
Abstract
Listeriosis is a severe infection that mainly affects pregnant women, neonates, and immuno-compromised adults. The commercially available semi-automated repetitive-sequence-based polymerase chain reaction assay system, DiversiLab, has been successfully used for subtyping several species of bacteria. In this article we compare the DiversiLab System with macrorestriction analysis by pulsed-field gel electrophoresis (PFGE), which is currently the gold standard for molecular subtyping of Listeria monocytogenes. We used a panel of 116 human and food L. monocytogenes isolates for the comparative evaluation. Among these isolates, there were 4 pairs of duplicates, 13 strains were epidemiologically related, and the remaining food isolates were epidemiologically unrelated. The isolates of different serotypes represented distinct DiversiLab types (DTs) and ApaI/AscI-PFGE types except for one DT-containing isolates of two serotypes, 4b and 1/2b. The four duplicates displayed the same DT and ApaI/AscI PFGE type demonstrating the good reproducibility of the two methods. The epidemiologically related strains were clustered in the same DT and PFGE type. The Simpson's index of diversity was 0.954; 0.988; 0.994; and 0.998 for DiversiLab, AscI-PFGE, ApaI-PFGE, and AscI/ApaI-PFGE, respectively. Thus, PFGE was more discriminating than DiversiLab. However, for 1/2a serotype strains, six AscI-PFGE, three ApaI-PFGE, and one ApaI/AscI PFGE type were divided into different DTs. DiversiLab enabled a good discrimination between serotype 1/2a strains. DiversiLab is less labor intensive than PFGE and provides results in <24 hours compared with 30 hours to 3 days for PFGE from the time a pure culture of the bacteria has been obtained. On the basis of these results, DiversiLab may be useful for tracking the source of contamination in food-processing facilities and their environments. Also, DiversiLab may be more appropriate for long-term epidemiological studies where less discrimination is needed.
Introduction
L
The causative organism of listeriosis is Listeria monocytogenes, which is mainly transmitted to humans through contaminated food. L. monocytogenes is a significant foodborne pathogen given its widespread distribution in nature, its ability to survive under a wide range of environmental conditions, its resistance to acid and high salt concentrations, and its ability to grow at refrigeration temperatures. L. monocytogenes can contaminate foods at each step of the food supply chain. Infections have been associated with various food products, such as unpasteurized milk, soft cheeses, ice cream, raw vegetables, fermented raw-meat sausages, raw and cooked poultry, raw meats, cold cuts, and raw and smoked fish (Lindstedt et al., 2008).
Out of the several molecular methods currently available, macrorestriction analysis by pulsed-field gel electrophoresis (PFGE) is one of the most discriminatory and reproducible methods for the subtyping of L. monocytogenes (Brosch et al., 1996; Kerouanton et al., 1998). The combination of restriction endonucleases AscI and ApaI has shown excellent discrimination for L. monocytogenes (Buchrieser et al., 1993; Brosch et al., 1996). PFGE is considered to be the standard subtyping method (Graves and Swamminathan, 2001). Although the protocol has been shortened to 30 hours from the time a pure culture of the bacteria has been obtained (Graves and Swamminathan, 2001), PFGE remains a manual and labor-intensive subtyping method.
Repetitive-sequence-based polymerase chain reaction (rep-PCR) has been successfully used for differentiating between L. monocytogenes strains isolated from humans, animals, and food (Jersek et al., 1999). Rep-PCR uses primers that target noncoding short repetitive sequence elements interspersed throughout the genome, whose function is probably associated with gene regulation (Harvey et al., 2004). The amplified DNA fragments, when separated by electrophoresis, constitute a genomic fingerprint that can be employed for bacterial discrimination. It has also been successfully performed for typing L. monocytogenes from ready-to-eat food products (Van Coillie et al., 2004) and isolated from cheese (Rijpens et al., 1998). This method has also been successfully applied to a set of L. monocytogenes food isolates obtained over an extended period from different food processors (Harvey et al., 2004). These authors have also demonstrated good reproducibility in their laboratory and good stability of rep-PCR types obtained. The discriminatory ability of the rep-PCR was shown to be similar to that of PFGE for typing L. monocytogenes isolates from seafood and humans (Chou and Wang, 2006).
The commercially available semi-automated rep-PCR assay system, DiversiLab (BioMérieux S.A.), is particularly useful because it eliminates the need for post-PCR processing steps. It enables a standardized method in comparison with manual gel-based rep-PCR. Previous investigations have described rapid strain-level discrimination using DiversiLab for different bacteria such as Salmonella enterica (Wise et al., 2009), Acinetobacter baumanii (Carretto et al., 2008; Fontana et al., 2008), Staphyloccocus aureus (Ross et al., 2005; Shutt et al., 2005; Moroney et al., 2007), and Shigella sonnei (Boumghar et al., 2008). For the characterization of L. monocytogenes, only one investigation used this technology: In this study, the authors have demonstrated that the majority of 1/2a isolates from a 61 isolates collection from raw milk samples were discriminated from the remaining serotypes (Van Kessel et al., 2005). Those results were not compared with more established methods such as PFGE.
The objective of this study was to evaluate the usefulness of the DiversiLab System in differentiating a panel of L. monocytogenes strains, including epidemiologically related and unrelated strains. The results were compared with those obtained by PFGE.
Materials and Methods
Strains
This study included 34 strains previously used in the World Health Organization (WHO) international multicenter L. monocytogenes subtyping study (Bille and Rocourt, 1996). These strains (TS02-TS77) were labeled TS (“Test study”) by Schönberg et al. (1996). This panel included 25 sporadic or epidemic human strains and 9 food strains, including 4 strains represented by 2 duplicates each (TS32,TS72; TS56,TS77; TS35,TS75; and TS63,TS73), 13 strains related to six different epidemiogical groups (03, 05, 15, 16, 19, and 22) and additional strains that were not epidemiologically related (Table 1). Eighty-one nonhuman field isolates, collected from food analysis laboratories, in the context of monitoring, surveillance sampling, or research projects, were also included. These isolates were collected from food and food-processing environment: milk and cheese products, fish and seafood, beef, pork, poultry, goat, lamb, pork, food industry, and sewage (Table 2). A French epidemic human strain of 1/2a serotype (CLIP 95058), provided by the National Reference Center of Listeria (Institut Pasteur), was also tested.
TS, test study; DT, DiversiLab type.
The number of strains was indicated within parentheses.
The food channel was unknown.
This DT included the auto-agglutinable strain.
PFGE, pulsed-field gel electrophoresis.
Serotyping
Species were identified using agar Listeria Ottaviani Agosti (ALOA) plates (AES) and Christie Atkins Munch-Petersen (CAMP) test (McKellar, 1994). Each strain was serotyped by agglutination using commercially available antisera (Denka, Eurobio), after adapting the manufacturer's instructions and using the procedures outlined by Seeliger and Höhne (1979). The laboratory has been certified by the French Accreditation Committee (COFRAC) for this serotyping method as an internal method (Accréditation No. 1-0245, Section Laboratories,
Rep-PCR
DNA extraction
DNA was extracted with the UltraClean™ Microbial DNA Isolation Kit (Mo Bio Laboratories) following the manufacturer's instructions. Extracts were adjusted to approximately 25 ng/μL using a spectrophotometer (Biophotometer, Eppendorf ).
DNA amplification
DNA samples were amplified by rep-PCR methodology using the DiversiLab Listeria kit for DNA fingerprinting following the manufacturer's instructions. Briefly, 2 μL of genomic DNA, 0.5 μL of AmpliTaq polymerase (Applied Biosystems), 2 μL kit-supplied primer mix, and 2.5 μL of GeneAmp 10× PCR Buffer (Applied Biosystems) were added to 18 μL of the kit-supplied rep-PCR master mix for a total of 25 μL/PCR reaction mixture. PCR was performed on a thermal cycler (GeneAmp PCR System, 9700, PE; Applied Biosystems) using the following parameters: initial denaturation of 94°C for 2 minutes, followed by 35 cycles of denaturation at 94°C for 30 seconds, annealing at 50°C for 30 seconds, extension at 70°C for 90 seconds, and a final extension at 70°C for 3 minutes.
Detection of amplification products
The automated microbial fingerprinting system (DiversiLab System) was used to detect DNA fingerprints of each isolate. The amplified fragments of various sizes and intensities were separated and detected using a microfluidic labchip with an Agilent 2100 Bioanalyzer. Each run, performed on a single chip, consisted of one positive control, one negative control, one ladder control, and 10 samples.
Data analysis
Data were analyzed using DiversiLab Software version 3.3. As many observed patterns were characterized by the presence of two or three major bands in the fingerprint pattern and many other small peaks, the Kullback–Leibler method was used to calculate similarity percentages and to define a DiversiLab type (DT), as recommended by the manufacturer. The relatedness was determined by cluster analysis and according to the manufacturer's guidelines. Since it is important to take into account both peak presence/absence and intensity for rep-PCR fingerprint analysis, the gel image comparison was sometimes not sufficient to detect the variation among isolates. Thus, the electrophoregrams and graph overlays obtained for each of the 116 isolates were also used to check peak differences and intensity differences. The Unweighted Pair Group Method with Arithmetic mean was used as a clustering method.
Pulsed-field gel electrophoresis
PFGE was performed using the standard CDC PulseNet protocol (Graves and Swamminathan, 2001) with minor modifications. Each strain was grown overnight in 10 mL of trypton soy agar yeast extract (TSAYE) instead of brain heart infusion agar (BHIA). For the DNA digestion step in agarose plugs with ApaI and AscI, 1 μL of each enzyme was used instead of 5 μL. The incubation timing of plugs with the enzymes mixtures was 4 hours instead of 5 hours, as advised in the PulseNet protocol.
Gels were then stained with ethidium bromide and banding patterns were observed under UV light, using the Gel Doc Eq system and Quantity One software (Bio-Rad). DNA patterns were analyzed with BioNumerics software (V 5.01; Applied Maths). Algorithms available within the program were used to compare patterns. Dendrograms were produced, using the Dice coefficient and Unweighted Pair Group Method with Arithmetic mean, with a 1% tolerance limit and 1% optimization. The recommendations of Barrett et al. (2006) were also followed for the gel analysis: gels including partial digestions or in which the bands were not clear were ran again. Each PFGE type differing by at least one band from a previously recognized type was considered to be a new type. Each new type was given a unique identification and added to the library of PFGE types. All the bands with sizes <33 kB were systematically removed. The laboratory has been COFRAC for PFGE analyses (Accréditation No. 1-0245, Section Laboratories,
Reproducibility of the subtyping methods
The reproducibility of the DiversiLab and PFGE methods can be determined from the results obtained with the four TS strains represented by two duplicates each and the epidemiologically related strains included in this study.
For DiversiLab, the four TS strains represented by two duplicates each was tested with two different chips. Moreover, for eight isolates, DNA from the same extraction was tested with two different chips.
Discriminatory power of the subtyping methods
The ability of each method to discriminate between subtypes was assessed in two ways: The ability of the method to recognize the homogeneity of strains inside a given group of epidemiologically related strains. Its capacity to discriminate between groups and/or between unrelated strains by calculating the Simpson's index of diversity (ID) (Hunter and Gaston, 1988). The ID was calculated on the basis of 105 strains, including the 82 isolates, 1 TS strain from each of the six epidemiologically related groups, and additional TS strains not epidemiologically related.
Results
Serotyping
Of the 82 analyzed isolates, 81 were successfully serotyped. Only one isolate was found to be auto-agglutinable. The collection could be divided into five serotypes: 1/2c (6 isolates), 3a (2 isolates), 1/2b (7 isolates), 4b (22 isolates), and 1/2a (44 isolates). Serotype 1/2a was predominant (54%).
Typing results
Reproducibility of the subtyping methods
The eight isolates evaluated on two distinct chips displayed a same DT (Fig. 1, Table 1). The DTs and combinated ApaI/AscI PFGE types were indistinguishable for the four duplicate TS strains (TS32,TS72; TS56,TS77; TS35,TS75; and TS63,TS73) (Fig. 1; Table 1). The DTs and PFGE types were correlated with the six epidemiological groups (03, 05, 15, 16, 19, and 22) for the 13 tested TS strains (Table 1).
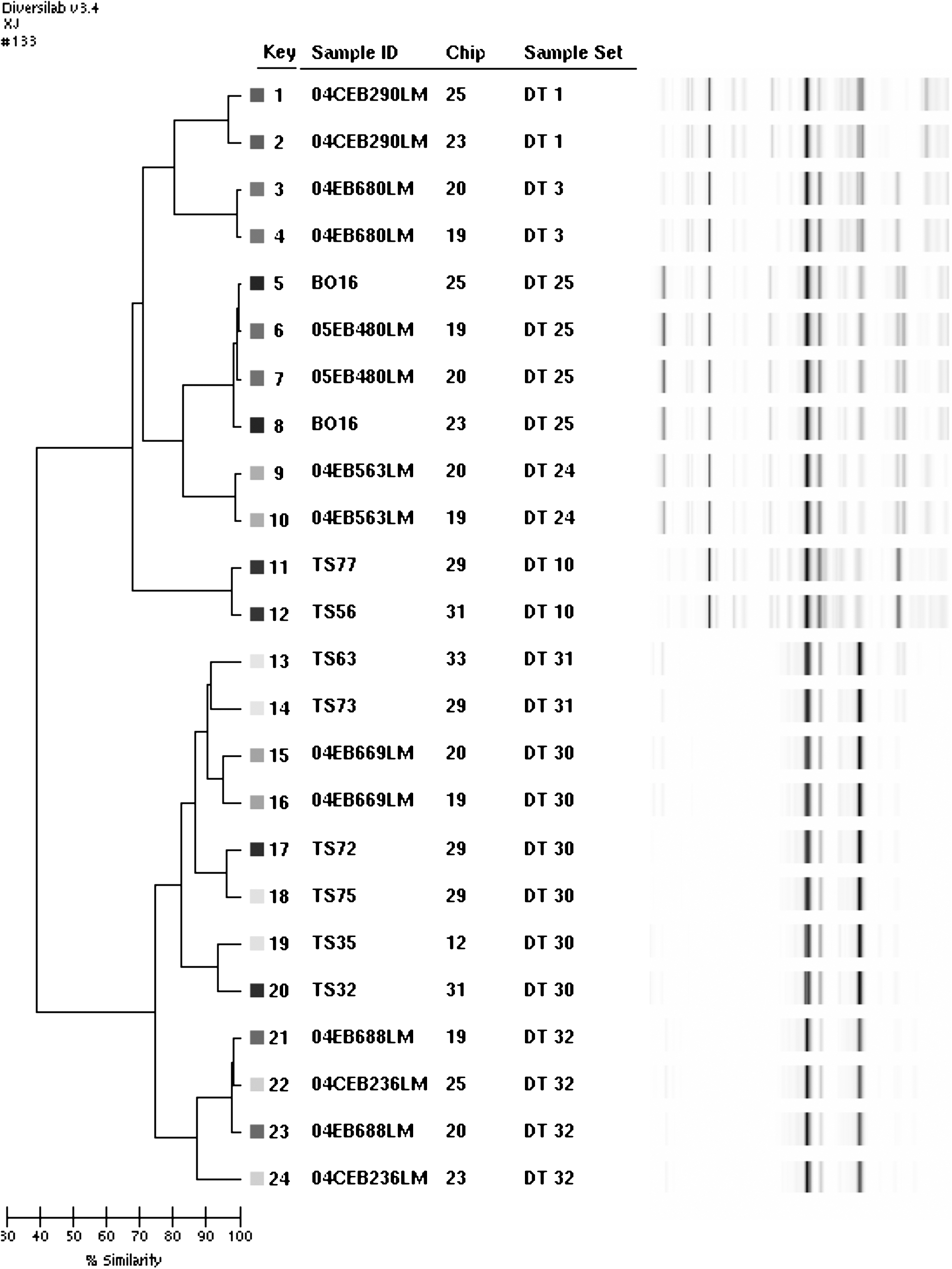
Repetitive-sequence-based polymerase chain reaction analysis with DiversiLab. Similarity dendrogram for 12 strains represented by two duplicates each and tested with two different chips: strain number, chip number, DiversiLab Type (DT) number, and virtual gel image. The four duplicate TS strains are TS32,TS72; TS56,TS77; TS35,TS75; and TS63,TS73.
Typing results on epidemiologically unrelated strains
Strains TS04, TS36, and TS56 showed the same DT (“10”) and ApaI/AscI PFGE type (“120/191”). Strains TS44 and TS30 displayed the same DT (“14”) and AscI type (“102”) but different ApaI types (“149” and “144”) (Table 1).
Five different DTs were encountered for six distinct epidemiologically groups: the same DT (DT 31) was observed for serotype 4b strains related to the epidemiologically groups “15”and “16.” Unique ApaI/AscI PFGE type was observed for each epidemiogical group (Table 1).
ID calculated on the panel of 105 unrelated isolates was 0.954, 0.988, 0.994, and 0.998 for DiversiLab, AscI-PFGE, ApaI-PFGE, and AscI/ApaI-PFGE, respectively. The DiversiLab ID was 0.958 and 0.695 for the serotype 1/2a isolates and for the serotype 4b isolates, respectively. The AscI-PFGE ID was 0.974 and 0.949 for the serotype 1/2a isolates and for the serotype 4b isolates, respectively. The ApaI-PFGE was 0.983 and 0.991 for the serotype 1/2a isolates and for the serotype 4b isolates, respectively. The ApaI/AscI-PFGE was 0.993 and 0.996 for the serotype 1/2a isolates and for the serotype 4b isolates, respectively.
Typing results on the total isolates panel
DiversiLab divided the 116 isolates into 40 DT (Fig. 2, Table 2). For 4b and 1/2b serotype isolates, the DiversiLab profiles were very closed. The DT assignments were systemically performed by comparing electrophoregrams and overlays.

Repetitive-sequence-based polymerase chain reaction analysis with DiversiLab. Similarity dendrogram for isolates representatives of each of the 40 DT: isolate serotype, DT number, isolate number in each DT, and virtual gel image.
The 40 DT were clustered into two groups. All the isolates of serotype 1/2a, 3a, 1/2c, 4a, and 4c were classified in a group, while all the isolates of serotypes 4b and 1/2b were in another group (Fig. 2). Out of 40 DT, 39 DT contained isolates of a same serotype (Table 2). Only one DT “35” contained isolates of two serotypes, 4b and 1/2b. The DT were composed of a variable number of isolates, between 1 and 19. The major DT were “25,” “30,” and “31” (Table 2).
The DT were composed of isolates of different food origins (Table 2). The 26 human strains (25 TS strains and 04 CEB 233LM) were divided into 11 different DT. Of these 11 DT, 9 included also food isolates (Table 2). All the 53 isolates of serotype 1/2a were clustered into 26 different DT (Tables 2 and 3). The eight isolates from serotype 1/2c were clustered into the same DT (DT 25) (Tables 2 and 3). All 38 serotype 4b isolates were clustered into seven different DT (Tables 2 and 3). The 14 isolates of serotype 1/2b were clustered into 5 DT (Tables 2 and 3).
All the isolates of serotype 1/2a, 3a, 1/2c, 4a, and 4c were classified in a group, while all the isolates of serotypes 4b and 1/2b were in another group, by ApaI/AscI PFGE (not shown). The two enzymes combination divided the isolates into 105 distinct profiles (Table 3). ApaI and combinated PFGE types contained isolates of a same serotype (Table 2). All AscI-PFGE types contained isolates of a same serotype except for type “46,” which contained one serotype 1/2a isolate and one serotype 3a isolate and type “62,” which contained one serotype 1/2b isolate and one serotype 4b isolate (Table 2). The 26 human strains were divided into 19 different combinated PFGE types. The combinated PFGE types were shared by isolates of different food origin. The AscI PFGE type “38,” “172,” “3,” “46,” and “22” were divided into 4 DT (“19,” “1,” “22,” and “26”), 3 DT (“1,” “4,” and “2”), 2 DT (“18” and “23”), 2 DT (“29” and “1”), and 2 DT (“3” and “24”), respectively. The ApaI PFGE type “12,” “149,” and “88” were divided into 2 DT (“23” and “9”), 2 DT (“12” and “14”), and 2 DT (“1” and “4”), respectively. The ApaI/AscI PFGE type “12/1” was divided into 2 DTs (“23” and “9”) (Table 2).
Discussion
PFGE is currently the gold standard for molecular subtyping of L. monocytogenes strains (Graves and Swamminathan, 2001). The objective of this investigation was to evaluate the potential of a new semi-automated rep-PCR technology, DiversiLab, for subtyping L. monocytogenes isolates of various origins, in comparison to PFGE. To our knowledge, these two methods have not been directly compared up to now.
The 116 tested isolates were divided by DiversiLab and PFGE into two clearly distinguishable groups: the first group contained serotypes 1/2a, 1/2c, and 3a, and the second contained serotypes 4b and 1/2b. This division correlated with the flagellar (H) antigen type and confirmed the phylogenetic divergence between strains of serotype 4b and 1/2b and those of serotype 1/2a and 1/2c, demonstrated by other typing results (Vines et al., 1992; Brosch et al., 1994; Harvey and Gilmour, 1994; Kathariou, 2002; Ragon et al., 2008).
The DiversiLab ID obtained for serotype 1/2a strains was 0.958. This result suggested that DiversiLab could provide an alternative tool for analyzing genetic relatedness among the 1/2a strains, a prevalent serotype in food and food-processing environments. The DiversiLab ID obtained for serotype 4b strains was low (0.695). However, this investigation demonstrated the ability of the DiversiLab System to differentiate strains of serotypes 4b and 1/2b, in contrast to results obtained by Van Kessel et al. (2005) in which all the 1/2b and 4b strains were clustered in the same DT. The relatedness between strain patterns obtained by Van Kessel et al. (2005) was calculated with the Pearson correlation coefficient instead of the Kullback–Leibler method used here. With DiversiLab, the eight strains of serotype 1/2c were genetically indistinguishable. When using PFGE with AscI or ApaI, these 1/2c strains differed in no more than two bands. These results confirmed the close relatedness within strains of this serotype, which had already been demonstrated by previous investigations using PFGE (Buchrieser et al., 1991; Carrière et al., 1991), ribotyping (Jacquet et al., 1995), or random amplified DNA PCR (RADP) (Niederhauser et al., 1994). However, the present investigation included a low number of strains of this serotype. It would be useful to analyze a larger number of 1/2c strains to validate this result.
All DT and combinated PFGE types were serotype specific except for one DT in which one strain of each serotype 4b and 1/2b was clustered and one AscI-PFGE type in which one strain of each serotype 1/2a and 3a was clustered. Lukinmaa et al. (2003) and Brosch et al. (1994) also obtained similar PFGE profiles for some strains of serotype 1/2a and 3a. These two serotypes were known to belong to the same flagellar H antigen group (AB) and have since been reported to belong the same genetic subgroup (Brosch et al., 1994; Nadon et al., 2001). The DT and PFGE types obtained here appeared not to be correlated with origin of strains; it would be useful to analyze a larger number of isolates to validate this result. However, these data were in accordance with data obtained by rep-PCR on 128 isolates from seafood (Chou and Wang, 2006) and by PFGE on 35 human and food strains (Buchrieser et al., 1991). On the contrary, the four rep-PCR groups obtained by Jersek et al. (1999) were correlated with the origin of isolates (human, food, and animal), but not with the serotypes as each of the 4 groups contained 64 isolates of various serotypes.
The epidemiologically unrelated strains TS04, TS36, and TS56 shared same types with both DiversiLab and PFGE. These three strains have been already tested by AscI and ApaI PFGE during the WHO multicenter international typing study, by four different laboratories (Brosch et al., 1996). For AscI PFGE, results obtained by the four laboratories confirmed our data. For ApaI, the same PFGE type was obtained by three laboratories. For one laboratory, a same type was obtained for the two strains, TS56 and TS36, different from the strain TS04. Previous data obtained during the WHO multicenter international typing study have shown that these three strains shared concordant RADP and ribotyping patterns. All these results suggested that the three strains may have been related. The epidemiologically unrelated strains TS44 and TS30 displayed the same DT and AscI type and a different ApaI type. For AscI PFGE, results obtained by the four laboratories during the WHO study confirmed our data. For ApaI, just one laboratory obtained two different ApaI PFGE types (Brosch et al., 1996).
The PFGE results showed a higher discriminatory ability with ApaI-PFGE than with AscI-PFGE, in agreement with previous results (Brosch et al., 1994; Chou and Wang, 2006). However, Harvey et al. (2004) obtained a lower discriminatory power with ApaI than AscI on a set of 45 food isolates. The results demonstrated the higher discriminative power of PFGE when compared with DiversiLab. Our results were in agreement with those of Harvey et al. (2004), who reported an ID value of 0.941 for a nonautomated rep-PCR, when compared with ApaI-PFGE (0.970) and AscI-PFGE (0.977). On the contrary, Chou and Wang (2006) have demonstrated that ApaI-PFGE and (nonautomated) rep-PCR had a similar discriminatory ability to differentiate isolates from catfish and non-catfish seafood. However, in this investigation, for serotype 1/2a strains, six different AscI-PFGE types and three ApaI-PFGE types were divided into different DTs. In this case, DiversiLab appeared to be more discriminatory than PFGE.
PFGE is a time-consuming and labor-intensive method: the analyses are completed in 30 hours to 3 days from the time a pure culture of the bacteria has been obtained. It also requires highly skilled operators and does not offer standardized reagents. In contrast, DiversiLab is a labor-saving and rapid method. As this semi-automated system includes a 1-day turnaround time and a large number of isolates can be analyzed rapidly. The method is easy to perform and does not require ethidium bromide. The amplicons are separated on microfluidic chips with internal standards added to each sample well; it thus avoids the need to compare separate gels. Standardized reagents are commercially available in kit form. Moreover, Healy et al. (2005) found that this system was robust for varying experimental conditions (operators, instruments, DNA concentrations, and laboratories). Web-based software provides standardized comparisons among isolates. Moreover, this software provides unlimited user access for any computer with an Internet connection.
Conclusions
DiversiLab allowed a good discrimination between serotype 1/2a strains, mostly encountered in food and food-processing environments. DiversiLab was less labor intensive than PFGE and provided results in <24 hours as compared to from 30 hours to 3 days for PFGE. The DiversiLab System could be useful for reducing a serotyping load during an outbreak. This system could be a useful tool during the first stages of an epidemiological survey, in addition to PFGE. In long-term epidemiological studies where less discrimination is needed, methods such as DiversiLab could be very promising tools.
Footnotes
Acknowledgments
This work was conducted in the frame of activities of the Community Reference Laboratory for L. monocytogenes and was supported by a grant from the Directorate-General Heath and Consumer Protection (DG Sanco) of the European Commission.
Disclosure Statement
No competing financial interests exist.