
Abstract
Cryptosporidium is a parasitic protozoan of great interest because of its widespread occurrence in surface waters, high degree of infectivity, and difficulty of risk management associated with its presence and control. Information about environmental loading and seasonal prevalence of Cryptosporidium oocysts is important for development of watershed management plans to protect public health. Healthy adult beef cattle are known to shed oocysts into the environment, and Cryptosporidium oocysts are often present all year in streams and groundwater in livestock agriculture areas. Surface soil and forage samples from 12 Virginia, United States, paddocks were analyzed bimonthly over 3 years for the presence and concentrations of Cryptosporidium oocysts. Half of the paddocks were grazed by stocker beef from November to September. The other half were managed for hay, but were grazed for a few days by the same animals in late fall and early spring to clean up late fall forage regrowth. Annual mean oocyst prevalences in soil were 57.9% and 48.4% in pasture and hay paddocks, respectively. Mean annual oocyst prevalences on forage were 52.4% and 40.5% in pasture and hay paddocks, respectively. Prevalence and concentration of oocysts on hay forage was highest in summer. Oocyst concentrations increased with increasing prevalences in both management systems. Wild animals appeared to be efficient vectors for oocyst distribution among paddocks. Canopy management, short-cycle rotational grazing, and control of wildlife are potential strategies for reduction of Cryptosporidium oocysts in pasture and lessening risk of contamination of water supplies, but further studies are needed before recommendations can be made.
Introduction
C
Transport of C. parvum oocysts to water sources is not completely understood. Agricultural sources of oocysts can take three different routes to water supplies. Oocysts are sometimes deposited directly in water or near-stream source areas by deposition of feces by grazing animals or application of manure for soil improvement (Hubbard et al., 2004; Schijven et al., 2004). Overland runoff transports oocysts to water bodies (Davies et al., 2004) and oocysts are transported by infiltrating water (Mawdsley et al., 1995), especially in macroporous soils (Harter et al., 2008; Boyer et al., 2009).
Information about total number of oocysts deposited on soil and plant surfaces (environmental loading) and seasonal prevalence of oocysts is important for development of watershed management plans to protect public health (Atwill et al., 2006). Calves are usually considered to be the primary source of oocysts (up to 107 oocysts/g of manure) in beef production (Atwill et al., 1999), but apparently healthy adult beef cattle have been shown to shed oocysts in concentrations of 25 to 1.8 × 104 oocysts/g of manure (Scott et al., 1994). Studies have found C. parvum oocysts present all year in streams flowing through livestock agriculture areas, but highest frequency of oocyst occurrence coincides with calving season (Kemp et al., 1995; Ong et al., 1996; States et al., 1997; Bodley-Tickell et al., 2002). Boyer and Kuczynska (2003) found oocyst concentrations in a karst spring to be greatest in late fall to early winter even though calving season was in the early spring. They concluded that oocysts were transported to epikarst where oocysts accumulated until saturated hydrologic conditions flushed oocysts into karst conduits and subsequently to springs.
On-farm strategies to reduce foodborne pathogens depend on an understanding of a myriad of complex interactions (Callaway and Oliver, 2009). Development of grazing strategies and pasture management systems that minimize opportunities for C. parvum oocysts to reach drinking water sources requires basic knowledge about temporal and spatial distributions of oocyst deposition densities on landscapes. Armed with that basic knowledge, pasture and livestock managers have the opportunity to develop management systems that reduce environmental loading of oocysts, increase the opportunity for environmental conditions to deactivate oocysts, and keep oocysts out of runoff source areas. The purpose of this study was to investigate the temporal distribution of oocysts at the soil surface and on standing forage in rotationally grazed beef pastures.
Materials and Methods
The study was conducted on a site consisting of Angus-cross stocker cattle grazing tall fescue (Schedonorus phoenix) pasture and fed fescue hay as needed. The pasture was located at Virginia Tech's Shenandoah Valley Agricultural Research and Extension Center, Steele's Tavern, VA (37°55.77′ N, 79°13.27′ W; 540–552 m above sea level) (Fig. 1). Runoff from the road and hill above the road were diverted away from the study site by roadside ditches. Study weather data were obtained from the U.S. Weather Bureau station at Lexington, VA, located about 25 km south of the study site. Soils at the study site are primarily Frederick (fine, mixed, semiactive, mesic Typic Paleudults) and Carbo (very-fine, mixed, active, mesic Typic Hapludalfs) silt loams. The study site is located on karstified carbonates consisting of Ordovician limestone and dolostone.
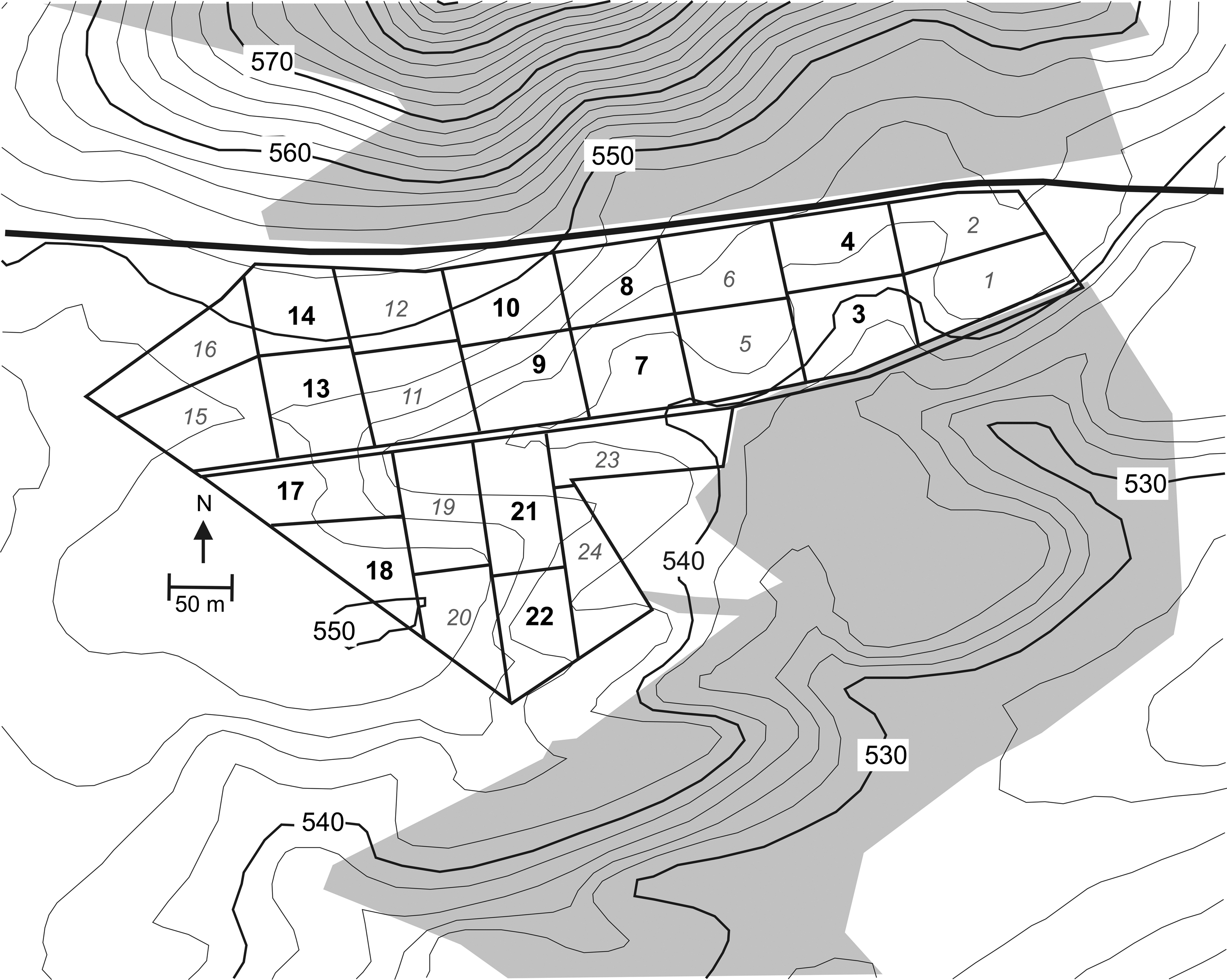
Study site at Steele's Tavern, VA. Paddocks used in this study are numbered in bold lettering. Shading represents forest, and elevation contour interval equals 2 m.
The 14 ha pasture was divided into 24 paddocks (∼0.6 ha each) that were rotationally grazed by stocker beef cattle. Twelve of the paddocks were used for this study (Fig. 1). Poultry litter was used as a soil amendment in the 12 excluded paddocks. The 12 selected paddocks underwent typical management for the region. Sample sizes were determined by availability of laboratory and personnel resources. Three samples per paddock on each visit were considered representative. Even-numbered paddocks (hay paddocks) were used to grow hay for stockpile feeding cattle in the odd-numbered paddocks (pasture paddocks) during winter. Hay paddocks were fertilized in fall and hayed in summer; cattle were rotated in for cleanup grazing of fall forage regrowth before winter rest. Pasture paddocks were stocked with four 180 kg Angus-cross steers each in November. Steers were rotationally grazed in the pasture paddocks until the cattle were sold at a weight of about 400 kg in August or September of the following year and new stocker steers were purchased in November from Midwestern U.S. suppliers. High-energy supplemental feeds for the cattle included corn and broiler litter.
The paddocks were visited bimonthly from May 2002 to November 2004, and the top 2.5 cm of soil beneath the thatch was sampled with a 5-cm-diameter soil probe at three separate arbitrary locations in each paddock where vegetation cover was present. Samples were not obtained from the same points during subsequent visits. A bimonthly sampling strategy was employed because of the long travel distance and resource limitations. Soil samples were not taken in proximity (∼1 m) to manure paddies or in low spots such as hoof prints. The three soil samples from each paddock were mixed by hand in sterile plastic bags and transported back to the laboratory for analyses of Cryptosporidium oocyst concentrations. Forage samples were also taken at each soil sample location, and combined and mixed by hand in sterile plastic bags in each paddock on seven of the sample months (May, July, December 2002; March, September, November 2003; May 2004) for Cryptosporidium oocyst analyses.
Soil (25 g) and grass (4 g) samples were processed using the NaCl flotation method as previously described (Kuczynska and Shelton, 1999). Three 10 μL aliquots (of the final 100 μL suspension) were pipetted onto slide wells (5 mm diameter), dried on a slide warmer, and stained using a commercial immunofluorescence antibody kit (Merifluor; Meridian Diagnostic, Cincinnati, OH). Numbers of oocysts per soil or grass sample were obtained by multiplying the average number of oocysts in three wells by 10. Microscope slides were examined with an epifluorescence microscope (Zeiss; Carl Zeiss MicroImaging, Thornwood, NY) at 250 × magnification. No speciation or test for infectivity was done on the oocysts since we were only looking at oocysts as indicators of Cryptosporidium.
Oocyst concentrations were converted to number of oocysts per gram of soil or grass, and oocyst concentration statistics were calculated from all oocyst-positive samples. Statistical analyses were performed with SAS Version 9.1 (SAS Institute, Cary, NC). The LOGISTIC procedure was used to test for season or treatment effects on the presence or absence of oocysts. The presence/absence of oocysts was treated as a binary variable, where presence was coded as 1 and absence as 0. Deviance Chi-square (called −2 Log L in SAS) was used to test the goodness of fit of the logistic model. Sign tests using paired samples tested for differences in oocyst prevalences and concentrations. The paired samples were paddock or seasonal means. Repeated measures were not considered since sign tests were performed on means. Linear and curvilinear regression were used to test for relationships between mean oocyst prevalences and mean oocyst concentrations. Statistical tests are significant at p ≤ 0.1 level unless stated otherwise.
Results
Figure 2 shows observed and normal total precipitation and mean daily temperature, by months, for the study years. Mean daily temperature was nearly normal all 3 years with the exception of cooler than normal temperatures during the 2003 growing season. The cooler than normal temperatures in 2003 were associated with a wetter than normal spring and summer.

Monthly and normal climatic conditions at Lexington, VA, for the years of study: (
Table 1 summarizes mean bimonthly and annual prevalences of oocysts in surface soils and on forage in pasture and hay paddocks. Table 1 also shows bimonthly and annual mean concentrations of oocysts per gram of soil or forage in samples that were positive for Cryptosporidium oocysts. Bimonthly prevalences of oocysts in pasture or hay soils were not significantly different. Bimonthly prevalences of oocysts on forage were significantly different in pasture and hay paddocks. Bimonthly prevalences of oocysts on forage were lowest in winter to early spring and highest in summer to early fall.
Statistically significant differences in seasonal means are indicated by italicized numbers in a column. Differences between pasture and hay are indicated by asterisks at the top of the columns (i.e., two columns with the same number of asterisks are significantly different).
n.d., not determined.
Mean bimonthly concentrations of oocysts in surface soil samples that tested positive for oocysts ranged from 0.17 to 0.27 oocysts/g in the pasture paddocks and 0.16 to 0.22 oocysts/g in the hay paddocks. Mean bimonthly concentrations of oocysts in soil were not significantly different in the pasture or the hay paddocks. Mean bimonthly concentrations of oocysts on forage samples that tested positive for oocysts were significantly different in the hay paddocks, but not in the pasture paddocks. Mean bimonthly concentrations of oocysts on forage in the hay paddocks ranged from 0.81 oocyst/g in May and November to 1.56 oocysts/g in September. Although mean bimonthly oocyst concentrations on forage in pasture were not significantly different, they ranged from 0.81 to 2.31 oocysts/g.
In the 16 months sampled, prevalence of oocysts was greater in the pasture paddock soils than in the hay paddock soils eight times. Prevalence of oocysts in soil was equal in both paddock treatments in three of the sample months. No significant difference between bi-monthly prevalences of oocysts in pasture versus hay paddock soils was observed.
Oocyst concentrations in pasture paddock soils were greater than oocyst concentrations in hay paddock soils when considering bi-monthly means. Most of the difference occurred in summer and fall when oocyst concentrations were 0.06 to 0.09 oocysts/g of soil greater in the pasture paddocks (Table 1). Mean oocyst concentrations on the forages also differed. Oocyst concentrations on pasture paddock forages were about twice the concentration (2.31 vs. 1.13 oocysts/g) of oocysts on hay paddock forages in July.
Seasonal shifts in oocyst prevalence and concentration on pasture forage suggest that there might be a relationship with air temperature. Figure 3 shows trends between mean monthly air temperature and mean prevalences and mean concentrations of oocysts on the pasture forage. Although the bimonthly concentrations of oocysts on pasture forage were not significantly different, the concentrations did follow the temperature pattern. Warmer air temperatures might change oocyst and vegetation surface properties, thereby enhancing the ability of oocysts to cling to vegetation.

Mean oocyst prevalences (open symbols) and mean oocyst concentrations (filled symbols) versus mean monthly temperature on pasture (squares) and hay (triangles) paddock forages.
Direct relationships between oocyst prevalence and concentration have been observed on an English lowland farm (Sturdee et al., 2003) and in an upland watershed in England (Sturdee et al., 2007). A linear relationship was observed for oocyst concentration versus prevalence on pasture forage in this study (Fig. 4). Mean oocyst concentrations on pasture forage increased about 0.32 oocysts/g for each 10% increase in prevalence. An increase in oocyst concentration with increased oocyst prevalence in pasture soil was also observed with mean concentration increasing about 0.035 oocysts/g for each 10% increase in prevalence (Fig. 5). All of the lines in Figures 4 and 5 were forced through the origin by using regression analysis with no intercept, because the concentration of oocysts has to be zero if prevalence of oocysts is zero. Obviously, if an intercept term was included in the regression analyses, the R-squares would be lower. Curvilinear line fitting did not improve predictability associated with the relationships.

Mean oocyst concentrations versus mean oocyst prevalences on pasture paddock (filled circles) and hay paddock (open triangles) forages. Regression lines represent trends for pasture (solid line) and hay (dashed line). Regression analyses were run with the no intercept option.
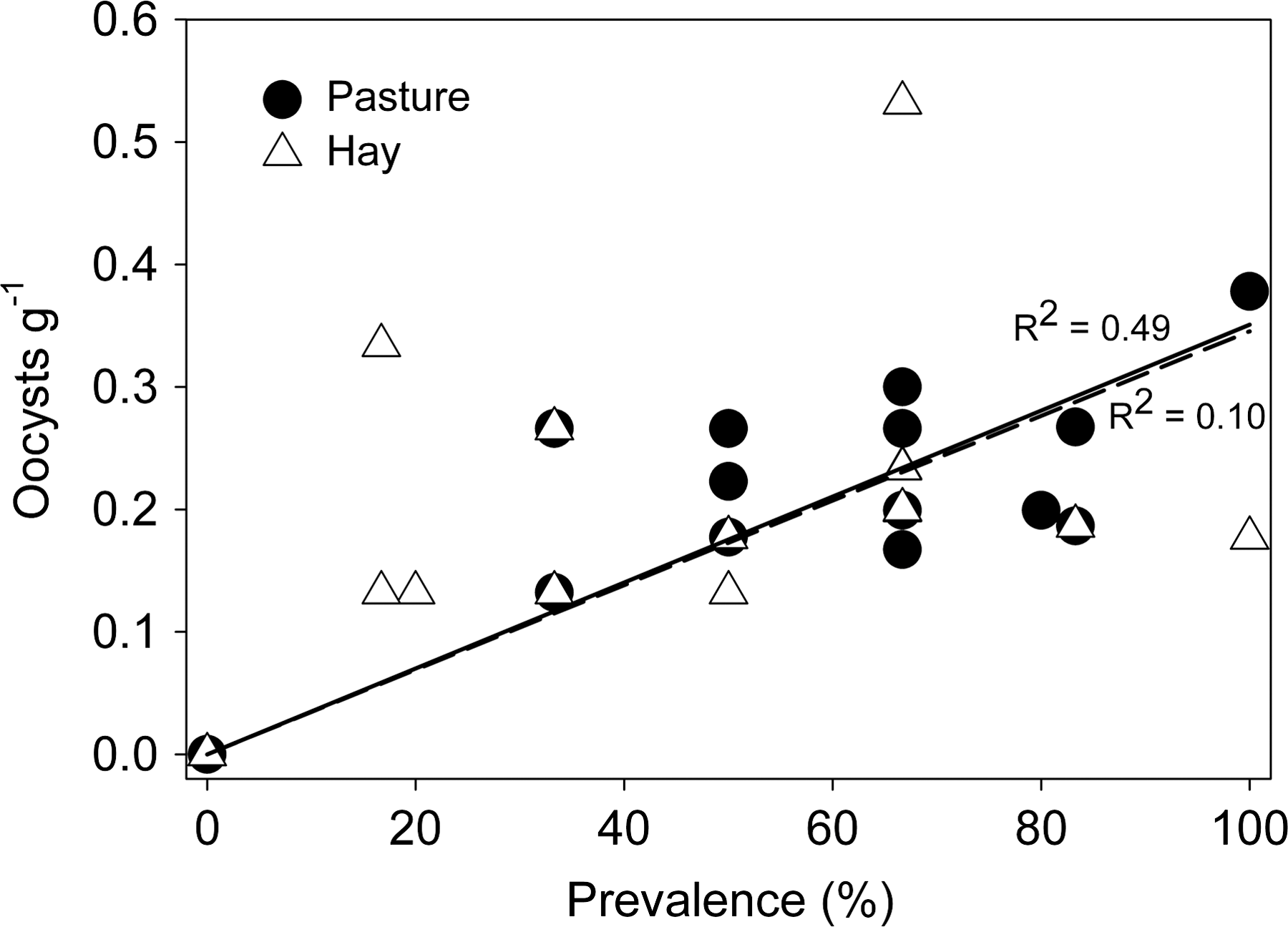
Mean oocyst concentrations versus mean oocyst prevalences in pasture paddock (filled circles) and hay paddock (open triangles) soils. Regression lines represent trends for pasture (solid line) and hay (dashed line). The slope of the regression line for hay is not statistically different from zero. Regression analyses were run with the no intercept option.
No significant differences were found in the annual prevalences of oocysts. There were significant differences in the annual mean concentrations of oocysts. Annual mean concentrations of oocysts were 0.04 oocysts/g greater in pasture paddock soils than hay paddock soils. Annual mean concentrations were 0.44 oocysts/g greater on pasture paddock forages than hay paddock forages.
Discussion
Pasture paddocks
Uniformity of oocyst prevalences and concentrations throughout the year in pasture soils suggests that oocysts were being deposited year round. Further, oocyst deposition rates had to be equal to rates of oocyst removal to maintain the uniformity in prevalences and concentrations of oocysts in the surface soil. Oocysts could have been removed from pasture soils by surface runoff, subsurface transport with infiltrating water, and die-off with subsequent microbial decomposition. Studies have shown that rainsplash can relocate fecal bacteria (Boyer and Belesky, 2009) and fungal spores (Paul et al., 2002) from soil forage and grass surfaces. A similar mechanism of oocyst removal from pasture soils would be expected.
Prevalence and concentration of oocysts were expected to be highest in late fall when younger stocker cattle were present. Cryptosporidium infection is known to be more prevalent in young calves than in adult cattle. Infections of adult cattle by Cryptosporidium are known to occur, but with excretion of lower concentrations of oocysts (Fayer et al., 2000). The greater quantity of manure deposition by adult cattle than young stocker cattle could account for a uniform environmental loading of oocysts throughout the year (Kuczynska et al., 2005). Barwick et al. (2003b) found that 17% of soil samples from New York dairy farms tested positive for Cryptosporidium and that 92% of farms had at least one soil sample test positive for Cryptosporidium. The high farm prevalence might have been a result of accumulation of feces in soil over time (Barwick et al., 2003a). Oocysts were more likely to be found in soil of land areas that accumulated fresh manure over time (Barwick et al., 2003b).
Bimonthly oocyst prevalences on the pasture forage changed systematically with the lowest prevalence in winter and the highest in summer. Oocyst concentrations on pasture forage did not show significant bimonthly changes even though mean concentrations followed a pattern similar to bimonthly prevalences. In summer, cattle roam throughout paddocks in search of food and would be expected to excrete manure more evenly over paddocks as opposed to winter, when most of the feeding is done with supplemental hay. By concentrating cattle near hay feeders, cattle do not roam the paddocks in search of food, and manure deposition is also concentrated near hay feeding stations.
Hay paddocks
Similar to the pasture paddocks, bimonthly oocyst prevalences and oocyst concentrations in hay paddock soils did not change significantly across the year. That was surprising since cattle were rarely present in the hay paddocks. Cattle were generally present in the hay paddocks in early spring before significant new grass growth occurred and late summer following the last hay harvest and before the fall sale of cattle. Mean oocyst prevalence in September was highest observed mean (66.7%), but it was not significantly different from those in other months (Table 1).
Bimonthly oocyst prevalences on the hay paddock forage followed a clear trend with highest prevalences and concentrations in late summer and early autumn and lowest in winter (Table 1). The trend was expected given that cattle were present in hay paddocks at times of highest oocyst prevalences and concentrations. In the period from the time cattle were sold (September) to the time just before new stocker cattle were introduced (November), the oocyst concentrations on forage dropped nearly 50% from 1.56 to 0.81 oocysts/g of forage. At the same time, prevalence of oocysts on the hay paddock forages dropped about 37%.
Mean oocyst prevalences and concentrations on hay paddock forages appeared to be related to mean monthly temperature (Fig. 3), indicating that there was some seasonality in the data. Mean oocyst concentrations on hay paddock forages were likely to increase with increased prevalence (Fig. 4). The regression line of oocyst concentration versus oocyst prevalence on hay paddock forages was significant and showed that concentration increased 0.19 oocysts/g of forage for each 10% increase in prevalence. The regression relationship between oocyst concentration and prevalence in hay paddock soils was nearly identical to the relationship observed in pasture paddock soils (Fig. 5), but not statistically significant.
Pasture versus hay
Greater oocyst prevalences and concentrations in pasture soils and on pasture forages than in hay soils and on hay forages were expected if cattle were excreting oocysts in their manure. Others have found that adult cattle excrete oocysts in moderated concentrations relative to concentrations excreted by infected calves (Sturdee et al., 2003). Infected adult cattle rarely exhibit clinical symptoms of cryptosporidiosis. Cattle were not tested for Cryptosporidium infection in this study. Cattle spent much more time throughout the year in the pasture paddocks than in the hay paddocks, thus excreting more manure in the pasture paddocks.
Relatively high oocyst prevalences and concentrations in the hay paddocks were surprising since cattle were rarely present there. Wild animals might have been one source of oocysts throughout the year. Effects of wild animals on the prevalence and concentrations of Cryptosporidium oocysts in the soils and on the forage are unknown. Meadow voles (Microtus pennsylvanicus), deer mice (Peromyscus maniculatus), raccoons (Procyon lotor), and white-tailed deer (Odocoileus virginianus) are common in the area and all are known sources of Cryptospridium spp. oocysts (Feng et al., 2007). Wild animals had unrestricted access to all paddocks and could have easily transported oocysts between paddocks. Sturdee et al. (2003) suggested that wild animals chronically infected with Cryptosporidium could serve as a perpetual source of infection for domestic livestock.
Regardless of effects of wild animals, higher oocyst prevalences and concentrations in pasture indicate that cattle were a source of oocysts. However, oocyst concentrations in cattle feces in this study were not assessed. Other studies have found that cattle feces are a source of oocysts (Scott et al., 1994; Atwill et al., 1999, 2006). The cycle of oocyst ingestion, infection, and oocyst excretion is exacerbated in pasture by higher prevalences and concentrations. Further, as the chance of ingestion (prevalence on forage) increases, numbers of oocysts ingested increases because of higher oocyst concentrations. The expected number of oocysts ingested per day can be estimated by the equation
where E(O i) = expected number of oocysts ingested per day, P = proportion of positive samples (prevalence), O c = mean number of oocysts/g of forage in samples that were positive for oocysts, and F = grams of forage ingested per day. If a 300 kg steer is considered to eat 3% of its weight in forage dry matter (9000 g)/day and the forage is 15% dry matter, then F equals 60,000 g/day. Substitution of the annual mean oocyst prevalences and concentrations (pasture: 0.52 and 1.44; hay: 0.41 and 1.00, respectively) into Equation 1 gives an expected mean daily oocyst ingestion per steer of 44,928 in the pasture paddock and 24600 in the hay paddock.
Expected mean daily ingestion of oocysts can be estimated for prevalences using the linear trends shown in Figure 4. Substitution of prevalences from 0% to 70% for P and the associated oocyst concentration for O c in Equation 1 gives the E(O i) as shown in Figure 6. Higher concentrations of oocysts on pasture paddock forage versus hay paddock forage causes the differences in expected mean daily ingestion between pasture and hay to increase with increasing prevalence. This exercise is only illustrative of potential differences between the two paddock managements. A great deal of uncertainty is associated with the relationship. First, the scatter around the lines in Figure 4, especially in the pasture paddocks, lends some uncertainty in the input values even though the slopes of both lines were significant. Additionally, Equation 1 is dependent on amount of forage ingested, which is determined by several factors, including animal size, forage dry matter content, and nutritional value of the forage. It is also important to consider infectivity of oocysts. Oocysts are expected to lose infectivity over time, but such factors as temperature, desiccation, and ultraviolet radiation have all been shown to affect longevity of oocyst infectivity (Robertson et al. 1992; Olson et al., 1999; Brookes et al., 2004; Li et al., 2005). Walker et al. (1998) suggested that vegetated soil protects oocysts from effects of temperature extremes and desiccation. Nevertheless, the data show that oocyst ingestion will be greater in pasture paddocks than in hay paddocks since prevalences and concentrations on the pasture forage is greater.

Expected mean daily number of oocysts ingested with forage versus prevalence of oocysts on forage in pasture and hay paddocks. Calculations based on a 300 kg steer ingesting 60,000 g of forage per day.
Water quality management challenges
Cryptosporidium is ubiquitous in surface water (Rose, 1997) and has been found in high concentrations in an Appalachian karst spring draining grazing lands (Boyer and Kuczynska, 2003). Samples drawn from a well in a shallow karst aquifer on this research site were found to contain Cryptosporidium oocysts (unpublished data). Protection from waterborne disease is the ultimate goal of landscape and water quality management for control of opportunities for Cryptosporidium oocysts to enter drinking water supplies.
Prevention of infection of livestock might be an effective means to prevent oocysts from reaching water supplies. Wild animals appear to be a source of environmental loading of oocysts on pasture. Sturdee et al. (2003) recommended that maintenance of good animal health might reduce severity of infection of grazing livestock and subsequent environmental loading of oocysts. Once oocysts are in the environment, management decisions and practices for reduction of opportunities for oocysts to reach water supplies are needed.
Reduction of surface runoff and exclusion of grazing animals from runoff source areas will reduce opportunities for oocysts to be transported to surface streams. Vegetative filter strips have often been found effective in reducing runoff and pathogen transport (Trask et al., 2004). Deep infiltration of water might be counterproductive because oocysts have been found to survive for long times in temperate climate agricultural soils (Jenkins et al., 2002). Oocysts have been found to leach through macroporous soils (Darnault et al., 2004; Harter et al., 2008; Boyer et al., 2009) and remain infective for extended time (Boyer et al., 2009). Management practices that leave vegetation short enough to allow radiant energy to reach the soil surface might produce enough surface heating and allow enough penetration of ultraviolet radiation to inactivate oocysts. Trevisan et al. (2002) suggested that canopy management of hay meadows might be an effective means for controlling fecal bacteria. Assuming that most oocysts on forage are located on lower levels of the forage, livestock would ingest fewer oocysts from short forage if they are moved to other pasture with taller forage. Short rotations might be one way to reduce the number of oocysts ingested, thereby reducing infection and subsequent environmental contamination.
On the basis of the high prevalences and concentrations of oocysts in the infrequently, lightly grazed hay paddocks, it appears that wild animals are an important source of oocysts in grazing systems. However, there is the possibility that oocysts are just persisting and reinfected cattle are maintaining the supply of oocysts. Control of wild animals, especially small animals, would be difficult if not nearly impossible. Multidisciplinary studies that include wildlife experts are needed to devise management strategies that lessen the role of wild animals in the dissemination and perpetuation of existence of oocysts in grazing systems. Runoff from grazed paddocks might have transported some oocysts onto hay paddocks, but previous infiltration studies (unpublished data) showed that infiltration rates on the site were high and runoff was not expected to be a problem. The farm-to-fork concept for protecting food safety targets all aspects of food production with control measures starting at the farm and ultimately ending at the consumer's fork or drinking cup (Callaway and Oliver, 2009). Identification of the causes of foodborne and waterborne contamination is needed to effectively manage associated risks (Jacob and Powell, 2009). Control of Cryptosporidium oocysts entering the environment at the farm level provides opportunities for reducing treatment costs and reducing public safety risks associated with food and water supplies.
Conclusions
Cryptosporidium oocysts were ubiquitous in surface soils and on forage in grazed paddocks as well as, infrequently, lightly grazed paddocks used for hay production. Prevalences and concentrations of oocysts were generally greater in the soils and on the forage of the pasture paddocks than the hay paddocks. Lack of seasonality in the prevalences of Cryptosporidium oocysts in soil suggests that oocysts are persistent. Persistence of Cryptosporidium oocysts in the hay paddock soils suggested that wildlife might be an important source and transport vector of Cryptosporidium oocysts in the area studied. High prevalence and concentration of Cryptosporidium oocysts on forage might be a source of infection to grazing livestock ingesting contaminated forage. Canopy management, short-cycle rotational grazing, and control of wildlife are potential strategies for reduction of Cryptosporidium oocysts in pasture and lessening the risk of contamination of water supplies, but further studies are needed before recommendations can be made. This study was limited to one specific geographical location with a single geology. Further studies across other locations and geologies are needed before broad recommendations can be made for pasture management systems that reduce opportunities for Cryptosporidium oocysts to contaminate water supplies.
Footnotes
Acknowledgments
Contribution of the U.S. Department of Agriculture–Agricultural Research Service (USDA-ARS). Trade and company names are used for the benefit of readers and do not imply endorsement by the USDA. This research was supported by the 205 Rangeland, Pasture, and Forages National Program of the USDA-ARS. The authors gratefully acknowledge the technical assistance of Laura Cooper and Derek Hall. Virginia Tech and the Shenandoah Valley Agricultural Research and Extension Center are thanked for allowing us to sample in their research paddocks.
Disclosure Statement
No competing financial interests exist.