular-epidemiological techniques requires robust fingerprinting methods. In this study, we investigated possible infection sources of Salmonella Infantis using a molecular-epidemiological approach that combined two fingerprinting methods, which both possess high discriminative power. Arbeit (1995) described the following five criteria for evaluating typing systems: typeability, reproducibility, discriminatory power, ease of interpretation, and ease of performance. In this study, AFLP is superior to PFGE in discriminatory power because the Simpson's index of AFLP (0.90) was better than that of PFGE (0.86) in this study. Regarding the other attributes of AFLP, typeability was 100%, and reproducibility was 86% in the preliminary experiments using three isolates. Further, the ease of interpretation did not differ from that of PFGE, and the ease of performance exceeded that of PFGE. Moreover, the association of more than 50% isolates into AFLP/PFGE chicken meat profiles or AFLP/PFGE chicken egg profiles were concurrent for both methods. These results reveal the relatively high reliability of AFLP, a novel PCR-based technique.
The combined usage of AFLP and PFGE has proven to be an advantageous molecular finger printing method for Salmonella (compared with singular usages of each method). Salmonella is a typical clonal population bacterium (Maynard Smith, 1995; Spratt and Maiden, 1999), so there is limited genetic diversity in their population compared with nonclonal population bacteria. Indeed, almost all of the Salmonella Infantis isolates in the Kyushu-Okinawa region showed the same sequence type (ST 32) with multilocus sequence typing (MLST) (part of data are available from MLST Databases at the ERI, University College Cork;
Footnotes
Acknowledgments
The authors express their gratitude to Dr. T. Yoshimura and Dr. Chijiwa, FIHES, for providing helpful advice. The authors also thank Mr. Y. Etoh, FIHES, for his help in creating the figures.
Disclosure Statement
No competing financial interests exist.