
Abstract
Contamination routes of Listeria monocytogenes were examined for 3 years in an Iberian pork-processing plant that produced high-quality ready-to-eat meat products. Molecular subtypes of L. monocytogenes were determined by polymerase chain reaction–based serotyping and pulsed-field gel electrophoresis (PFGE) restriction analysis. A total of 541 L. monocytogenes isolates were recovered from the environment and equipment (n = 165), carcasses (n = 28), raw products (n = 321), and dry-cured products (n = 27). Only 29 different PFGE types were identified, 3 of which were repeatedly found to be persistent types and accounted for 73% of the isolates. One PFGE type dominated (45% of the isolates) and was mostly recovered from intermediate manufactured products and the environment of the manufacturing area. L. monocytogenes persistence appeared strongly linked to the manufacture of products and not to its sustained entrance with the raw material. Some clones were found to survive in the manufacturing area for 3 years. Controlling the contamination of raw ingredients, improving the compartmentalization, and changing the cleaning protocols resulted in reduced prevalence rates of L. monocytogenes on products; two persistent PFGE types were eliminated from the processing plant, although eradication of other adapted strains has not been achieved.
Introduction
The Iberian pig is an autochthonous breed from the southwest Iberian Peninsula, traditionally fed under free-range conditions during the late finishing period in a Mediterranean forest ecosystem with acorn and grass fully available. Quality characteristics of these pigs are highly recognized for the market due to the added value of its cured products, mainly dry-cured ham, loin, and sausages (López-Bote, 1998).
The inherent characteristics of dry-cured meat products aid in the gradual reduction of pathogenic bacterial populations during storage, and cured products are usually considered safe. However, in the case of L. monocytogenes, small numbers of cells can persist for several weeks in dry sausages not made with starter culture (i.e., hard salami) (Farber et al., 2007). The level of risk associated with cured meat products may also depend on the extent and origin of contamination, storage temperature, and time between contamination and consumption (Simpson et al., 2008). Thus, although the risk of dry-cured product–associated listeriosis decreases over time, an opposite trend from most ready-to-eat products, the presence of L. monocytogenes in these products, should be considered and investigated (Simpson et al., 2008).
Palá and Sevilla (2004) reported that the percentages of isolation of Salmonella from carcasses of Iberian pigs were lower than those generally reported in the literature for white pigs, a name used to describe all other breeds raised under intensive conditions. For L. monocytogenes insufficient information is available on the contamination patterns in plants that produce Iberian pork products (López et al., 2008b). Different reports on molecular typing of L. monocytogenes from the environment and products of meat-processing plants indicate the presence of a limited number of strains or genotypes, together with the persistence of specific strains even after cleaning and disinfection (C&D) procedures (Farber et al., 2007). Isolates recovered from end products can share identical genotypes to those from the processing machines (Lundén et al., 2003). Some persistent strains have been found to be unique to that particular processing unit (Møretrø and Langsrud, 2004; Fugett et al., 2007), and control procedures targeting such specific strains must be established. Thus, the aim of this work was to check for L. monocytogenes subtypes in fresh and dry-cured pork products and both environmental and equipment surfaces of the Iberian pig slaughterhouse and processing plant, to identify contamination routes and possible persistence patterns. The final goal was to assist in the design of customized C&D practices. (The results of the first year of this study were presented in part at the International Symposium on Meat Safety: From Abattoir to Consumer, Valencia, Spain, February 14, 2007 [López et al., 2008b]).
Materials and Methods
Slaughterhouse and processing plant
The facilities investigated comprise the only Spanish slaughterhouse authorized to export pork meat products to the United States. It is a medium-sized industrial slaughterhouse (300 animals/day), annexed to a processing plant. Cured products were the main plant products, either as whole pieces (hams, shoulders, and loins) or Spanish traditional sausages “chorizo” and “salchichón.” Within these main five groups there are more than 30 different products with different sizes, formulations, ripening conditions, pH (4.3–6.3), and so on, and all, in principle, are not considered capable of supporting the growth of L. monocytogenes (water activity value of <0.92).
Surveys and sampling procedure
As a prominent step of the L. monocytogenes testing program in the facilities, the environment (noncontact surfaces) and equipment (food and nonfood contact surfaces), and the main product categories (carcasses, primal cuts, intermediate manufactured products, and end products) were regularly analyzed (Fig. 1). The samples analyzed had been collected throughout a 3-year period (August 2005–July 2008). Different classes of products were elaborated each month, and sampling sites and sampling frequency were considered based on features of the type of product being elaborated. In general, specific samples were taken monthly, but large samplings were carried out every 3–4 months.

Diagram showing the main processing stages of the plant (boxes). Capital letters indicate operations. Lower-case letters indicate raw materials and products. Boldface indicates isolation of Listeria monocytogenes. Parentheses indicate number of isolates obtained. Dashed line indicates manufacturing room. *Includes cold rooms and other areas (corridors, etc.).
Products (25 g) were sampled in the drying place at the end of the drying process (dry-cured products) or in the manufacturing room (intermediate manufactured products). All surface samples were collected using dry sponges (Whirl-pak; Nasco VWR International, Madrid, Spain) premoistened with sterile culture broth. Sponges were used to sample the area rigorously in both horizontal and vertical directions for 1 min. Carcasses were sampled by swabbing animals in the slaughterhouse before any cutting operation. Primal cuts were sampled also by swabbing in the cutting room. Environmental and equipment samples were collected after C&D, just before the begining of the working day, at each processing stage in the different plant areas; sampling locations were chosen to represent those most likely to harbor L. monocytogenes (Kornacki and Gurtler, 2007). The area of sampling varied depending on the sampling location. Minimum areas were knives or cutters (ca. 5 cm2); maximum areas were surfaces of conveyor belts, floors, or doors (up to 1 m2). In the case of floor drains and wastewater gutters, 50 mL of liquid was sampled for analysis.
Overall, a total of 2127 samples collected during the 3 years were tested for the presence of L. monocytogenes.
Isolation and identification of L. monocytogenes
Examinations for L. monocytogenes and other Listeria spp. were carried out according to the ISO method 11290 part 2 (Scotter et al., 2001). The USDA procedure for detection of L. monocytogenes (USDA, 2006) was also followed in selected cases. In the case of dry-cured products, simultaneous detection and enumeration was accomplished by spreading 0.1 mL of the enrichment homogenate on chromogenic agar (Hitchins, 2003).
In most samples, only 1 colony was isolated and tested, but from selected positive samples (mainly environment and carcass surfaces), several colonies (up to 10) from each chromogenic agar plate were randomly selected for further identification and typing, in an attempt to detect different strains in a single sample (Navas et al., 2007). The isolates were identified by the methods described in the Bacteriological Analytical Manual of the USFDA (Hitchins, 2003).
Polymerase chain reaction serotyping and pulsed-field gel electrophoresis typing
Isolates were classified in polymerase chain reaction (PCR)–based serotypes using a multiplex PCR assay, as described by Doumith et al. (2004). This PCR use four primer pairs that were specific for the four major serotypes of L. monocytogenes, 1/2a (or 3a), 1/2b (or 3b), 1/2c (or 3c), and 4b (or 4d and 4e), in addition to one primer pair that was specific for Listeria spp. (Doumith et al. 2004).
Pulsed-field gel electrophoresis (PFGE) was performed according to the Centers for Disease Control and Prevention PulseNet–standardized procedure for typing L. monocytogenes (Graves and Swaminathan, 2001; López et al., 2008a, 2008b), using a Clamped Homogeneous Electric Field system (CHEF DR II; Bio-Rad Laboratories, Madrid, Spain). Restriction enzymes AscI and ApaI (New England Biolabs, Beverly, MA) were used for cleaving the DNA. Composite PFGE types or pulsotypes were obtained by combining AscI and ApaI profiles and were designated with an “S” followed by a number identifying both the AscI and the ApaI profiles (López et al., 2008b); when more than one ApaI profile corresponded to one AscI profile, the first number identifying the AscI profile was followed by a hyphen and a second number identifying the different ApaI profiles (Fig. 2).
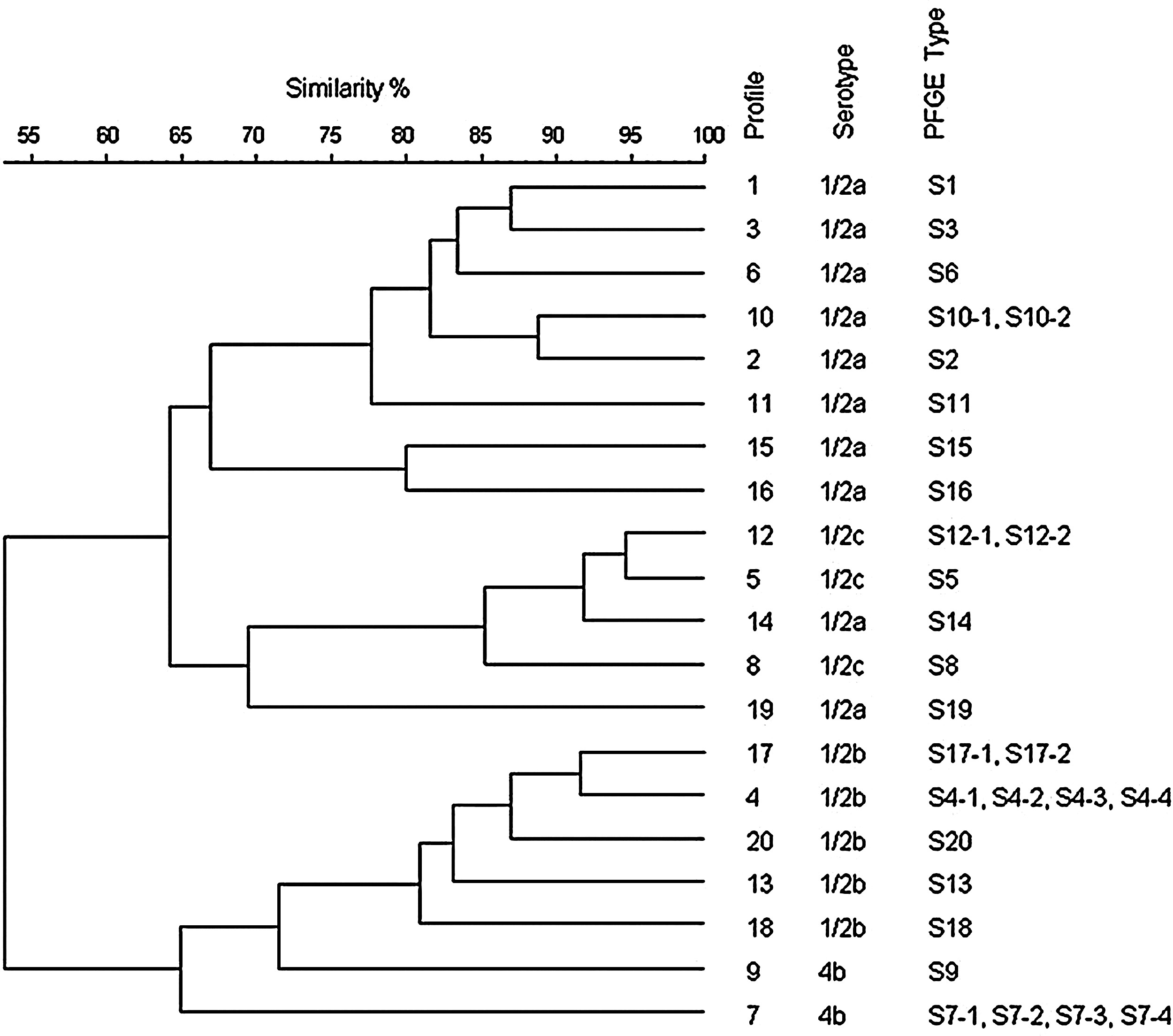
Dendrogram showing similarities among AscI restriction profiles of L. monocytogenes isolates. A total of 29 PFGE types resulted from the combination of these 20 AscI profiles and the 29 ApaI profiles. PFGE, pulsed-field gel electrophoresis.
Strain persistence
L. monocytogenes strains were considered persistent when they were found repeatedly (three times or more) in the environment or equipment over a minimum of 3 months (Keto-Timonen et al., 2007). Strains that were isolated sporadically (fewer than three times) or within a shorter time period (<3 months) were considered nonpersistent.
Results
Isolation of L. monocytogenes from Iberian pork products and environmental samples
There was a relatively low contamination rate in the environment and equipment, with 9% of positive samples (Table 1), L. monocytogenes being detected at some processing stages but not in others (Fig. 1). Altogether, as shown in Table 1, 24% of the samples of raw materials and products were positive. The evaluation of L. monocytogenes contamination on the different raw meats grinded, mixed, and marinated in the manufacturing room was validated as an effective critical control point in specific hazard analysis and critical control points (HACCP) plan. It significantly reduced the total number of contaminated products in the third year of the study (Table 1). On the other hand, the overall number of positive samples obtained from the environment and equipment did not vary essentially during the study period. Most of the contaminated products were intermediate-manufactured products, with end (dry-cured) products representing 6% of the contaminated products. Contamination level of end products usually ranged below 102 CFU/g, but in two samples of loin (11% of the contaminated dry-cured products) the level was 103 CFU/g.
Samples were collected between August 2005 and July 2006 (year 1) and the same months of 2006–2007 (year 2) and 2007–2008 (year 3).
During this 3-year period, a subset of 355 positive samples was subjected to strain identification by molecular subtyping. The number of isolates obtained from these samples is presented in Figure 1. From selected positive samples, several colonies were isolated and tested (see Materials and Methods), resulting in a total of 541 isolates identified as L. monocytogenes.
PCR-serotyping and PFGE typing of L. monocytogenes
Altogether, the four PCR-based serotypes (1/2a, 1/2b, 1/2c, and 4b) were detected, with two of them being predominant (1/2a and 1/2b, respectively, representing 68% and 21% of the isolates) (Table 2). Serotype 1/2a isolates were present in the environment and equipment, and in the different product categories, whereas serotype 1/2b isolates were mostly present in environmental sites and raw products. Serotypes 1/2c and 4b were less represented as shown in Table 2. Serotype 1/2c isolates were mainly associated with the slaughterhouse environment and carcass surfaces, whereas serotype 4b isolates were only present in raw products.
S, slaughterhouse; C, cutting room; M, manufacturing room; O, other zones.
+, persistent PFGE type.
PFGE, pulsed-field gel electrophoresis.
The PFGE analysis of isolates using AscI yielded 20 (S1–S20) restriction profiles. ApaI turned out to be adequate for discriminating four different profiles for AscI profiles S4 and S7, and two different profiles for S10, S12, and S17 (Fig. 2 and Table 2). Thus, analysis with ApaI yielded 29 different profiles, the same number of the pulsotypes obtained when the two enzymes were combined. The dendrogram of the PFGE profiles generated with AscI is shown in Figure 2. Similar results were observed with ApaI profiles (data not shown). Clustering of the L. monocytogenes pulsotypes in two groups at a 0.5 similarity level was observed. Fifteen pulsotypes belonging to serotypes 1/2a and 1/2c (lineage II) were grouped in one cluster, and 14 pulsotypes belonging to serotypes 1/2b and 4b (lineage I) were grouped in another. Isolates belonging to serotype 1/2c, mostly obtained in the slaughterhouse, formed one independent subcluster within the main lineage II cluster, together with S14 (1/2a) (Fig. 2).
Contamination patterns
The plant sites where one or more pulsotypes of L. monocytogenes were isolated are listed in Table 2. Contamination time span of each pulsotype is also shown in Table 2. Only eight pulsotypes seemed to persist in the plant during the study period, but they represented 91% of the isolates.
The slaughterhouse was contaminated with four persistent PFGE types (S2, S4-1, S5, and S10-1). The environment and equipment in the cutting room were contaminated with persistent PFGE types usually shared with the slaughterhouse (S2, S4-1, S5, and S10-1), the manufactuing room (S1, S4-1, and S5), or other zones (S4-2), the exception being S4-3 and S17-1, associated exclusively to the cutting room (Table 2). The manufacturing room was contaminated with three persistent PFGE types (S1, S4-1, and S5). In this area some production lines (e.g., loins) were persistently contaminated, and L. monocytogenes was found to be harbored in a loin-marinating machine. Further, a grinding machine was also shown to cause contamination of ground products.
Only 27 isolates came from dry-cured products (4% of the isolates) and they belonged to four different PFGE types (S1, S2, S4-2, and S12-1) (Table 2). On the other hand, 349 isolates from raw products (64% of the isolates) belonged to 25 different PFGE types.
Predominant pulsotypes
We arbitrarily considered as predominant those pulsotypes representing >10% of the isolates. Accordingly, three of the persistent pulsotypes, S1, S2, and S4-1, were predominant. They were isolated during all the 3-year study period from all kind of samples, and accounted for 73% of the isolates (Table 2).
The pulsotype S1 dominated (45% of the isolates), being the main pulsotype in the processing plant. However, it was not found in the slaughterhouse (neither on the carcasses nor in the environment). It comprised 92% of the isolates in the environment and equipment of the manufacturing area. This was the main pulsotype recovered from products (Table 2), mostly from whole intermediate manufactured products (e.g., loins before stuffing, where 73% of the isolates were S1).
On the other hand, the pulsotype S2 (16% of the isolates) was not found on the surfaces of the manufacturing area. It was only found during the second year in the environment of the slaughter and cutting areas and on several occassions on the surface of carcasses (Table 2). In spite of this distribution in the environment, it was more frequently associated with intermediate manufactured products (93% of the S2 isolates coming from products), mostly ground meat products. During the third year it was only detected during the first quarter (Table 2), and just twice (one product sample and one environmental sample), disappearing at the end of the study.
The pulsotype S4-1 (11% of the isolates) was irregularly found in the environment of the slaughterhouse and processing plant during the 3 years. The first year it was recovered from drain water samples and in a grinding machine of the manufacturing area, suggesting a possible association of this machine with the contamination of the ground meat products (López et al., 2008b). The second year it was only isolated from the environment and equipment of the cutting room, and during the third year it was only found in the environment and cleaning equipment of the slaughterhouse (10 isolates). With regard to its presence on carcasses and meat products, this is the third most frequent pulsotype (Table 2). It was similarly recovered both from whole and ground intermediate manufactured products. However, 85% of the isolates were obtained during the first year of the study and 15% during the second year, and the third year it was not detected in product samples.
Discussion
The Iberian pig industry presents several special features, deserving a better understanding of L. monocytogenes contamination patterns in Iberian pork-processing plants. Ripening periods of dry-cured Iberian products are usually longer (over 20 months for Iberian hams, compared to 7–14 months for dry-cured hams from white pigs, such as Serrano, Parma, or Bayonne hams) (Jiménez-Colmenero et al., 2009). Probably related to that, food technologies such as high hydrostatic pressure processing show different effect on the survival of L. monocytogenes in the case of Iberian ham compared to other dry-cured hams (Morales et al., 2006). In spite of this and other characteristics of the Iberian pig industry (i.e., diet and population density of the animals, and cutting operations of carcasses) (López-Bote, 1998), our results on the contamination flow of L. monocytogenes were not so different from previously published results in other pork-processing plants (reviewed by Thévenot et al., 2006).
During the first year of our study, L. monocytogenes was not detected in the environment of the slaughterhouse, and in the first and the third year the organism was not detected on the pig carcasses before processing. This low contamination rate in the slaughterhouse is similar to that reported in other studies indicating an incidence of listeriae ranging from 0% to 9% in pork carcasses and 0% to 3% on slaughter plant equipment (Gill and Jones, 1995; Farber et al., 2007).
Table 1 shows data on the presence of L. monocytogenes on environmental and equipment surfaces when sampled after the routine C&D protocol. Sampling revealed a moderate L. monocytogenes contamination in the processing areas of the plant (around 10% of positive samples, Table 1), similar to findings of other studies (Chasseignaux et al., 2001; Gudbjörnsdóttir et al., 2004; Kornacki and Gurtler, 2007).
With regard to raw material and products analyzed during the 3-year period, 24% were positive. The figures of contaminated products were not so different when compared to reported incidences for L. monocytogenes in meat products (Farber et al., 2007). However, in our case most of the contaminated products were intermediate manufactured products, with end products representing <10% of all the contaminated product samples. If we consider only the results of end products, it can be said that figures were lower than those previously published on incidence of L. monocytogenes on cured pork products (12%–22%; cited by Farber et al., 2007).
During the second year of the study, control of L. monocytogenes contamination of different raw ingredients entering the manufacturing room was used as a critical control point. Further, compartmentalization of the cutting room limited the flow of L. monocytogenes from the slaughterhouse into the processing plant, and the C&D protocols of the manufacturing area were changed. Main changes involved cleaning procedures, product application sequence, incorporation of new products (enzymatic cleaning agents designed for bacterial biofilm removal), and new disinfection procedures for equipment not covered so far. For example, machines used in the preparation of sausages were disinfected paying close attention to difficult-to-clean places where bacteria may easily hide. These changes had no clear effect on the overall isolation of L. monocytogenes from the environment (Table 1), probably due to the nature of the reservoir sites, but in the case of contaminated products, the number of positive samples decreased in the third year of the study period (Table 1).
Most outbreaks of listeriosis are caused by serotype 4b, even though serotype 1/2a is more frequently isolated from food and environmental samples (Gray et al., 2004). Therefore, it is likely that a number of factors are involved in the ability of a strain to cause foodborne disease or persist in a given food environment. For example, serotypes 1/2b and 1/2c have often been isolated from meat and meat-processing environments, but not from animals (Boerlin and Piffaretti, 1991; Gianfranceschi et al., 2003). Heir et al. (2004) also found that fresh meat isolates almost exclusively grouped within lineage I (serotypes 1/2b and 4b), and similar results were found in our previous study (López et al., 2008a), where serotype 1/2b represented 89% of the isolates in a broiler abattoir. However, in the present study, serotype 1/2a (lineage II) clearly predominated among isolates from raw products (74%). Overall, we also found a great number of strains belonging to serotype 1/2a (68% of all the isolates) that might be especially adapted to colonizing the manufacturing zone of the plant and were predominant on intermediate manufactured products. Although the reason for the dominance of serotype 1/2a is unclear, a previous study demonstrated that strains belonging to lineage II (serotypes 1/2a and 1/2c) are generally more capable of biofilm formation compared to lineage I strains (Borucki et al., 2003).
The PFGE analysis using AscI yielded 20 restriction profiles, and analysis with ApaI yielded 29 different profiles, the same number of the pulsotypes obtained when the two enzymes were combined (Fig. 2 and Table 2). Other observations (Brosch et al., 1996; Fugett et al., 2007) also indicate that ApaI PFGE may be more discriminatory than AscI PFGE when the number of PFGE types is the only fact considered, because more ApaI than AscI PFGE types are usually differentiated. These 29 different PFGE types represent potentially 29 different sources of contamination. Despite this significant number of pulsotypes encountered, eight pulsotypes included 91% of the L. monocytogenes isolates, and 17 pulsotypes (59%) were represented by one or two isolates (Table 2). Most of these sporadic pulsotypes were detected in carcasses and raw products, where the high diversity of sporadic strains may result from the range of different materials used in production. On the other hand, only four PFGE types were observed on dry-cured products, and all four were also detected in the cutting room, indicating contamination by fresh meat or fresh meat environments together with differential survival during the drying process of selected strains. This highlighted the need to re-address the design and cleaning of the cutting room and the flow for fresh and processed meat, as well as staff behavior (through new education programs on specific biological hazard). Simpson's index of diversity (Hunter and Gaston, 1988) can be calculated to estimate the genetic diversity present in the population sampled. Fugett et al. (2007) studied the L. monocytogenes PFGE type diversity across populations from different sources. They found that PFGE with AscI and ApaI exhibited a very high discriminatory power (Simpson's index of diversity [D] = 0.995). With the same two enzymes, the PFGE type diversity of our sample was calculated to be 0.749. This can be considered a relatively low genetic diversity for this group of 541 L. monocytogenes isolates obtained during 3 years in a single plant. Similar results on the genetic diversity of L. monocytogenes in pork-processing plants have been previously published (reviewed by Thévenot et al., 2006).
In the Iberian pork plant studied here, three predominant pulsotypes seemed to persist during the study period, which demonstrates that they are in-house strains of this plant environment. Because biofilm protects bacteria against disinfection (Møretrø and Langsrud, 2004), one hypothesis to explain persistence is that biofilm-forming abilities are linked to persistence. Such a linkage has been previously reported for strains of L. monocytogenes from bulk milk samples (Borucki et al., 2003) and from different food-processing environments, including meat-processing plants (reviewed by Møretrø and Langsrud, 2004). Being a good biofilm producer could be a logical explanation for cases where a single isolate is repeatedly isolated from a given environment (Borucki et al., 2003). That is the case of persistent pulsotype S1, which was recovered from the environment of the manufacturing room and products manufactured there during 3 years, suggesting a reservoir of this strain on product contact surfaces of this area. Autio et al. (2003) noted that strains of L. monocytogenes that were characterized as persistent within a plant were genetically distinct from those that were isolated only sporadically. Persistent strain S1 may be a clone specifically adapted to the environment of the manufacturing room of the Iberian pork-processing plant, being able to survive and grow there.
Conclusion
Reducing all contamination pathways of the manufacturing area to the maximum extent practicable was probably the most important factor in achieving desired food safety objectives. It had a significant effect on subsequent contamination of manufactured products and was probably the cause of the elimination of two persistent PFGE types (S2 and S4-1) at the end of the study. Therefore, special attention should be paid to raw ingredient contamination, compartmentalization of fresh meat environments, and continuous quality control of machines when manufactured foods are produced.
Footnotes
Acknowledgments
This work was supported in part by Embutidos Fermín, S.L., and the Spanish Ministry of Science Grants RTA2005-00202-C02-01, TRT2006-00048-00-00, and RTA2008-00080-C02 (INIA-FEDER). We are grateful to C. Jacquet (Insitut Pasteur, Paris, France) and L.M. Graves (Centers for Disease Control and Prevention, Atlanta, GA) for respectively providing strain H2446 and molecular typing protocols.
Disclosure Statement
No competing financial interests exist.