
Abstract
Several foodborne human pathogens, when exposed to harsh conditions, enter viable but nonculturable (VBNC) state; however, still open is the question whether VBNC pathogens could be a risk for public health, because, potentially, they can resuscitate. Moreover, cultural methods for food safety control were not able to detect VBNC forms of foodborne bacteria. Particularly, it has not been established whether food chemophysical characteristics can induce VBNC state in contaminating pathogen bacterial populations, especially in food, such as salads and fresh fruit juices, not subjected to any decontamination treatment. In this preliminary study, we intentionally contaminated grapefruit juice to determine whether pathogen bacteria could enter VNBC state. In fact, grapefruit juice contains natural antimicrobial compounds, has an average pH of about 3 and low content in carbohydrates. Such characteristics make grapefruit juice a harsh environment for microbial survival. For this purpose, Escherichia coli O157:H7, Salmonella enterica serovar Typhimurium ATCC 14028, Listeria monocytogenes ATCC 7644, and Shigella flexneri ATCC 12022, at two different inoculum sizes, have been used. Viability by the LIVE/DEAD BacLight Bacterial Viability kit and culturability by plate counts assay were monitored, whereas “resuscitation” of nonculturable populations was attempted by inoculation in nutrient-rich media. The data showed that L. monocytogenes lost both culturability and viability and did not resuscitate within 24 h independently on inoculum size, whereas E. coli O157:H7 was able to resuscitate after 24 h but did not after 48 h. Salmonella Typhimurium and S. flexneri, depending on inoculum size, lost culturability but maintained viability and were able to resuscitate; moreover, S. flexneri was still able to form colonies after 48 h at high inoculum size. In conclusion, entry into VBNC state differs on the species, depending, in turn, on inoculum size and time of incubation.
Introduction
However, the existence of VBNC state is actually a matter of debate due to contrasting results. Optimal growth conditions are reportedly effective methods for rescuing bacterial populations from the VBNC state (Roszak et al., 1984; Smith et al., 1994; Ravel et al., 1995); but they may simply support the growth of a small number of undetected surviving culturable cells, giving the illusion of recovery (Ravel et al., 1995; Bogosian et al., 1996, 1998).
Whether or not the VBNC state could truly be a survival strategy or a simple artifact, the loss of culturability in a stressed bacterial population seems not to be questionable.
Relatively few studies have been reported on the presence of VBNC cells in food (Rowan, 2004) and, although the possible transmission of VBNC pathogens by food has been suggested (van Duynhoven and de Jonge, 2001), this remains a much underinvestigated area of food safety.
Outbreaks of human infections transmitted by raw and minimally processed fruits and vegetables have increased during the past decades, due to changes in dietary habits, incorrect agronomic and processing practices, global trading among countries with different hygienic standards, and the emergence of unusual associations of microbial pathogens with raw materials (Bean et al. 1997; Beuchat and Ryu, 1997).
Ready-to eat fruits and vegetables, such as salads and juices, are not subjected to any antimicrobial treatment, so they cannot be microbiologically safe, because they are susceptible to contamination by foodborne pathogens (Gould, 1992; Breidt and Fleming, 1997; Beuchat, 2002; Lanciotti et al., 2004).
Up to date, it remains unclear whether the chemophysical characteristics of ready-to-eat food, such as osmolarity, pH, free water content, and storage time and temperature, could induce a pathogen contaminating population to enter VBNC state.
Particularly, grapefruit juice is one of the most frequently demanded juices by consumers. It is a good source of vitamin C and contains high quantities of polyphenol antioxidants (Vinson et al., 2002). Moreover, due to low content in carbohydrates, it is used for weight loss (Fujioka et al., 2006), for beneficial effects on serum cholesterol (Borradaile et al., 1999), and in patients suffering from coronary artery disease (Gorinstein et al., 2004).
As chemophysical characteristics, grapefruit has a very low pH value, due to the content of organic acids (Cancalon and Xu, 2002; Soyer et al., 2003) and antimicrobial compounds present in the peel (Negi and Jayaprakasha, 2001), seeds, and pulp (Cvetnić and Vladimir-Knezević, 2004; Xu et al., 2007).
Such characteristics support the idea that grapefruit juice could induce VBNC state in contaminating bacteria.
On the other hand, grapefruit juice is a heterogeneous matrix, difficult to investigate, due to viscosity and pulp residues; moreover, the residing microflora could also play a role in contaminations by exogenous bacterial populations.
Then, the aim of this preliminary work was to study whether refrigerated grapefruit juice, after removal of solid particles and pasteurization, was able to induce VBNC state in foodborne pathogens, such as Escherichia coli O157:H7, Listeria monocytogenes ATCC 7644, Salmonella enterica serovar Typhimurium ATCC 14028, and Shigella flexneri ATCC 12022.
Materials and Methods
Grapefruit juice preparation
Five liters of juice were obtained from about 10 kg of yellow grapefruit, by squeezing fruits with Oranfresh® Vitaljuicer home centrifuge (AAT S.p.A., Catania, Italy).
Aliquots of 250 mL of yellow grapefruit juice were centrifuged (Beckman J2-21, 2000 g, 5 min, 4° C) to eliminate pulp residues, pH was measured, and then all aliquots were pasteurized (Cortés et al., 2006) before inoculation with the appropriate pathogen. To treat the samples, an Armfield FT74P unit with a plate exchanger was used. Juice placed in a feeding tank was impulsed by a pump to the heat exchanger, where the treatment conditions (90°C, 20 sec) were reached. After treatment, the juice was cooled with cold water from a cooler (Armfield FT61) and aseptically stored at 4°C until use. After pasteurization, pH was measured again, to establish whether thermal treatment would have modified acidity value, and no variations could be recorded.
Bacterial strains
E. coli O157:H7 was kindly provided by G. Bisignano (Department of Chimico-Biologico, University of Messina, Italy). L. monocytogenes ATCC 7644, Salmonella enterica serovar Typhimurium ATCC 14028, and S. flexneri ATCC 12022 were purchased from American Type Culture Collection (ATCC, Manassas, VA) and used in all the experiments.
Cryopreserved glycerol stocks of each strain, kept in 50% glycerol at −80°C, were put at −20°C to thaw for about 1 h, then were inoculated in 20 mL of rich medium contained in 100-mL Erlenmeyer's flasks, and incubated overnight at 37°C under shaking conditions. Tryptone soya broth (TSB; Oxoid, Cambridge, United Kingdom) for L. monocytogenes ATCC 7644 and Luria Bertani broth (Oxoid) for the others strains were used.
Inoculation of grapefruit juice with pathogens
Overnight cultures of each strain, corresponding to about 1 × 109 cell/mL, were centrifuged (Beckman J2-21, 9000 g, 10 min, 25°C), washed twice with isovolumes of sterile phosphate-buffered saline solution (PBS; Sigma, St. Louis, MO), inoculated in 30 mL of pasteurized grapefruit juice, to two distinct final inoculum sizes of about 1 × 104 (low-inoculum-size-culture [L-culture]) and 1 × 107 colony-forming units (CFU)/mL (high-inoculum-size-culture [H-culture]) respectively, and incubated at 4°C for 48 h. For L- and H-cultures, three replicates have been setup for each strain.
Analysis of culturability by plate count assay (CFU)
Culturability, intended as the ability to form colonies on solid media, was monitored at 0, 24, and 48 h and was assayed by plate counts and estimation of CFU. One hundred microliter aliquots were withdrawn from the three replicates of each culture and then progressively diluted in 900 μL of sterile PBS until a final factor of 10−10. One hundred microliter aliquots of each dilution and of undiluted cultures were spread in triplicate on plate count agar (PCA) (Oxoid) and incubated for 24–48 h at 37°C. Dilutions with number of colonies ranging from 30 to 300 were considered, and the mean of all the replicates was used as CFU/mL value. Results are expressed as the mean ±SD of the log10 CFU/mL.
Evaluation of viability by direct viable count
A fluorescent dye exclusion assay was used to assess the viability of cells in H-cultures. This was achieved with direct viable count by using the LIVE/DEAD BacLight Bacterial Viability kit (Invitrogen, Carlsbad, CA), which differentiates between bacteria with intact and damaged membranes. The kit is based on the principle that cells with intact membrane should be considered alive, whereas cells with damaged membrane should be considered died. Even if some exception has been observed, the test is commonly used to assess cell viability, because it shows a good correlation with standard plate counts (Terzieva et al., 1996; Bester et al., 2005).
Daily, 1-mL aliquots were withdrawn from each replicate of H-cultures; cells were harvested by centrifugation at 9000 g at 4°C for 10 min (Eppendorf 5417R centrifuge), washed twice with 1 mL of sterile PBS, and resuspended in 1 mL of 0.25% bovine serum albumin saline solution. The LIVE/DEAD BacLight kit was used according to the manufacturer's instructions. Briefly, 3 μL of the dye mixture were added to 1 mL of bacterial sample and incubated for 15 min at room temperature in the dark. The stained samples were filtered on a 0.22 μm pore size polycarbonate black membrane filter (Osmonics, Inc., Minnetonka, MN) and directly analyzed for red and green fluorescent cells by epifluorescence microscopy. A Leica DMRE epifluorescence microscope, fitted with 10 × eyepieces and 40 × and 100 × (oil immersion) objectives and excitation filter of 450–490 nm, was used to count intact (green fluorescence) and membrane-damaged (red fluorescence) bacterial cells. For each grapefruit juice culture, 30 microscopy fields, containing at least 150 cells, were counted; and the corresponding values were then referred to 1 mL and expressed as cells/mL. Results were expressed as the mean ± SD of the log10 cells/mL.
VBNC resuscitation
When no colonies were observed on PCA, 1-mL aliquots of all three replicates of each grapegruit juice unculturable culture were inoculated in 30 mL of either phosphate-buffered TSB (p-TSB; Oxoid), containing 1.5 g/L of K2HPO4, or buffered peptone water (BPW; Oxoid) (Reissbrodt et al., 2002); 25 aliquots, each of 300 μL, were, then, transferred in honeycomb plates (Thermo Fisher Scientific, Inc., Waltham, MA) (a total of 75 aliquots for each strain in each resuscitation medium). Samples were incubated at 37°C under shaking conditions; optical densities at 600 nm were measured at 20-min intervals over a period of 72 h by Bioscreen C (Thermo Fisher Scientific, Inc.).
To exclude the possibility of contaminations and to verify colony-forming ability restoration, any growth would have been confirmed in triplicate with either cultural techniques on selective agar media, McConkey agar (Oxoid) for E. coli O157:H7, SS agar (Oxoid) for both Salmonella Typhimurium and S. flexneri, L-palcam (Oxoid) for L. monocytogenes, specifically, or with biochemical profile by API tests (Biomérieux, Marcy l'Etoile, France), specifically API 20 Listeria for L. monocytogenes and API 20E for the other strains.
Results for each strain were resumed as the number of resuscitating aliquots and percentage.
Statistical analysis
For both viability and culturability cell counts, data from different experimental groups were compared using the one-way analysis of variance and Tukey-b test for post hoc analysis. All statistical values were considered significant at the p level of 0.05. Statistical analysis was performed using Instat version 2.10 for Microsoft Windows (GraphPad Software Inc., La Jolla, CA).
Results
Culturability of L-cultures is represented in Figure 1. Culturability and viability of H-cultures are represented in Figure 2. Resuscitation of VBNC forms in BPW and p-TSB media is summarized in Table 1.
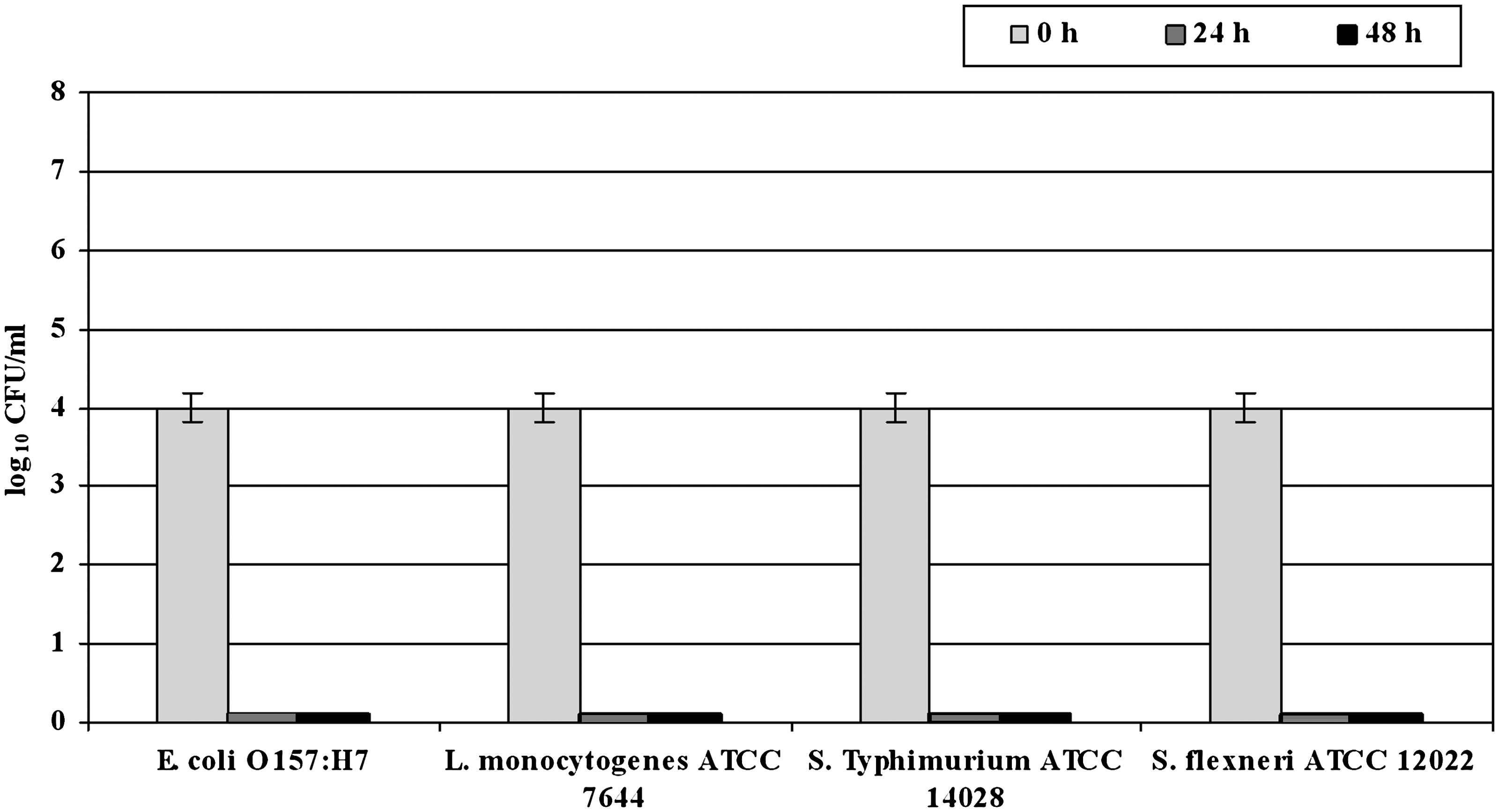
Culturability of low-inoculum size (104 cell/mL) cultures (L-cultures) of Escherichia coli O157:H7, Salmonella enterica serovar Typhimurium ATCC 14028, Listeria monocytogenes ATCC 7644, and Shigella flexneri ATCC 12022 in grapefruit juice incubated at 4°C, assayed at 0, 24, and 48 h by plate counts. Results are expressed as the mean ± SD of the log10 cells/mL on three replicates for each strain. All the strains lost culturability within 24 h.
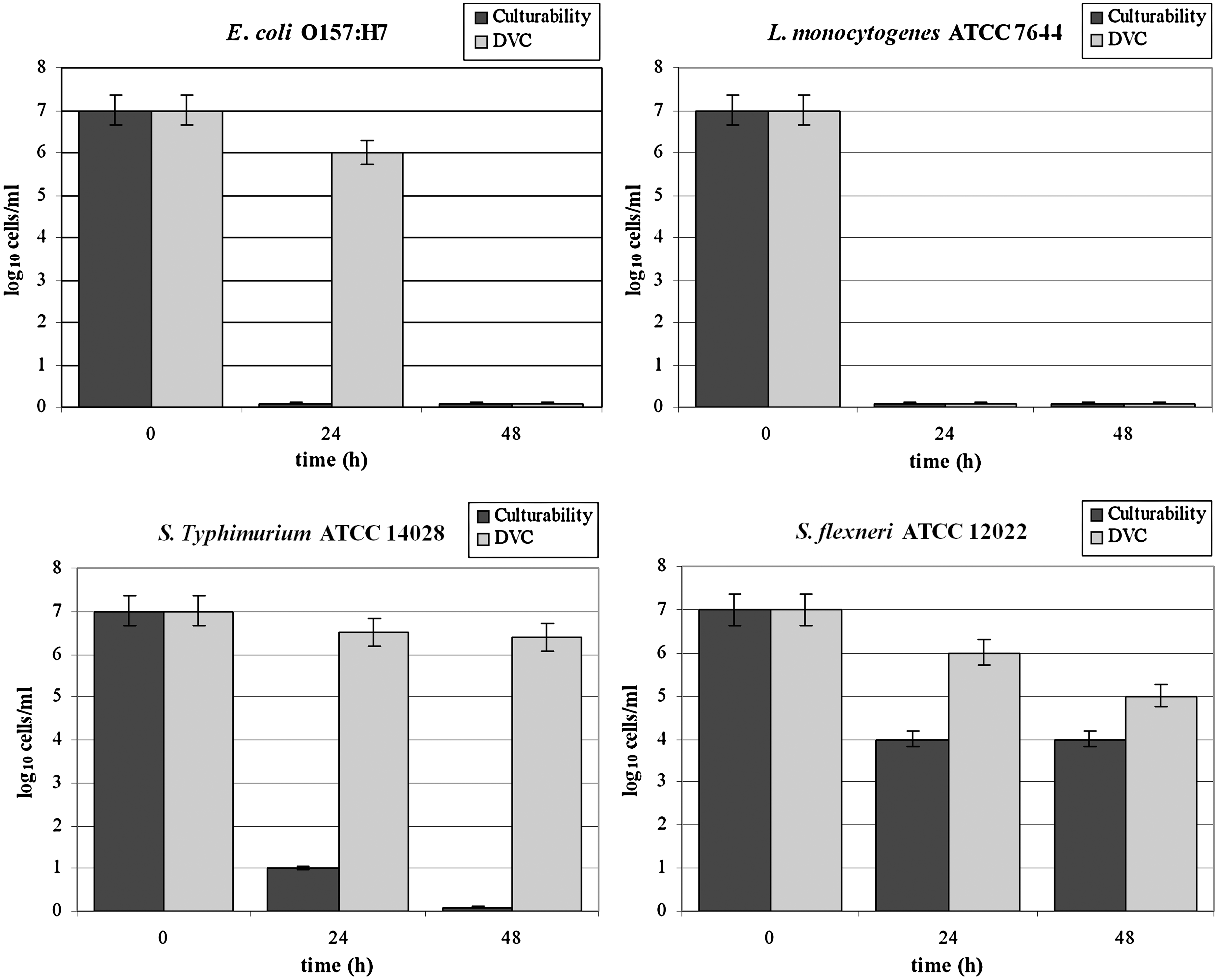
Culturability and viability of high-inoculum size (107 cells/mL) cultures (H-cultures) of E. coli O157:H7, Salmonella enterica serovar Typhimurium ATCC 14028, L. monocytogenes ATCC 7644, and S. flexneri ATCC 12022 in grapefruit juice incubated at 4°C, assayed by plate counts and LIVE/DEAD BacLight Bacterial Viability kit, respectively, at 0, 24, and 48 h. Results are expressed as the mean ± SD of the log10 cells/mL on three replicates for each strain. E. coli O157:H7 lost culturability within 24 h, both viability and culturability between 24 and 48 h; L. monocytogenes lost both culturability and viability within 24 h; Salmonella Typhimurium lost culturability but retained viability; S. flexneri was still culturable at 48 h. DVC, direct viable count.
From the three replicates of all unculturable L- and H-cultures, samples were inoculated in BPW and p-TSB media, partitioned in 25 aliquots (a total of 75 aliquots for each strain for each resuscitation medium), and incubated at 37° C. Growth (OD600) was monitored each 20 min for 72 h by Bioscreen C. L. monocytogenes did not resuscitate; Escherichia coli O157:H7 had 100% of aliquots able to resuscitate; for Salmonella Typhimurium and S. flexneri, only a fraction of the total 75 aliquots, with regard to each inoculum size and each resuscitation medium, was able to resuscitate.
BPW, buffered peptone water; n.p., not performed, because no fully unculturable state could be observed; p-TSB, phosphate-buffered tryptone soya broth.
In L. monocytogenes ATCC 7644 L-cultures, culturability decreased to zero within 24 h, as no colony growth was observed on PCA plates. No resuscitation has been observed after inoculation in resuscitation media. After 48 h, unculturability was confirmed and, as for 24-h-old L-cultures, no resuscitation has been detected.
In L. monocytogenes ATCC 7644 H-cultures, both viability and culturability decreased to zero within 24 h. Resuscitation, after inoculation in BPW and p-TSB, failed, as none of the aliquots were able to grow. Also, in 48-h-old H-cultures, viability and culturability were undetectable and no growth was obtained in resuscitation media.
In E. coli O157:H7 L-cultures, after 24 h of incubation, culturability was lost and resuscitation was observed in all 75 aliquots (100%) in both media. Resuscitated populations in BPW and p-TSB were then confirmed both by cultivation on solid selective medium and by API tests. Also, after 48 h, L-cultures were fully unculturable, but, in contrast to 24-h-old L-cultures, no resuscitation was detected in either BPW or p-TSB, as growth did not occur in any of the aliquots.
In E. coli O157:H7 H-cultures in grapefruit juice, after 24 h, viability was reduced of 1 log and culturability was totally abolished. After inoculation in BPW and p-TSB, resuscitation was observed in all 75 replicates (100%), resuscitated cultures were able to form colonies on solid selective medium, and API test identified such cultures as E. coli O157:H7. After 48 h, viability was no further detectable, and no culturable cells could grow on PCA plates. As observed in L. monocytogenes, after inoculation in resuscitation media, no growth has been observed in any of the aliquots.
In Salmonella Typhimurium ATCC 14028 L-cultures, culturability was not detected after 24 h, and resuscitation of 24-h-old L-cultures was obtained only in a fraction of aliquots, specifically 12 aliquots (16%) in BPW and 5 aliquots (6.7%) in p-TSB. As observed after 24 h, also in 48-h-old L-cultures, only a limited number of aliquots was able to resume growth in resuscitation media, specifically 10 aliquots (13%) in BPW and 5 aliquots (6.7%) in p-TSB. Resuscitated populations, then, were able to regenerate on solid selective media and were referred to Salmonella Typhimurium by API test.
In Salmonella Typhimurium ATCC 14028 H-cultures, within 24 h, viability decreased of 0.5 log, whereas culturability was almost lost, with 10 cells/mL still able to generate colonies on PCA. This was again the case after 48 hours, and remained unchanged through the end of observation period. Interestingly, as described for 24-h-old L-cultures, resuscitation involved only a fraction of the 24-h-old H-culture aliquots, corresponding to 21 aliquots in BPW (28%) and 12 aliquots (16%) in p-TSB. After 48 h, viability was kept constant, with regard to 24 h, and no culturable cells were detected on PCA plates. After incubation in resuscitation media, growth has been observed, as for 24-h-old aliquots, with identical percentages of resuscitation. Resuscitated Salmonella Typhimurium, after restreaking on solid selective media, generated colonies, and the corresponding metabolic profile was evidenced by API test.
In S. flexneri ATCC 12022 L-cultures, culturability dropped to zero within 24 h, as no growth of colonies has been observed when undiluted cultures in grapefruit juice were spread on PCA plates. Loss of culturability was confirmed also after 48 h of incubation. Twenty-four-hour-old and 48-h-old cultures were then inoculated in resuscitating media, and growth has been observed, in both cases, for 18 aliquots (24%) in BPW and 10 aliquots (13%) in p-TSB. Resuscitated cultures, after restreaking on solid selective medium, could grow and were metabolically confirmed by API test.
In S. flexneri ATCC 12022 H-cultures, within 24 h, viability showed a reduction of about 1 log with regard to the starting value; and a further decrease of 1 log occurred between 24 and 48 h. Culturability showed a drastic reduction of three logs within 24 h and, after 48 h, maintained an average value of 104 culturable cells/mL. Since 48-h-old cultures of S. flexneri did not become fully unculturable, no resuscitation experiments were performed.
Discussion
Fresh vegetable- and fruit-derived products are particularly susceptible to pathogen contamination, because they are not subjected to any antimicrobial treatment.
For pH value, low content in carbohydrates and the presence of natural antimicrobial compounds (Negi and Jayaprakasha, 2001; Cvetnić and Vladimir-Knezević, 2004; Xu et al., 2007), and storage time and temperature (usually, 4–8°C for 24–48 h), grapefruit juice is a very limiting environment for bacterial survival. For such characteristics, it can be considered as a good approach to investigate pathogen survival and the eventual entry into VBNC state.
The data raise several intriguing observations, even if the laboratory conditions do not allow their complete generalization on fresh juice.
First of all, grapefruit juice does not allow growth for any of the bacterial strains used in this study.
Moreover, VBNC state is not observed in all the strains, depends on inoculum size, and cannot be always considered as a long-term survival strategy.
In fact, grapefruit juice chemophysical conditions had a sterilizing effect on L. monocytogenes ATCC 7644, which lost both culturability and viability and was not able to resuscitate, independently on inoculum size.
Such results are not surprising, in that, despite its great significance as a foodborne pathogen, few studies have been reported on the ability of L. monocytogenes to enter the VBNC state (Oliver, 2005); and factors usually inducing VBNC state in other foodborne pathogens have no such effect in L. monocytogenes (Li et al., 2003). However, other studies described that several strains of L. monocytogenes enter VBNC state (Besnard et al., 2002; Cappelier et al., 2005). Moreover, some L. monocytogenes strains differed from each other in terms of time and factors inducing VBNC state (Besnard et al., 2002).
At least for this pathogen, it can be argued that grapefruit juice does not allow entry into VBNC state but induces massive cell death.
The other strains used in this study entered VBNC state, but factors such as time of incubation and inoculum size, whose effect has been described for several species (Ramadan et al., 1990; Ohno et al., 2003; Lang et al., 2004), play an important role.
For E. coli O157:H7, at least in our conditions, it seems that VBNC state does not confer any resistance to external injuries and appears as a transient phase, which precedes bacterial death.
In fact, in E. coli O157:H7, entry into VBNC state occurs within 24 h, involving the whole population, independently on inoculum size. Also, resuscitation is very efficient, because it was achieved in all the aliquots tested. However, between 24 h and 48 h, the population did not maintain VBNC state and died, as no further resuscitation has been observed in 48-h-old H-cultures (see Table 1).
In Salmonella Typhimurium ATCC 14028, the time of complete loss of culturability was depending on inoculum size, because in 24-h-old H-cultures, a few cells were still culturable, whereas 24-h-old L-cultures were already fully unculturable. Data clearly indicate that Salmonella Typhimurium ATCC 14028 entered the VBNC state, but, differently from E. coli O157:H7, not the whole population was affected, as indicated by resuscitation data of grapefruit cultures at both inoculum sizes (see Table 1). The data strongly indicate that VBNC state is not a transient phase, as for E. coli O157:H7, but is maintained for all the observation period, with no significant variations in the number of resuscitating aliquots (see Table 1); moreover, by comparing the loss of culturability in L- and H-cultures, it can be argued that it depends on inoculum size.
It has been suggested that genetic characteristics of both E. coli and Salmonella Typhimurium can explain their ability to survive in nonhost environments (Winfield and Groisman, 2003). In the same study, survival capacities have been compared and, according to the authors, the better ability of Salmonella to survive in nonhost environments is due to a better genetic versatility, with regard to E. coli, and may represent a critical adaptation to its cyclic lifestyle involving host and nonhost environments.
For S. flexneri ATCC 12022, inoculum size is a crucial factor that influences the entry into VBNC state. In fact, entry into VBNC state occurs in L-cultures within 24 h and is maintained also after 48 h, involving only a fraction of the population, as indicated by resuscitation data (see Table 1), whereas H-cultures show residual culturability and viability. Residual culturability of S. flexneri in refrigerated food has also been described (Islam et al., 1993).
Very few studies have described VBNC state in S. flexneri (Colwell et al., 1985).
Then, the role of inoculum size has to be taken in account when considering food safety control, due to its influence on residual culturability. In fact, detection of pathogens could fail if food is contaminated by a light load of bacteria.
Moreover, VBNC cells in L-cultures, as well as culturable cells in H-cultures, represent only a minor fraction of their respective populations. Such data suggest that the loss of culturability does not simultaneously involve the whole population, thus indicating that bacteria under food-induced stress have a nonlinear response.
Conclusions
Our preliminary study shows that grapefruit juice induces VBNC state in S. flexneri ATCC 12022, in Salmonella Typhimurium ATCC 14028, and, transiently, in E. coli O157:H7, but did not in L. monocytogenes ATCC 7644, indicating that not all the species react in the same way to grapefruit juice injuries.
However, entry into VBNC state differs on the basis of the species, the inoculum size, and the time of incubation.
An interesting observation is that in E. coli O157:H7, VBNC state does not confer prolonged resistance. In fact, even if the VBNC state involved the whole population after 24 h, as confirmed by resuscitation, it was not maintained after 48 h and no further resuscitation has been observed.
Such an observation lets us hypothesize that VBNC state is transient and that the loss of culturability observed after 24 h should be attributed to cell death and not a consequence of VBNC state itself.
On the contrary, VBNC state allows survival after 48 h in S. flexneri ATCC 12022 L cultures and in Salmonella Typhimurium ATCC 14028, but the population displays a heterogeneous behavior, because not all the cells are able to maintain viability and die. In such a case, VBNC state allows the survival of the species, but only in a subset of the population.
Moreover, in S. flexneri, but also in Salmonella Typhimurium, the time of entry into VBNC state depends on inoculum size, as in S. flexneri H-cultures after 48 h of incubation, the population did not become fully unculturable.
The data show that the VBNC state of foodborne pathogens induced by grapefruit juice is difficult to fit into a general schema, because its meaning seems to be dependent on different factors, such as bacterial species, time of incubation, and inoculum size.
Even if analysis of virulence is necessary to establish the possibility that hypothetical VBNC forms in fresh juice can be a risk for public health, food-induced loss of culturability and VBNC state in pathogens have to be considered when safety tests are performed on food contaminated by low loads of bacteria; moreover, “resuscitation” of food samples in appropriate media could be a useful implementation for eventual VBNC forms detection.
Footnotes
Disclosure Statement
No competing financial interests exist.