
Abstract
A total of 360 spatially and temporally related Campylobacter isolates, including 168 from clinical human cases (Campylobacter jejuni n = 148; Campylobacter coli n = 20) and 192 from retail meats (C. jejuni n = 114; C. coli n = 78), were analyzed for antimicrobial susceptibilities, virulence, and genotypic profiles. Ciprofloxacin-resistant C. jejuni was observed in 13.5% and 19% of the isolates from humans and retail chicken breasts, respectively. Antimicrobial resistance to ciprofloxacin and erythromycin was detected in C. coli isolates recovered from 29% and 16.6% of retail meats and 15% and 5% humans, respectively. Overall, virulence determinants were more prevalent in Campylobacter isolates recovered from retail meats than from humans. C. jejuni isolates from humans were significantly associated with the rakR, dnaJ, and pld genes, whereas C. coli isolates from retail meats were associated with the dnaJ, pld, and virB11 virulence genes. Genotyping of 262 C. jejuni isolates using pulsed-field gel electrophoresis revealed a total of 186 unique SmaI patterns, with 14% of patterns composed of isolates recovered from retail meats and ill humans. All unique groups with indistinguishable SmaI patterns were further analyzed by a second restriction enzyme (KpnI), which revealed limited overlap between isolates from different sources. Significant association between doxycycline-resistant C. jejuni strains recovered from humans and different virulence genes (e.g., cdtB) was identified at the statistical level but not at the genotypic level. In conclusion, significant differences observed in the distribution of antimicrobial resistance profiles, virulence determinants, and genotypic diversity among C. jejuni and C. coli isolates indicate that there are sources other than retail meats that may also contribute to human Campylobacter infections.
Introduction
The majority of Campylobacter infections are sporadic and self-limiting in nature, which makes it difficult to accurately determine the true incidence rate. Due to the self-limiting nature of campylobacteriosis, antimicrobials are usually not recommended for treatment except in severe cases where fluoroquinolones and macrolides are the treatment options of choice (Butzler, 2004; Yates et al., 2005). Frequency of resistance to these important classes of antimicrobials has reported to be on the rise in the United States and globally (Gupta et al., 2004; Rozynek et al., 2008; NARMS, 2007). Additionally, multidrug-resistant C. jejuni and C. coli have been reported from food animals and retail meats, including poultry and swine (Zhao et al., 2001; Datta et al., 2003; Ge et al., 2003; Gebreyes et al., 2005; Thakur and Gebreyes, 2005; Thakur et al., 2006; Mena et al., 2008; Pointon et al., 2008).
Several putative virulence factors have been identified in Campylobacter species that contribute to motility, intestinal adhesion, colonization, toxin production, and invasion. Adhesion of the pathogen to the intestinal epithelium is important for colonization and to increase the secretion of bacterial toxins. An important adhesion is the outer membrane protein CadF, which is conserved in both C. jejuni and C. coli and binds to fibronectin in the gut, thereby helping the pathogen in adhesion and invasion (Konkel et al., 2001; Müller et al., 2006). Motility is via flagella, which is coded by the flagellin gene ( flaA), and is important for the bacteria to reach the attachment sites in the intestine (Konkel et al., 2004). Heat shock proteins, including DnaJ, are associated with the thermal stress response and have been shown to play an important role in pathogenesis and colonization (Snelling et al., 2005). Other important genes that have been associated with Campylobacter pathogenesis include pldA (Zheng et al., 2006), the plasmid-encoded virB11 (Bacon et al., 2000), and racR (Datta et al., 2003). Cytolethal distending toxin production, encoded by the cdtA, cdtB, and cdtC genes, damages host enterocytes and facilitates penetration of the intestinal epithelium (Rozynek et al., 2005). Carriage of these genes is variable, and helps explain virulence differences among strains.
Recent studies have been conducted to determine the association of virulence genes and antimicrobial resistance in important bacterial pathogens such as Escherichia coli and Enterococcus faecalis (Bagger-Skjøt et al., 2007; McGowan-Spicer et al., 2008). Ciprofloxacin-resistant C. jejuni strains have been shown to have an enhanced in vivo fitness over isogenic susceptible C. jejuni strains in the chicken gut even in the absence of fluoroquinolone selection pressure (Luo et al., 2005), suggesting a linkage between resistance and colonization potential. Studies have also shown that humans infected with antimicrobial-resistant Campylobacter species have a longer duration of diarrhea when compared with antimicrobial-susceptible strains (Nelson et al., 2004; Helms et al., 2005). The major objective of this study, therefore, was to determine the prevalence and association of virulence genes and antimicrobial resistance profiles at the statistical and genotypic levels in different strain types of C. jejuni and C. coli isolated from retail meats and clinically ill humans.
Materials and Methods
Isolate source
Campylobacter isolates in this study were obtained from a baseline surveillance project conducted in Iowa from June 2001 to June 2002 (Hayes et al., 2003). A total of 814 retail meat samples were collected, consisting of 209 samples each of chicken breast, pork chop, and ground beef and another 187 ground turkey. Campylobacter was isolated using standard bacteriological methods and speciated by polymerase chain reaction (PCR) (Zhao et al., 2001; Thakur and Gebreyes, 2005). In addition, 148 C. jejuni and 20 C. coli isolates from clinically ill human patients collected in Iowa during the same period were included for comparison. A total of 262 C. jejuni and 98 C. coli were recovered for analysis.
Antimicrobial susceptibility testing
We used agar dilution to determine the minimal inhibitory concentrations (MIC) for four antimicrobials, following the Clinical and Laboratory Standards Institute protocol (CLSI, 2006), and C. jejuni ATCC 33560 as the quality control organism. Clinical and Laboratory Standards Institute interpretative criteria were used for ciprofloxacin (Cip, 4 μg/mL), erythromycin (Ery, 32 μg/mL), and doxycycline (Dox, 8 μg/mL) (CLSI, 2006). For gentamicin (Gen), the breakpoint level of 8 μg/mL was used following the National Antimicrobial Resistance Monitoring System criteria (NARMS, 2007). Multidrug resistance (MDR) in this study was defined as isolate exhibiting resistance to two or more antimicrobials at the same time.
Virulence gene characterization
Genomic DNA used for PCR was purified using the UltraClean Microbial DNA Kit following the manufacturer's instructions (MoBio Laboratories, Carlsbad, CA). The presence of virulence and toxin genes, including flaA, cadF, rakR, cdtA, cdtB, cdtC, dnaJ, pld, and virB11, were determined as described previously (Datta et al., 2003; Zheng et al., 2006). The primers were developed based on conserved sequences that were available in the GenBank database. A separate set of primers was developed for virulence genes where difference in the sequences were noticed between C. jejuni and C. coli. All PCR amplifications were performed in total reaction volumes of 50 μL consisting of 0.1 μg of template DNA, 0.25 mM concentration of deoxynucleotide triphosphate, 1.5 mM MgCl2, 50 pmol of each primer, and 0.2 U of AmpliTaq Gold (Perkin-Elmer, Foster City, CA). PCR cycling conditions included an initial denaturation of 94°C for 10 minutes followed by 30 cycles of 94°C for 30 seconds, annealing at primer-specific pair for 30 seconds, followed by extension at 72°C for 20 seconds. A final extension step was done at 74°C for 6 minutes. The final PCR products were run of 1.5% agarose gel mixed with ethidium bromide for analyzing the result.
Pulsed field gel electrophoresis
We used the U.S. Centers for Disease Control–standardized protocol for genotyping Campylobacter species by pulsed-field gel electrophoresis (PFGE) with slight modifications (Ribot et al., 2001). Fresh Campylobacter cells were suspended in phosphate-buffered saline (0.01 mol/L phosphate, pH 7.4; 0.85% NaCl) and adjusted to a reading of 0.42 ± 0.02 by Dade Microscan Turbidity Meter (Dade Behring, West Sacramento, CA). Next, genomic DNA embedded in agarose was digested with SmaI restriction enzyme (Boehringer Mannheim, Indianapolis, IA). The digested DNA was then separated by electrophoresis in 0.5 × TBE (Tris-Borate 15 EDTA) at 14°C and 6V/cm for 17 hours using a CHEF DR-III apparatus (Bio-Rad Laboratories, Hercules, CA) with pulse times of 6.7–38.4 seconds. Salmonella enterica serovar Braenderup H9812 was used as the reference marker. Gels were stained with ethidium bromide for 30 minutes and photographed under UV transillumination (Bio-Rad Laboratories). Campylobacter isolates showing identical patterns were further digested with a second restriction enzyme KpnI (Boehringer Mannheim). This was done to determine whether the use of a second restriction enzyme could further discriminate between the clusters created by the SmaI restriction enzyme.
Statistical and data analysis
The frequency of antimicrobial resistance patterns and MIC levels comparison between Campylobacter isolates from different sources were compared using the χ 2-test (Minitab, PA) and Fisher's exact two-tailed test when applicable. Significant association of antimicrobial resistance and virulence gene profile for the two Campylobacter species and isolation source was determined using the odds ratio test (Egret version 2.0.3; Cytel, Cambridge, MA). Results with a Type I error of p ≤ 0.05 was considered statistically significant. Analysis of PFGE data was performed using Bionumerics 4.0 software (Applied Maths, Kortrijik, Belgium) after normalization of the three Salmonella control lanes on each gel. The banding patterns were compared using the Dice coefficient and unweighted pair group method using arithmetic averages with 0.45% optimization and 0.85 position tolerance for SmaI and 0.41% optimization and 1.5% position tolerance for KpnI respectively. Isolate relatedness was determined using the unweighted pair group method using arithmetic averages.
Results
Antimicrobial resistance phenotypes
Antimicrobial resistance and the distribution of MIC50, MIC90, and MIC ranges to ciprofloxacin, doxycycline, erythromycin, and gentamicin for the two Campylobacter species and their sources are shown in Table 1. Antimicrobial resistance to ciprofloxacin (CipR) was observed in 13.5% and 19% of the C. jejuni isolates from humans and chicken breasts, respectively. None of the C. jejuni isolates were EryR. However, 14.2% of the C. coli isolates, including those isolated from humans (n = 1) and retail meats (n = 13), displayed EryR. CipR C. jejuni from chicken breast had a higher MIC90 (16 μg/mL) than those recovered from humans (0.25 μg/mL). The Ery MIC90 distribution in C. coli from retail meats was >64 μg/mL compared to 8 μg/mL from humans. Overall, the MIC50 and the MIC90 values to both Ery and Cip for C. coli were consistently higher compared with C. jejuni isolates across all sources. Within the retail meat types, significantly more C. coli isolates obtained from ground turkey displayed resistance to Cip, Ery, and Dox than isolates recovered from chicken breast (p < 0.01). All tested Campylobacter isolates demonstrated susceptibility to gentamicin. We found a significantly higher frequency of pan-susceptible C. jejuni than C. coli from chicken breast (p < 0.001).
MIC50 and MIC90 indicate the concentration (μg/mL) at which 50% and 90% of the isolates tested were susceptible to the antimicrobial, respectively.
Indicates the MIC level for Campylobacter isolates at a particular antimicrobial concentration.
Antimicrobials and associated resistance breakpoints (μg/mL).
Represents the number of Campylobacter isolates tested from different sources—C. jejuni: humans (n = 148), chicken breast (n = 110), and ground turkey (n = 4); C. coli: humans (n = 20), chicken breast (n = 65), ground turkey (n = 11), and pork chops (n = 2).
MIC, minimal inhibitory concentrations.
MDR was more common in C. coli than in C. jejuni from all sources, with EryR-DoxR (11.2%) being the dominant phenotype observed among overall C. coli isolates. Among C. jejuni isolates recovered from humans, the CipR-DoxR MDR pattern (8.7%) was found to be the most prevalent (Table 2). Overall, C. coli isolates from ground turkey tended to be more MDR than C. jejuni including the CipR-DoxR (54.5%) and the CipR-EryR-DoxR (27%) than from any other meat type or humans. Both the C. coli isolates from pork chops exhibited the EryR-DoxR MDR pattern.
Represents the number of Campylobacter isolates tested from different sources—C. jejuni: humans (n = 148), chicken breast (n = 110), and ground turkey (n = 4); C. coli: humans (n = 20), chicken breast (n = 65), ground turkey (n = 11), and pork chops (n = 2).
Number and percent of isolates displaying resistance to antimicrobial(s).
Cip, ciprofloxacin; Dox, doxycycline; Ery, erythromycin; Pan, pansusceptible.
Virulence gene profiles
Overall, virulence determinants were found to be more prevalent in isolates recovered from retail meats than humans (p < 0.01) (Fig. 1). The majority (95%) of Campylobacter isolates were positive for three or more virulence genes. Among the retail meat isolates, we found six C. jejuni (including five from chicken breast and a single from ground turkey) and a single C. coli isolate from chicken breast that were negative for all nine tested virulence determinants. All C. jejuni isolates from humans except one were positive for the cadF adhesion gene. The cadF gene was found significantly higher in C. jejuni from humans than from retail meats (p < 0.05). Among the human isolates, flaA was present in a significantly higher number of C. jejuni isolates than in C. coli (p < 0.001). The cytolethal distending toxin gene cluster (cdtA, cdtB, and cdtC) was detected in both C. jejuni and C. coli from different sources, with a higher proportion of C. jejuni from ill humans than for all the meat types combined. We detected significant differences in the prevalence of rakR, dnaJ, pld, and virB11 genes depending on the Campylobacter species and source. All Campylobacter isolates were negative for the virB11 gene, except for C. coli isolated from retail meats (45%). In addition, the rakR, dnaJ, and pld genes were found in a significantly higher number of C. jejuni isolates recovered from humans as compared to retail meats (p < 0.01). An important observation was that none of the C. coli isolates from humans possessed any of the above virulence determinants. However, a significantly higher number of C. coli isolates from retail meats were positive for the dnaJ, pld, and virB11 genes than C. jejuni isolates (p < 0.001).
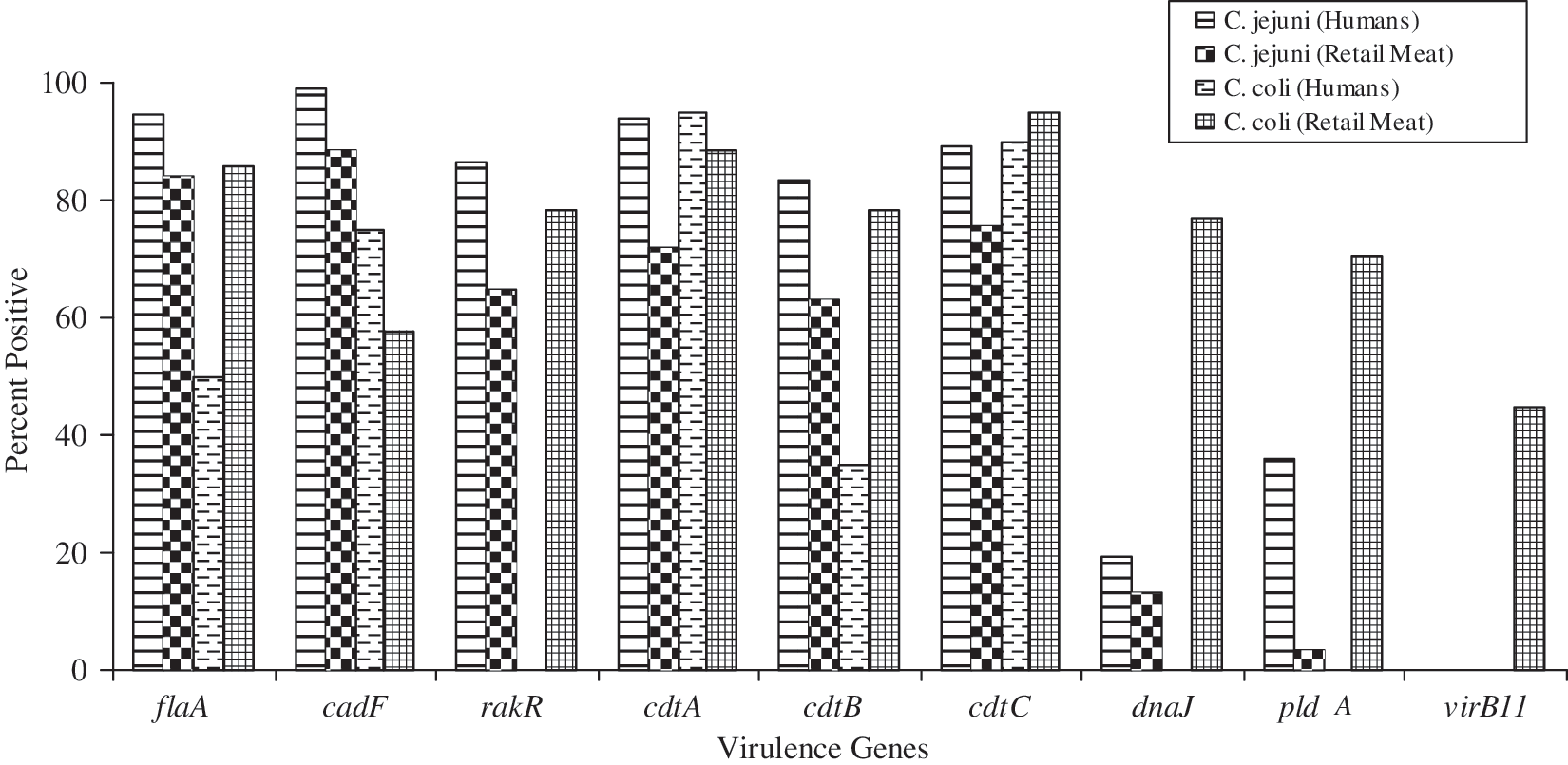
Prevalence of virulence genes among the Campylobacter jejuni and Campylobacter coli isolates recovered from humans and retail meats. The number of Campylobacter isolates tested from different sources include C. jejuni: humans (n = 148), chicken breast (n = 110), and ground turkey (n = 4); C. coli: humans (n = 20), chicken breast (n = 65), ground turkey (n = 11), and pork chops (n = 2).
Campylobacter PFGE profiles
A total of 252 C. jejuni and 98 C. coli isolates were first typed using SmaI restriction enzyme. The C. jejuni isolates were grouped into 182 unique PFGE patterns (represented by a single C. jejuni isolate) and 43 clusters that were represented by multiple isolates (data not shown). The majority (n = 31) of these 43 clusters were composed of isolates recovered from either ill humans or a particular retail meat. Sixty-seven unique SmaI PFGE patterns (represented by a single C. coli isolate) were identified among the 98 C. coli isolates, with 23 patterns had isolates from both humans and retail meats. We found three major clusters in the C. coli population including humans (n = 5), chicken breast (n = 5), and the largest cluster (n = 7) that was comprised of isolates predominantly recovered from chicken breasts with the exception of a single ground turkey isolate.
A total of 38 C. jejuni isolates representing the 43 common clusters generated by SmaI PFGE patterns were further analyzed with KpnI restriction enzyme. These 38 isolates were selected based on the source, date of isolation, and antimicrobial resistance profile. The KpnI digestions produced 33 patterns from the 38 isolates and were represented by three large clusters based on 60% similarity in their banding pattern (Fig. 2). Cluster A was the largest with 21 isolates, followed by clusters B (n = 8) and C (n = 9). Clusters A and C were composed of C. jejuni isolates from chicken breast and ill humans, with a single isolate each from a chicken breast and human having the same PFGE fingerprint but different virulence gene profiles. Cluster B was represented by isolates from chicken breast, ground turkey, and humans. We found identical subtypes shared between isolates from ground turkey and humans and between isolates from chicken breast and humans. KpnI analysis of the 23 C. coli patterns, which included 44 isolates, revealed no common patterns among isolates recovered from retail meats and humans (data not shown). We did, however, find four clusters composed of C. coli isolates from chicken breast. We also found several C. jejuni and C. coli isolates were nontypeable using either SmaI or KpnI enzymes and appears to be a case of DNA smearing rather than restriction.
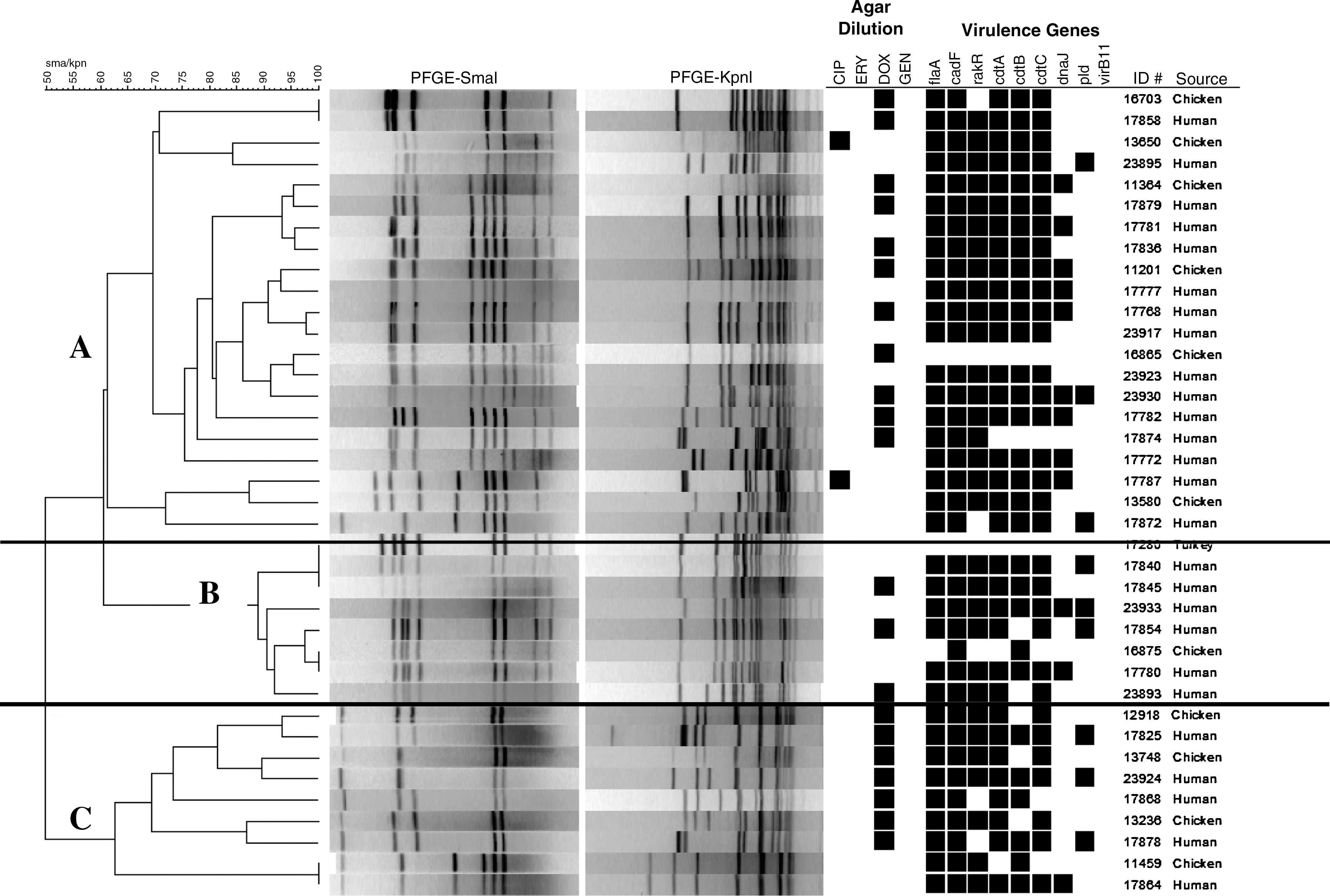
Dendrogram highlighting the pulsed-field gel electrophoresis genotypic patterns, virulence, and antimicrobial resistance profile of C. jejuni isolates recovered from humans and retail meats.
Associations between virulence genes, antimicrobial resistance, and genotypic profiles
An important aspect of this study was to determine whether significant associations between antimicrobial resistance and virulence determinants could be ascertained among the recovered Campylobacter isolates (Table 3). Among DoxR C. jejuni strains, those from human clinical cases (n = 94) and chicken breast (n = 41) were significantly associated with the majority of the virulence determinants tested, with odds ratio ranging from 3.42 for cdtB (95% confidence interval 1.5–7.7) to 21.63 for pld (95% confidence interval 3.5–128). No significant statistical association with any virulence gene was detected in CipR C. jejuni isolates either from humans or from chicken breast. We identified two clusters (based on 50% similarity) in the PFGE dendrogram generated by restricting C. jejuni and C. coli isolates (n = 29) with the two restriction enzymes that displayed resistance to both Cip and Dox (Fig. 3). None of the Campylobacter isolates in the dendrogram shared the same PFGE fingerprint. Cluster A was represented by both C. jejuni and C. coli isolates recovered from humans, chicken breast, and ground turkey. Cluster B was represented by only C. coli isolates recovered from the three sources as described above. Another distinct difference between the two clusters was the complete absence of virB11-positive isolates in cluster A. Both the EryR C. coli isolates, one each from chicken breast and ground turkey, were also restricted to cluster B.
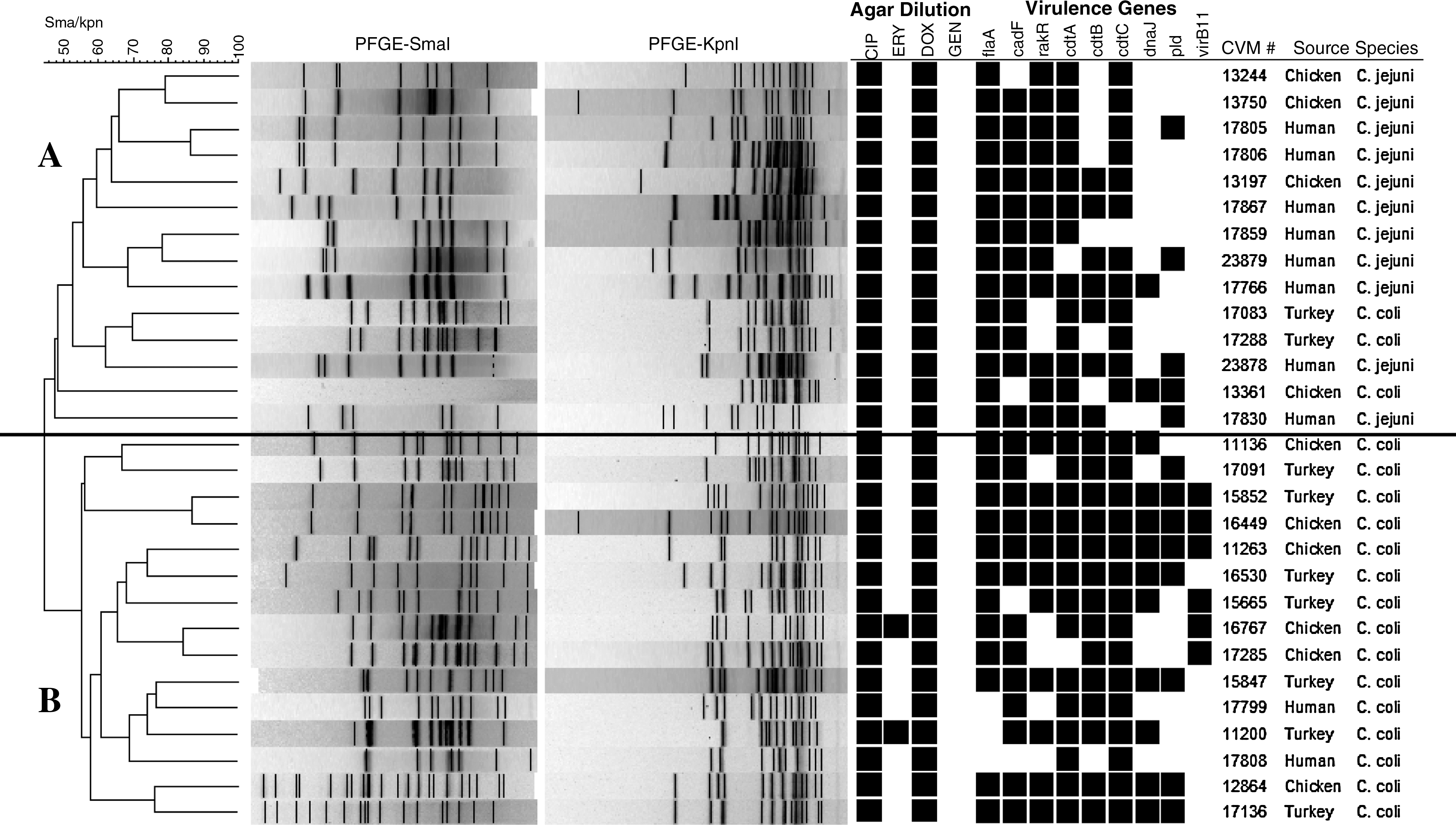
Genotypic association between virulence and antimicrobial resistance profile of C. jejuni and C. coli isolates from humans and retail meats.
Represents the number of Campylobacter species isolated from humans and chicken breast with a particular antimicrobial resistance profile; “R” denotes resistance.
Percent of isolates positive for tested virulence determinants.
Discussion
Campylobacter is among the leading causes of bacterial foodborne gastroenteritis cases in the United States, and consumption of contaminated retail meat products is a major risk factor for humans. Subgrouping of Campylobacter isolates is usually done by utilizing PFGE profiles in outbreak investigations and for studying epidemiological trends. Antimicrobial resistance provides an additional level of strain information that can help describe the epidemiology of Campylobacter. The antimicrobial susceptibility results of our study are comparable to others showing that resistance is generally more common in C. coli than in C. jejuni, including resistance to fluoroquinolones and macrolides (Ge et al., 2003; Thakur and Gebreyes, 2005; Thakur et al., 2006; Little et al., 2008). A recent study from Poland reported 40% of the C. jejuni isolates obtained from humans and chickens to be CipR (Rozynek et al., 2008), whereas another from the United Arab Emirates reported 85.4% of C. jejuni from humans to be fluoroquinolone resistant (Sonnevend et al., 2006). The retail meat samples in our study were collected from 2001 to 2002 when fluoroquinolones were still approved for use in the poultry industry. The impact of removal of these antimicrobials from the poultry industry on rates of resistance among Campylobacter in the United States in 2005 has yet to be fully determined. However, the National Antimicrobial Resistance Monitoring System 2007 executive report on retail meats highlights the steady increase in the frequency of CipR in C. jejuni up from 15.2% in 2002 to 17.2% in 2007, whereas in C. coli it is more concerning and has gone up from 10% in 2002 to 25.9% in 2009 (NARMS, 2007).
Among the isolates from retail meat, both C. jejuni and C. coli from ground turkey samples had the highest frequency of CipR, EryR, and DoxR. Our results are in agreement with other studies that have reported similar findings with larger numbers of Campylobacter isolates from ground turkey exhibiting antimicrobial resistance (Ge et al., 2003; D'lima et al., 2007). In the United States, EryR in C. coli isolates from ground turkey has gone up from 0% in 2002 to 22.2% in 2005 (NARMS, 2007). An increase in the percentage of EryR isolates is a concern since macrolide drugs are important for therapeutic treatment of severe campylobacteriosis cases in humans. MDR strains that exhibited resistant to both erythromycin and fluoroquinolone were found in higher numbers of C. coli than C. jejuni in humans and retail meats, which from a public health perspective is highly undesirable. However, the significantly higher number of humans campylobacteriosis cases caused by C. jejuni than C. coli highlights the importance of virulence determinants that aid in causing infection in the host.
Variation in the pathogenicity traits of isolates observed in this study, including adherence, motility, invasion, and toxin genes, has been reported previously (Coote et al., 2007; Fearnley et al., 2008). Campylobacter strains that are cadF mutants have been shown to be unable to colonize the chicken gut and have significantly reduced internalization ability into INT 407 intestinal cells (Ziprin et al., 2001; Monteville et al., 2003). Since all the human Campylobacter isolates were obtained from clinical cases and carried this marker (except one), it highlights the importance of this gene in establishing the infection. However, the mere presence of this one gene is not enough to set up infection, and it is important to determine the profile of other virulence determinants. A significantly higher proportion of C. jejuni than C. coli human isolates were positive for the flagellin-coding flaA gene. Motility expression via the flagella is an important virulence factor for Campylobacter to adhere and invade the intestine (Konkel et al., 2001; Malik-Kale et al., 2007), and strains that carry a mutation in this gene have severely reduced motility and are unable to colonize the chicken and human gut mucosa (Konkel et al., 2004). These findings may help explain why C. jejuni is much more common as a cause of human infections (90%–95%) than C. coli (5%–10%). The cdtB gene encodes the active subunit of the toxin, whereas the cdtA and cdtC gene products are responsible for the binding of the holotoxin (Lara-Tejero and Galan, 2001; Lee et al., 2003). Since all the three cdt gene products are required for the functional activity of the cytotoxin (Lara-Tejero and Galan, 2001; Asakura et al., 2007), the absence of the cdtB gene in significant numbers among C. coli isolates from humans could possibly explain the reduced potential of C. coli to cause infections in humans. However, it is important to mention that the presence or absence of a few genes cannot be used to predict whether a particular species of Campylobacter is virulent or not, and that further studies have to be conducted to make valid conclusions. In addition, a negative result by PCR can be caused by sequence variation at the primer binding site and does not necessarily indicate the absence of a gene. This is a limitation of the PCR method for detecting target regions and can have an impact on the data that should be interpreted carefully.
The PCR results for the rakR, dnaJ, pldA, and virB11 genes depicted an interesting profile. The rakR, dnaJ, and pldA genes were found in significantly higher numbers in C. jejuni from humans than from retail meats (p < 0.01). All Campylobacter isolates, except C. coli recovered from retail meats (45%), were negative for the virB11 gene. The above virulence genes were recently reported in 100% of C. jejuni strains (n = 40/40) isolated from human stool samples in Bangladesh (Talukder et al., 2008). This distinction could possibly be attributed to geographic location of the isolates. However, none of the C. coli isolates from humans in our study were positive for the rakR gene, which is involved in thermoregulation, and mutants have been shown to have a significantly reduced colonization potential in chickens (van Vliet and Ketley, 2001; Li et al., 2008). Similar observations were made when we compared the other important virulence genes in C. coli. The complete absence of the dnaJ, pldA, and virb11 genes in C. coli isolated from human strongly highlights the reduced potential of these strains to cause infection in humans. However, all the C. coli isolates from humans in this study were recovered from clinical cases; therefore, it is quite possible that the patients were suffering with a mixed Campylobacter infection. It is also important to highlight that the exact pathogenesis of Campylobacter infection in humans has not yet been fully elucidated. Therefore, it is quite possible that C. coli cause disease in host by a different mechanism compared with C. jejuni. Overall, the marked differences in the prevalence of virulence genes between C. jejuni and C. coli isolated from humans and retail meats in our study suggests that not all Campylobacter strains from retail meats are able to cause disease in humans.
Genotyping the Campylobacter isolates from humans and retail meats in this study highlighted the diversity in the population. Although we found clusters (SmaI) that were composed of isolates from different sources, the ability of the second restriction enzyme (KpnI) to further differentiate within these clusters highlighted the importance of using a second enzyme to make valid interpretations on the relationships shared between different Campylobacter strains. Although stable clones of Campylobacter have been described, the utility of PFGE for typing Campylobacter is limited, due to the high plasticity of the genome (On et al., 1998). Campylobacter has a hypervariable genome, which can be the result of many factors, including intra-genomic and inter-genomic recombinations and bacteriophage predation (Wassenaar et al., 1995, 1998; On et al., 1998; Weijtens et al., 1999; Scott et al., 2007). In a previous study, we genotyped the C. coli isolates from this study using multilocus sequence typing and found the overall population to be genotypically diverse (Thakur et al., 2009). Only a minority of isolates in our study showed genotypic similarity between humans and retail meats, which is in agreement with results of recent studies and indicates the potential role of other sources of Campylobacter transmission for humans (Uzunović-Kamberović et al., 2007; Gormley et al., 2008). There were four groups of C. jejuni isolates with indistinguishable PFGE patterns when restricted with one or two enzymes. The detection of 100% similar PFGE patterns in isolates from humans, turkey, and chicken (Fig. 2) after using two restriction enzymes does point to the ability of these C. jejuni strains to infect multiple hosts. However, this does not indicate that C. jejuni was transmitted from either the chicken or the turkey to humans in our study. Such a conclusion can only be reached in trace-back studies conducted in disease outbreak investigations.
An important aim of this study was to determine whether there was a significant association between antimicrobial resistance and virulence profile at the statistical and genotypic levels. Contrasting information is available in the literature on the relationship between clonally related isolates and their virulence or antimicrobial resistance profiles. For example, associations between PFGE type, biotype, in vitro virulence properties, and source have been reported in Campylobacter isolates from different sources (Nadeau et al., 2003; Hänel et al., 2007). In our study we did find DoxR C. jejuni isolates from humans significantly associated with majority of the virulence genes. Antimicrobial resistance to tetracycline is coded by the tet(O) plasmid, which also confers resistance to doxycycline (Thakur and Gebreyes, 2005). Plasmids have been shown to code for virulence traits, and it is possible that the presence of such a plasmid is providing these isolates with virulence potential (Friis et al., 2007). It is important to highlight that this statistical association was not replicated at the genotypic level for all the virulence genes. Similarly, a recent study reported the absence of any association between PFGE type and the virulence properties of C. jejuni strains isolated from humans and poultry (Van Deun et al., 2007). Therefore, it is important to determine whether statistical association is also correlated with genotyping results before making valid conclusions.
A few studies have reported that fluoroquinolone-resistant Campylobacter have the ability to cause severe infection characterized by prolonged diarrhea and clinical outcome in humans as compared to antimicrobial susceptible strains (Nelson et al., 2004; Helms et al., 2005). Recent work on Enterococcus faecalis from retail foods revealed both positive and negative associations between antimicrobial resistance and virulence determinants (McGowan-Spicer et al., 2008). In another study, positive statistical associations between resistance and virulence in Escherichia coli isolated from healthy pigs were reported with the authors concluding that antimicrobial resistance has the potential for selecting virulence in bacteria that may contaminate food products and animals (Rosengren et al., 2009). No such association was detected in CipR C. jejuni isolates from either humans or retail chicken in our study. A similar observation was made in E. coli isolates recovered from human blood and feces where no association was observed between fluoroquinolone resistance and virulence genes (Bagger-Skjøt et al., 2007). It is important to mention that association between virulence and antimicrobial resistance could be dependent on the bacterial population, strain, source, and other important factors, which must be taken into account before making valid interpretations.
In conclusion, the presence of different combinations of virulence gene determinants clearly indicates that Campylobacter strains with diverse pathogenic potential are circulating in the retail meat and human populations. To the best of our knowledge, this is the first study to determine whether any association exists between the virulence and resistance profile at the statistical and genotypic levels in Campylobacter. It is, however, difficult to define a relationship between virulence and antimicrobial resistance in Campylobacter. This is especially true in clinical cases since Campylobacter pathogenesis has not been fully elucidated. It is evident from our study that C. jejuni has an advantage over C. coli in causing clinical cases in humans primarily due to carriage of important virulence determinants that are essential for eliciting its pathogenesis. To fully understand Campylobacter pathogenesis, DNA microarrays and in vivo models will have to be developed to determine how the virulence genes work together to establish the infection. Even though the samples and isolates were temporally and spatially related, the genotypic diversity observed by PFGE typing in our study highlights the importance of conducting further detailed studies to better understand the epidemiology of Campylobacter in the environment and its role in transmission to humans.
Footnotes
Disclosure Statement
No competing financial interests exist.