
Abstract
Seventeen phenolic compounds that are allowed to be used in the European food industry as aromatizants or antioxidants or that are naturally present in plants were tested for their ability to inhibit 19 strains of Staphylococcus aureus by using a standardized paper disc assay. Most of the strains assayed were foodborne (dairy and meat products). Human isolates and/or strains recommended for testing antimicrobial agents were also included in the study, and some of the test strains were enterotoxin producers. When the content was 200 μg/disc, various phenolic compounds had shown antimicrobial activity against all (hydroquinone, thymol, carvacrol, butylated hydroxyanisole, octyl gallate, and tannic acid) or most (gallic acid, propyl gallate, and ellagic acid) of the S. aureus strains tested. Significant differences in the inhibition zones (p < 0.05) among strains of the same, or similar, origin and among the different origins were observed for most of the phenolic compounds that showed antimicrobial activity for all or most of the strains tested.
Introduction
In addition to their antioxidant activity and other potential beneficial effects (e.g., anticarcinogenic and antiinflamatory activity) (Garrote et al., 2004), phenolic compounds have also been shown to possess antimicrobial activity against a wide range of microorganisms, not only bacteria, yeasts, and molds, but also viruses and protozoa (Buffo and Holley, 2006; Davidson and Taylor, 2007). With regard to bacteria, it is generally recognized that Gram positive are more sensitive than Gram negative to phenolic compounds (Dorman and Deans, 2000; Walsh et al., 2003; López-Malo Vigil et al., 2005). Staphylococcus aureus belongs to the first group of bacteria and is considered to be the main agent of staphylococcal food poisoning (Seo and Bohach, 2007). In spite of being one of the most prevalent causes of gastroenteritis worldwide, few investigations have been carried out to compare the effect of a large number of chemically identified phenolic compounds, both synthetic and natural, against a range of strains of S. aureus of different origins (dairy, meat, clinical, and so on) and with different virulence traits (enterotoxigenic and nonenterotoxigenic). Some studies have exclusively concentrated on single strains of S. aureus, and others are not directly comparable due to methodological differences such as choice of naturally occurring mixtures (e.g., essential oils), test micro-organism(s), and antimicrobial test method (Janssen et al., 1987; Hammer et al., 1999). Recent studies have revealed that the range in growth or inactivation characteristics can vary significantly within a species and that judgments on the safety of food processes must consider these ranges (Whiting and Buchanan, 2007).
Taking all this in account, the aim of the present study was to determine the susceptibility of S. aureus strains recovered from specific sources (food vs. clinical or other) to pure phenolic compounds (simple phenols, phenolic acids, and their derivatives, flavonoids, stilbenes and tannins), some of which were allowed to be used in the European food industry as aromatizers and antioxidants (Official Journal of the European Union, L 49 and L 204). We used them at the maximum concentration generally recommended in specific foods for synthetic antioxidants (200 mg/kg, based on the fat or oil content of the food product) (Codex STAN 192-1995, 2007). Here, we used a semiquantitative disk diffusion assay to select compounds with interesting activity for further study.
Materials and Methods
Bacterial strains and storage
Nine collection strains of S. aureus and 10 foodborne isolates (6 from bulk tank ewes' milk and 4 from rabbit carcasses, provided by Dr. M.R. García-Armesto and Dr. J.M. Rodríguez-Calleja, respectively, both from the Department of Food Hygiene and Food Technology, University of León, Spain) were used in this study. Additional information about the bacterial strains is given in Table 1. Stock cultures were maintained in eppendorfs with brain heart infusion (BHI) broth (Oxoid, Basingstoke, Hampshire, United Kingdom) in the presence of 30% v/v glycerol at −40°C.
In commercial strains, it is pointed out some applications and characteristics specified by the Culture Collections; in laboratory isolates, it is mentioned the enterotoxigenic character according to García-Armesto (1990) for ewes' milk strains and to Rodríguez-Calleja et al. (2006) for rabbit meat ones.
CECT, Colección Española de Cultivos Tipo; ATCC, American Type Culture Collection; CCM, Czech Colletion of Microorganisms; CCTM, Centre de Collection de Types Microbien, Université de Lausanne (Switzerland); MRSA, methicillin-resistant S. aureus.
Phenolic compounds and preparation of test disks
Carvacrol, eugenol, octyl gallate, and thymol were all obtained from Cymit Química, S.L. (Barcelona, Spain). Vanillin was obtained from Merck KGaA (Darmstadt, Germany). Tannic acid was obtained from Panreac Química, S.A. (Barcelona, Spain). Ellagic acid and gallic acid were obtained from Fluka and Riedel-deHaën® (Buchs SG, Switzerland). p-Hydroxybenzoic acid, protocatechuic acid, vanillic acid, BHA, BHT, hydroquinone, propyl gallate, quercetin, and resveratrol were obtained from Sigma-Aldrich (St. Louis, MO). Reference disks of gentamicin (10 μg/disc, code GM10) were purchased from Mast Group (Merseyside, United Kingdom).
Stock solutions were freshly prepared by dissolving 20 mg of a phenolic compound in 1 mL of absolute ethanol (for ellagic acid the dissolvent was isopropanol). The solutions were regularly mixed and sometimes heated (p-hydroxybenzoic and quercetin) at 60°C ± 1°C for 10–15 min. Stock solutions were protected from light exposure.
Sterilized blank 6-mm filter paper disks (BD Diagnostics, Sparks, MD) were soaked with 10 (L of each stock solution of phenolic compound, protected from light and dried to facilitate evaporation of the dissolvent. Negative control disks soaked only with ethanol or isopropanol and positive control commercial gentamicin discs were used in each experiment.
Activation and standardization of bacterial inocula
Frozen stock cultures were activated by transferring 20 μL into 4 mL of BHI broth and incubating for 18–24 h at 35°C ± 1°C. A loopful of broth culture was streaked onto BHI agar plates and incubated under the above-specified conditions. According to Barry (1976), at least three colonies of the same morphological type were touched on the top with a loop and the growth transferred to 4 mL of sterile saline solution (0.85% w/v of NaCl in distilled water). The turbidity of the actively growing bacteria was adjusted with sterile saline to obtain a turbidity visually comparable to the 0.5 McFarland standard (Remel, Inc., Lenexa, KS), which is equivalent to a bacterial concentration of 1 to 2 × 108 CFU/mL (Leiva, 1999).
Antimicrobial susceptibility testing
According to the standard method proposed by Barry (1976) a swab soaked with suspension was streaked onto the dried surface of a Mueller–Hinton agar (Oxoid) plate with a uniform depth of 4 mm. The same plate was swabbed twice more, rotating the plate 60° each time to ensure an even distribution of inoculum. Subsequently, the phenolic compound disks were applied on the inoculated agar plates and evenly distributed so that they were no closer than 24 mm from center to center (no more than five disks on a standard plate with a diameter of 90 mm). The plates were placed in an incubator set at 35°C ± 1°C within 15 min after the disks were applied. After 16–18 h of incubation the diameters of the zones of complete inhibition were measured, including the diameter of the disks. All tests were performed, at least, in duplicate and experiments were repeated twice on different days.
Statistical analysis
All data were expressed as means ± standard deviations of at least quadriplicate measurements. One-way analysis of variance was carried out to test any differences among individual strains (interstrain) included in each one of the groups obtained according to the origin of the strains, and among the groups (intergroups) obtained according to the origin of the strains.
Analysis of variance was performed by using the SPSS 15.0 package (SPSS Software available at the University of León). Significant differences (p < 0.05) between means were determined by post hoc Tukey method (assuming equal variances) or by post hoc Dunnet's T3 or the Games–Howell method (no assuming equal variances).
Results
Two of the simple phenols (eugenol and BHT) and four of the phenolic acids and their derivatives investigated (p-hydroxybenzoic acid, protocatechuic acid, vanillic acid, and vanillin) (Fig. 1) did not show antimicrobial activity against any of the strains tested. Conversely, four simple phenols (hydroquinone, thymol, carvacrol, and BHA), three phenolic acids and their derivatives (gallic acid, propyl gallate, and octyl gallate), and two hydrolysable tannins (ellagic acid and tannic acid) (Fig. 1) were effective against all or most of the strains. Finally, quercetin and resveratrol showed a limited inhibitory activity for only 5 (CECT 4459, CECT 5192, SA 49, SA 164, and SA 181) and 1 (SA 181) out of the 19 of S. aureus strains tested (results not shown), respectively. The nine phenolic compounds that exhibited antimicrobial activity against all or most of the S. aureus strains were studied in depth.
(

Susceptibility to natural and synthetic simple phenols
According to Table 2, the diameter of inhibition zones of S. aureus strains by natural (hydroquinone, thymol, and carvacrol) and synthetic (BHA) simple phenols ranged from 8.0 to 42.5 mm. Particularly, the strain CECT 4465 (from clinical origin and enterotoxin C producer) was the most resistant of the 19 strains tested in the study to all simple phenols.
Diameter of inhibition zones is expressed in millimeter (including the 6 mm of diameter of the paper disc).
Values are the mean of, at least, two experiments in duplicate ± standard deviation of the mean.
Data with the same letter are not significantly different among them (p > 0.05) for one origin and one antimicrobial compound.
SO, several origins; DO, dairy origin; MO, meat origin.
Hydroquinone was the simple phenol with more variable antimicrobial effect against the different strains tested (Table 2) with diameter of the inhibion zones ranging from 13 mm, for the most resistant strain (CECT 4465), to 42.5 mm for the most susceptible ones (172, 202, and SA 49). This variable behavior was also observed when the S. aureus strains were gathered according to their origin (Table 2) and the diameter of inhibition zones of the strains included into each of the groups showed significative differences (p < 0.05) among them.
For thymol and carvacrol, the differences in inhibition zones among strains were lower than for hydroquinone, ranging from 8.0 mm, for the more resistant strains (thymol and carvacrol against CECT 4465, and carvacrol against 181 and CECT 4466), to 14.0 mm (thymol, against 168 and 202) and 12.0 mm (carvacrol, against CECT 4013, 168 and 202) for the more susceptible ones (Table 2). This behavior was coincident with the lack of significant differences in the diameter of the inhibition zones among individual strains included in the group of several origin (clinical and other) and in the meat origin group by thymol, as well as in the meat origin group by carvacrol (Table 2).
BHA produced intermediate inhibition zones between hydroquinone and thymol/carvacrol, ranging from 10.5 mm, for the most resistent strain (CECT 4465), to 21.5 mm, for the most susceptible one (361) (Table 2). The strains included in each group that was obtained according to their origin (several, dairy, or meat) had shown significant differences (p < 0.05) among them in the diameter of the inhibition zones produced by this simple phenol (Table 2).
When we look at the mean value of the diameter of the inhibition zones of the strains included in each of the groups of strains gathered according to their origin (Fig. 2) for each of the simple phenol tested, we can see that thymol and carvacrol showed the lowest inhibitory activity against all the groups of strains, followed by BHA and hydroquinone. The statistical analysis of the mean inhibition zones for each group of strains revealed that there were no significant differences between the groups of several and meat origins for all simple phenols tested, as well as for the group of dairy origin for carvacrol. This group was significantly more susceptible (p < 0.05) to hydroquinone, thymol, and BHA than those of several origins and meat origin. In general, the differences between groups of S. aureus from different sources appear to be minor, although statistically significant.
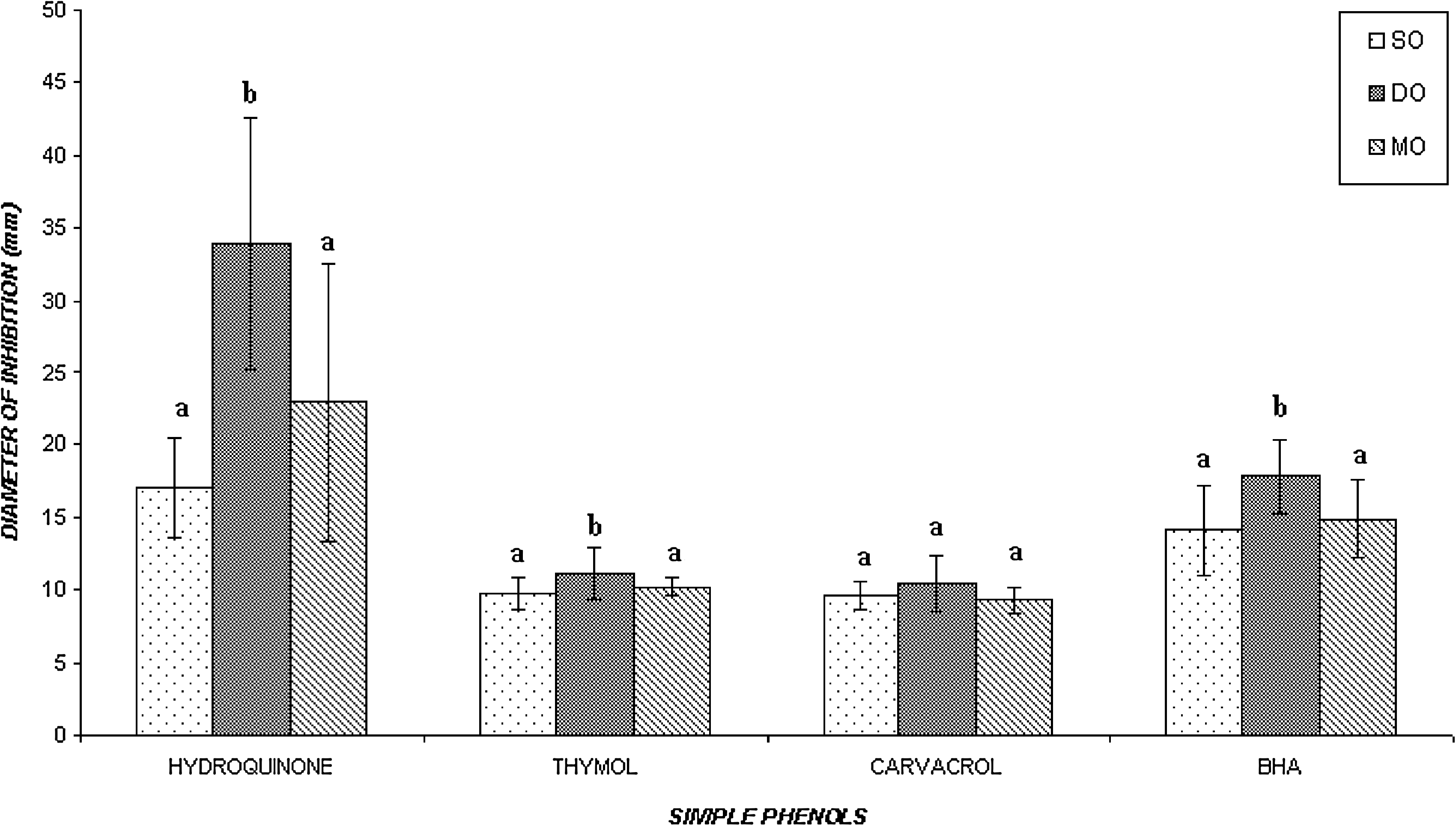
Inhibitory effect of simple phenols on groups of Staphylococcus aureus strains clustered according to their origin. Bar heights represent the mean of the diameters of the zones of inhibition for all the strains included in one group. Means with the same letter superscript are not significantly different (p > 0.05). Error bars indicate the standard deviation. SO, several origins; DO, dairy origin; MO, meat origin.
On the other hand, the statistical analysis of the mean inhibition zones obtained for each group of strains according to their enterotoxigenic character (data not shown) showed that for thymol and BHA there were significant differences (p < 0.05) between the groups of nonenterotoxigenic and enterotoxigenic strains, the latter being more resistant to these antimicrobials than the nonenterotoxigenic strains. Conversely, for carvacrol and hydroquinone there were no significant differences between both groups of strains (nonenterotoxigenic and enterotoxigenic).
Susceptibility to selected phenolic acids and their derivatives, and tannins
Table 3 shows the results of phenolic acids and their derivatives and hydrolysable tannins that were effective against all (octyl gallate and tannic acid) or most (gallic acid, propyl gallate, and ellagic acid) of the S. aureus strains tested. According to the table, the diameter of the inhibition zone ranged from 8.5 to 26.0 mm. These results suggest that the variations in the susceptibility of the different strains to phenolic acids and their derivatives were less marked than those obtained for the simple phenols studied (Table 2).
Diameter of inhibition zones is expressed in millimeter (including the 6 mm of diameter of the paper disc).
Values are the mean of, at least, two experiments in duplicate ± standard deviation of the mean.
Data with the same letter are not significantly different among them (p > 0.05) for an origin and an antimicrobial compound.
Antibiotic used as control at the concentration of 10 μg/disc.
ND, nondetected inhibition.
Gallic acid and alkyl gallates showed a variable antimicrobial effect against the different strains tested (Table 3); this heterogenicity was higher for gallic acid than for propyl gallate. Both (gallic acid and propyl gallate) were more heterogeneous than octyl gallate. According to this, the diameter of the inhibition zones ranged from 11.3 to 26.0 mm, 9.3 to 22.0 mm, and 14.0 to 21.5 mm, for gallic acid, propyl gallate, and octyl gallate, respectively. Also, the statistical analysis of the inhibition zones of the strains included in each origin group (Fig. 3) confirmed a greater variability inside all groups (interstrains) for gallic acid than for their derivatives (propyl gallate and octyl gallate). Particularly, there were no significant differences (p < 0.05) in the inhibition zones obtained by octyl gallate in six out of the seven strains included in the group of dairy origin as well as in the meat origin group.
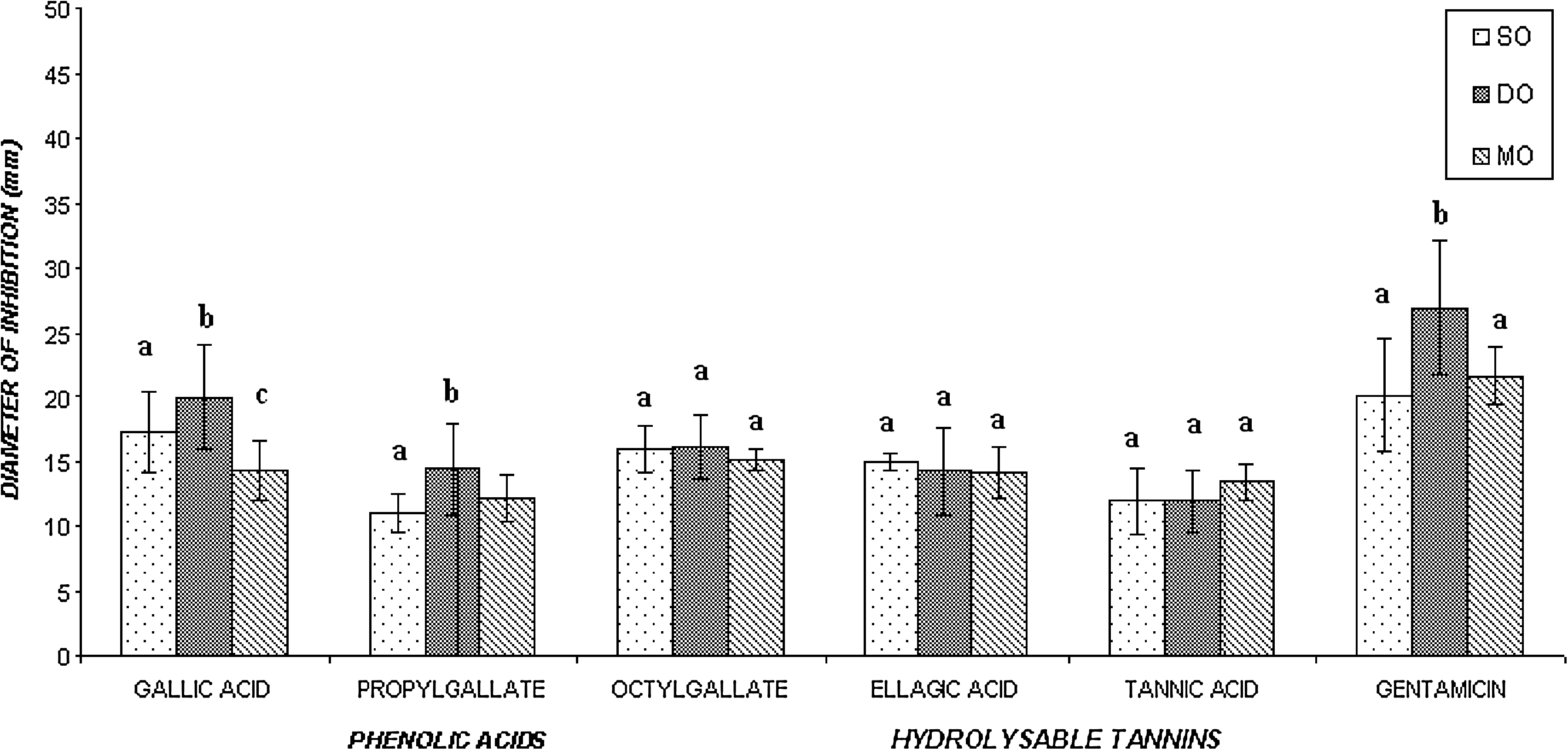
Inhibitory effect of phenolic acids and gentamicin on groups of S. aureus strains clustered according to their origin. Bar heights represent the mean of the diameters of the zones of inhibition for all the strains included in one group. Means with the same letter superscript are not significantly different (p > 0.05). Error bars indicate the standard deviation.
With regard to ellagic acid, this compound was not inhibitory for 4 (168, 172, 180, and 202) out of the 19 strains tested, and the inhibition zones of the susceptible ones ranged from 11.0 mm (181 and SA 49) to 17.5 mm (CECT 4013 and SA 164) (Table 3). Conversely, all strains of S. aureus tested were susceptible to tannic acid, showing inhibition zones that ranged from 8.5 mm for the most resistant strain (CECT 59) to 16.5 mm for the most susceptible one (168) (Table 3). When the strains were clustered according to their origin there were no significant differences (p < 0.05) between the inhibition zones ellicited by ellagic acid on the strains included in the group of strains of several origins, but there were for the inhibition zones observed in the susceptible strains included into the two other groups (dairy and meat origin) (Table 3). The strains from dairy origin showed a variable behavior against the ellagic acid (four of them being resistant, and three susceptible).
When we compared the mean results of inhibition zones produced by phenolic acids and their derivatives on the strains included in each of the three groups generated according to their origin (Fig. 3), we observed that gallic acid and octyl gallate were the most efficient of the phenolic acids and their derivatives tested to inhibit the strains of S. aureus. On the other hand, there were no significant differences (p < 0.05) between the mean of inhibition zones of strains gathered according to their origin (several, dairy, and meat) for octyl gallate and tannic acid, whereas for gallic acid and propyl gallate the group of dairy origin was more susceptible than the other two groups (several and meat origin). With regard to ellagic acid, although there were no significant differences between groups of strains according to their origin, it is remarkable that the results corresponding to the four strains of dairy origin that were resistant to this compound were not included in the histogram. Also here, as it was indicated above for simple phenols, the differences between groups of S. aureus from different sources appear to be minor, although statistically significant.
We also observed that the enterotoxigenic groups (regardless of the type of enterotoxin produced) did not show significant differences (p < 0.05) against octyl gallate (data not shown). With respect to the rest of the phenolic compounds, the group of toxin D/E producer strains was significatively more resistant to tannic acid and propyl gallate, whereas gallic acid was most effective against the group of enterotoxin C producer strains. Finally, the strains most susceptible to ellagic acid were the enterotoxin B or C producers (data not shown).
Gentamicin was a stronger inhibitor than all the phenolic acids and the simple phenols, except hydroquinone, which showed higher values of diameter of inhibition than those of gentamicin for some strains in the groups of dairy origin and meat origin. The most resistant strain against the antibiotic was CECT 4465 included in the group of several origins, which is also an enterotoxin C producer. On the contrary, strain 168 of dairy origin and an enterotoxin A producer was the most susceptible. A higher variability among strains was recorded in the group of dairy origin. In general, the group of dairy origin was the most susceptible to gentamicin, whereas with regard to the enterotoxigenic groups, the enterotoxin A producer was the most susceptible and the enterotoxin B producer was the most resistant (data not shown).
Discussion
A content of 200 μg/disc is the maximum amount generally permitted for synthetic phenolic compounds used as antioxidants in foods (López-Malo Vigil et al., 2005; Codex STAN 192-1995, 2007). At this content, six compounds (eugenol, BHT, p-hydroxybenzoic acid, protocatechuic acid, vanillic acid, and vanillin) had no activity under the tested conditions. Also, limited activity was recorded for quercetin and resveratrol. Reasons for the weak activity of these compounds could include (1) lack of solubility in aqueous media and, hence, poor diffusion through the agar; (2) dissociation of the organic acids and, hence, loss of activity, and (3) loss of the compounds in the gas phase. Solubility could affect the diffusion of the compounds from the disk through the agar, but we obtained different results of inhibition, regardless of this. So, the ineffective or less effective compounds (eugenol, BHT, propyl gallate, elagic acid, quercetin, and resveratrol) show bad solubility, as do some effective compounds of this study, such as BHA and octyl gallate. In fact, the addition of alkyl groups to a molecule decreases the solubility, whereas the addition of a hydroxyl group or a system of delocalized electrons to a molecule increases the solubility (Ultee et al., 2002). To overcome this factor of variation in the stock solutions of the pure phenolic compounds tested, we dissolved all of them in absolute ethanol or isopropanol.
About the deprotonation of the phenolic compounds at the pH of the test medium (6.6), only the acids showed pKa values under this pH value, which would not be an important feature in the loss of activity, as the pKa value of p-hydroxybenzoic and protocatechuic acids (ineffectives) is 4.48, very close to the pKa value of gallic acid (4.41), which is an effective compound. On the other hand, the simple phenols would remain mostly undissociated at the pH of the medium, as the pKa of phenolic compounds is already 10 (Machleidt et al., 1972), but the effect was very different for the phenols used; for instance, hydroquinone (pKa of 10.35) was the most effective phenolic compound tested, but others showed null (eugenol and BHT) or minimum (thymol and carvacrol) activity. It is reported that the undissociated form of the acid is primarily responsible for inhibition of microorganisms (Davidson and Taylor, 2007). Eklund (1983), however, demonstrated that while the undissociated species has significantly greater activity, the anion does contribute slightly to antimicrobial activity.
Particularly for eugenol, Moleyar and Narasimham (1992) also reported that at concentrations of 250 and 500 μg/mL in nutrient agar, this compound was not inhibitory to Staphylococcus spp.; however, at concentrations of 750 and 1000 μg/mL, it was active in inhibiting the organism by 15 days and more than 30 days, respectively. This lack of effectivity of eugenol in comparison with other simple phenols tested in our study could be related to its chemical structure—particularly, to the presence of a methoxy group in ortho-position—that would make the release of a proton from the hydroxyl group difficult (Arfa et al., 2006), reducing the expression of the acidic character of this compound and, then, its antimicrobial activity.
As for BHT, Tunçel and Nergiz (1993) found that the minimum inhibitory concentration (MIC) value of BHT against S. aureus was 400 μg/mL. This result will support our findings and confirm the lack of inhibitory effect of BHT on S. aureus at the concentration of 200 μg/disc. Conversely, BHA, a structurally related compound, was able to inhibit all strains of S. aureus under the same conditions as BHT. According to our results, and those reported by other authors using a single test strain of S. aureus (Davidson et al., 1981; Moleyar and Narasimham, 1992; Tunçel and Nergiz, 1993), concentrations of BHA above 100–150 μg/mL of nutritive, solid or liquid, media would be inhibitory to S. aureus, and could therefore be used as an effective antimicrobial against this microorganism at the maximum concentration generally allowed to be added as antioxidant in particular foods (Codex STAN 192-1995, 2007), or even less. In relation to the different effect of BHT and BHA, Rico-Muñoz et al. (1987) found that the BHA stimulated and the BHT had no effect on the activity of the ATPase of membrane of two strains of S. aureus, at concentrations of 0, 150, 300, 600, and 1200 μg/mL. According to these authors, the different effect on the ATPase of membrane could be connected with the different molecular structure and properties of both phenolic compounds, which would determine the lesser ability of BHT to form relative stable aqueous suspension in the concentration range used. Consequently, too low a concentration of BHT would diffuse in the agar media to react either with the lipid of the cell membrane or the enzyme itself (Rico-Muñoz et al., 1987). Also, the difference in the solubility of BHA and BHT probably accounts for the observed effect.
With regard to the other simple phenols tested in our study, the data obtained for inhibition by hydroquinone are relevant, as we have not found in the published bibliography any study of the effect of this compound on S. aureus or, even, on other Gram-positive or Gram-negative bacteria. However, several studies of carvacrol and thymol have been published recently, according to which they are considered strong broad-spectrum antimicrobials (Ultee et al., 2002; Davidson and Taylor, 2007; Falcone et al., 2007), also with the ability to be used to control biofilm formation of S. aureus (Knowles and Roller, 2001; Knowles et al., 2005). According to our results, thymol and carvacrol, particularly, the former, were the least effective (smaller diameter of the inhibition zones) but most homogeneous (less significant differences among interstrains included in the groups and intergroups of strains clustered according to their origin) simple phenols against S. aureus of those that produced inhibition of this microorganism under the assayed conditions. One explanation for our findings could be related to the molecular structure of both isomeric compounds (Fig. 1), with a nonpolar moiety (which might hinder their diffusion through the agar, thus reducing the inhibition zones generated) and a hydroxyl group and a system of delocalized electrons that confer an acidic character to these molecules. For this reason, thymol and carvacrol could release protons inside the cells, causing a cytoplasmic acidification. The maintenance of internal pH homeostasis implies the existence of an electrochemical gradient of H+, generated by a plasma membrane H+-ATPase, which would cause a depletion of the ATP pool and eventually lead to cell death, as it has been proposed previously for carvacrol (Ultee et al., 2002; Knowles et al., 2005). On the other hand, our results are in agreement with those of Falcone et al. (2007), who, working with S. aureus ATCC 25923, reported that the upper noninhibitory concentration of thymol for this strain was 44.99 μg/mL. Also, Juven et al. (1994) found that thymol was inhibitory against one enterotoxin producer strain of S. aureus at concentrations in the 175–250 μg/mL range.
Concerning the phenolic acids and their derivatives assayed, p-hydroxybenzoic acid, protocatechuic acid, vanillic acid, and vanillin were not effective against S. aureus at the content of 200 (g of test compound per disc; however, gallic acid and alkyl gallates (propyl gallate and octyl gallate) were effective, under the same conditions, against nearly all the S. aureus strains tested. It has been reported that the site(s) and number of hydroxyl groups at the benzoic ring could be related to their relative toxicity to microorganims, with evidence that increased hydroxylation results in increased toxicity (Cowan, 1999). The difference in the chemical structure of the above-mentioned phenolic acids and derivatives (number of hydroxyl groups) could contribute to the different effect observed against S. aureus. Further, Fernández et al. (1996), using a disk diffusion method, reported that it was necessary to use amounts per disk higher than 200 μg/mL of phenolic fractions obtained from Scrophularia frutescens and Scrophularia sambucifolia to detect inhibition or appreciable inhibition of the S. aureus growth. These fractions contained vainillic, p-hydroxybenzoic, and protocatechuic acids, among other phenolic compounds, but not gallic acid or gallic derivatives.
Kubo et al. (2004) also found that six selected Gram-positive bacteria (including one strain of S. aureus) were susceptible to alkyl gallates, and this activity was found to correlate with the alkyl chain length. According to the mentioned authors, using a broth dilution method, gallic acid was less effective than alkyl gallates against Bacillus subtilis. It is possible that in our results the more hydrophobic character of the alkylled gallic acid derivatives hindered the diffusion of these compounds from the disks into the solid media, thus reducing the size of inhibition zones of S. aureus in comparison with those obtained with gallid acid. Additionally, our results coincide with those of Kubo et al. (2002, 2004), who found that propyl gallate was less inhibitory than octyl gallate for Salmonella cholerasuis and B. subtilis, respectively. In our study, the greater difficulty in the diffusion of octyl gallate in comparison with propyl gallate (length of alkyl chain of C8 and C3, respectively) might have been overcome by the higher antimicrobial potency of octyl gallate (it has been reported that the MIC for this compound is one or more logarithm unit less than that of propyl gallate against B. subtilis) (Kubo et al., 2004). On the other hand, it is interesting to note that the only strain that was not sensitive (ATCC 19095) and the strain that showed lower inhibition (ATCC 6538P) to propyl gallate in our study were, in both cases, from clinical origin (being isolated from a leg access and a human lesion, respectively).
With regard to ellagic acid, Chung et al. (1993) reported that this compound did not inhibit any of 15 different genera and species of Gram-positive and Gram-negative bacteria tested, including the CECT 240/ATCC 6538P strain of S. aureus, at the amount of 200 (g of ellagic acid/well and using distilled water as the solvent of the phenolic compound. In the present study, the mentioned strain was inhibited at the same initial content (200 (g) by the disk diffusion test and using isopropanol as solvent, but four isolates of S. aureus of dairy origin were not inhibited. Akiyama et al. (2001) reported that the ellagic acid showed an MIC value of 8000 μg/mL against all of the 18 strains of S. aureus tested isolated from infections in human skin (foruncle lesions and impetigo).
Tannic acid is an ester of gallic acid and glucose, which, in our study, was inhibitory for all the strains of S. aureus tested showing slightly lower mean diameters of inhibition zones than those obtained for gallic acid. However, other authors working with various genera and species of Gram-positive and Gram-negative bacteria (Henis et al., 1964; Payne et al., 1989; Chung et al., 1993) reported that tannic acid was more inhibitory than gallic acid. Again, it is possible that the different methodology used in our study in comparison with those of the other studies and the different structure of both compounds (gallic and tannic acids) might have affected the different results. Concerning the inhibitory concentration of tannic acid for S. aureus, our results with the CECT 240/ATCC 6538P strain are in good agreement with those reported by Chung et al. (1993), who found that the same strain was inhibited by a concentration of 200 (g of tannic acid/well. On the other hand, Akiyama et al. (2001) studying 18 strains of S. aureus isolated from infections in human skin (foruncle lesions and impetigo) observed that tannic acid showed an MIC value of 250 μg/mL for 14 out of the 18 strains tested and of 1000 μg/mL for the rest of the strains. Also, Amarowicz et al. (2008), using a broth dilution method, reported for S. aureus (ATCC 25923) (of clinical origin) an MIC value ranging from 250 to 500 μg/mL, depending on the plant source of tannin fraction tested. It is possible that, in addition to other methodological factors, the clinical origin (and probably exposure to antibiotic treatments) of the strains studied by Akiyama et al. (2001) might have contributed to the higher resistance to tannic acid of these strains in comparison with our results, although this hypothesis would need further verification. At the moment, it is known that S. aureus is able to acquire genetic resistance to heavy metals and antimicrobial agents used in clinical medicine; however, the resistance of this bacterium to common food preservation methods is generally considered unremarkable (Seo and Bohach, 2007).
In the present study, quercetin could only inhibit 5 out of 19 strains of S. aureus tested at the amount of 200 μg/disc. Our results are in agreement with those reported by Rauha et al. (2000), who observed, by using a well diffusion agar assay modified, that quercetin showed a clear antimicrobial activity against one strain of S. aureus (DSM 20231) and two strains of Staphylococcus epidermidis (ATCC 12228 and FOMK) at the concentration of 500 μg/well. With regard to the effect of quercetin against other Gram-positive bacteria, Payne et al. (1989), using an agar dilution technique, reported that quercetin was inhibitory against eight strains of Listeria monocytogenes, but showed MIC values higher than 205 μg/mL.
The fact that several strains of S. aureus showed differences in sensitivity toward most of the phenolic compound assayed in this study should be taken into account when one of those compounds was intended to be used to control the growth of this species. Other authors (Puupponen-Pimiä et al., 2001) studying the antimicrobial activity of 17 pure phenolic compounds representing flavonoids and phenolic acids against selected Gram-positive (lactic acid bacteria) and Gram-negative bacteria (Escherichia coli and Salmonella) have also found that different strains of the same bacterial species showed different sensitivities to one flavonoid. Also, several modeling studies have reported that variability between strains of the same species is greater than the variability due to experimental error observed in studies with single strains or cocktails of strains and that this variability can have profound effects on the outcomes of risk assessments (Ross, 2008).
Experiments in progress to estimate MIC and MBC are being carried out to check the kind of inhibitory effect (bacteriostatic or bactericidal) displayed by the most consistent phenolic compounds against S. aureus and to test their relative activity and how long the inhibition can be expected in foods.
Conclusion
The results of this study indicate that some simple phenolic compounds, such as hydroquinone, thymol, carvacrol, BHA, as well as octyl gallate and tannic acid, could be used as antimicrobial additives for inhibiting the growth of foodborne S. aureus strains. These positive inhibition results might be the basis for further studies to apply these antioxidants in hurdle technology, combined with other inhibiting factors to obtain a gentle, but effective control of this pathogen in a variety of foods. However, the selection of effective concentrations of phenolic compounds should be carefully selected, particularly to control the growth of enterotoxigenic strains.
Footnotes
Acknowledgment
The present study was financially supported by research grant AGL2005-01793/ALI from the Dirección General de Investigación Científica y Técnica (Scientific and Technical Research Council) of the Spanish Ministry of Science and Technology.
Disclosure Statement
No competing financial interests exist.