
Abstract
Salmonella Enteritidis is responsible for human gastroenteritis outbreaks worldwide, and the molecular characterization of isolates is an important tool for epidemiological studies. Fluorescent amplified fragment length polymorphism (FAFLP) analysis was performed on 31 Salmonella Enteritidis strains from South Brazil isolated from human, foods, swine, broiler carcasses, and other poultry-related samples to subtype isolates in comparison to pulsed-field gel electrophoresis (PFGE) analysis. Five strains of Salmonella Enteritidis from different geographical regions, Salmonella Enteritidis ATCC 13076, and four isolates of different Salmonella serovars were also tested. Among the 41 isolates tested, 96 polymorphic AFs and 40 distinct profiles were obtained, displaying a Simpson's index of diversity of 0.99; whereas the PFGE analysis presented 13 patterns and the resulting Simpson's index was 0.55. Nine FAFLP and seven PFGE clusters could be inferred based in Dice similarity coefficient. FAFLP clustering readily identified different serotypes of Salmonella but did not distinguish isolates epidemiologically nonrelated or distinct phage types. Therefore, these results indicate that FAFLP is a rapid method for epidemiological investigations of Salmonella outbreaks, presenting a high discriminatory power for subtyping of Salmonella Enteritidis.
Introduction
For outbreak investigation and epidemiological surveillance, a correct identification and characterization of the causal bacteria is essential. Serotyping and phage typing are classical phenotypic typing methods used to differentiate isolates of Salmonella (Ward et al., 1987; Olsen et al., 1994), but these methods alone are many times not sufficient for an accurate identification (Liebana et al., 2004). Thus, molecular characterization becomes an important tool for epidemiological studies. However, the investigation of foodborne outbreaks caused by Salmonella Enteritidis by molecular methods has been proved difficult, as this serovar displays great genetic homogeneity and the techniques most used for subtyping Salmonella Enteritidis show limited discriminatory power (Boxrud et al., 2007). Therefore, it is crucial that a higher discriminatory molecular typing technique be developed in a way to differentiate Salmonella Enteritidis strains (Foley et al., 2007), and the use of multiple methods is often required for an effective discrimination (Liebana et al., 2001; Oliveira et al., 2007).
Several molecular epidemiological techniques, such as pulsed-field gel electrophoresis (PFGE) (Ridley et al., 1998; Fernandes et al., 2003), plasmid profiling (Olsen et al., 1994; Miljkovic-Selimovic et al., 2008; Kalender et al., 2009), random amplified polymorphic DNA (RAPD) (Betancor et al., 2004), amplified fragment length polymorphism (AFLP) (Desai et al., 2001; Scott et al., 2001), and multilocus sequence typing (Kotetishvili et al., 2002; Fakhr et al., 2005), have been performed together with phenotypic techniques in an attempt to improve the reproducibility and discriminatory ability of Salmonella Enteritidis genetic typing. The implementation of these techniques has led to an enhanced detection of outbreaks worldwide (Winocur, 2003; Sukhnanand et al., 2005).
Fluorescent AFLP (FAFLP) is an accurate, rapid, and reproducible method based on fragmentation of genomic DNA with restriction endonucleases, followed by ligation of adaptors and amplification with selective or nonselective fluorescent labeled primers (Scott et al., 2001). The fragments generated by a large number of isolates can be directly analyzed by automated sequencing in a short period of time (Desai et al., 2001; Foley et al., 2009). In this sense, the aim of this study was to characterize Salmonella Enteritidis isolates from different sources by FAFLP to evaluate its discriminatory potential to this genetically homogeneous serovar.
Materials and Methods
Bacterial strains
Thirty-one Salmonella Enteritidis strains isolated from humans, foods, pigs, broiler carcasses, and poultry-related samples collected over the period 1995–2001 were evaluated by FAFLP. These strains were isolated from the Rio Grande do Sul and Santa Catarina States, Brazil, and phagotyped in preview studies (Santos et al., 2003; Oliveira et al., 2005). Five epidemiologically unrelated Salmonella Enteritidis strains isolated from other countries, Salmonella Enteritidis ATCC 13076, and four isolates of other serovars (Salmonella Senftenberg, Salmonella Typhimurium, Salmonella Panama, and Salmonella [4,5:-:1,2]) were also included (Table 1). Isolates were grown overnight in trypticase soy broth at 37°C.
FAFLP, fluorescent amplified fragment length polymorphism; PFGE, pulsed-field gel electrophoresis; ND, Not determined.
DNA extraction
A 1 mL aliquot of culture in trypticase soy broth was centrifuged at 12,000 g for 5 min. Cells were washed twice in 1 mL of 1 M NaCl and pelleted by centrifugation at 12,000 g for 3 min. They were resuspended in 100 μL of TE (10 mM Tris-HCl pH 8.0, 1 mM EDTA).
Genomic DNA was extracted according to the method described by Rademaker and de Bruijn (1997). Briefly, bacterial cells were lysed with 500 μL of 5 M guanidine thiocyanate, 0.03 M N-lauroyl sarkosine, and 0.1 M EDTA for 5 min at 4°C. Later, 250 μL of cold 7.5 M ammonium acetate were added; tubes were gently shaken and incubated for 5 min at 4°C. An aliquot of 500 μL of chloroform/iso-amyl-alcohol (24:1) was added, and the mixture was vigorously vortexed. After centrifugation at 16,000 g for 10 min, the DNA-containing pellet was further washed with isopropyl alcohol. The DNA samples were stored at −20°C. The DNA quantification was performed in a spectrophotometer (Spectronic Genesys 8) according to Sambrook et al. (1989).
FAFLP typing
FAFLP was performed using ∼400 ng of genomic DNA from each isolate and simultaneously digested with 2.5 U of EcoRI and MseI restriction endonucleases (Invitrogen) in a total volume of 25 μL. The reaction mixtures were incubated at 37°C for 2 h and then at 70°C for 15 min to heat inactivate the enzymes. Restriction fragments were ligated to double-stranded adapters in a reaction containing 5 ρmol of EcoRI adapter and 50 ρmol of MseI adapter, 1 U T4 DNA ligase, and 0.2 μL of 10 × T4 ligase DNA buffer (Fermentas Life Sciences). The reaction mixture was incubated at 12°C for 17 h and heated at 65°C for 10 min to inactive the ligase (Desai et al., 1998).
The forward primer, a nonselective EcoRI primer (5′GACTGCGTACCAATTC3′), was labeled with a 6-carboxyfluorescein. The reverse primer, a nonlabeled MseI primer, had an extra selective base C at the 3′ (5′GATGAGTCCTGAGTAA
Amplification reactions were performed in a volume of 25 μL, containing 1 μL of ligated DNA, 20 μM of 6-FAM labeled EcoRI primer, 100 μM of MseI primer, 0.2 U Taq DNA polymerase (Photoneutria Biotecnologia e Serviços), 2.5 μL of 10 × polymerase chain reaction (PCR) buffer (10 mM Tris-HCl pH 8.0, 50 mM KCl), 1.5 mM MgCl2, and 0.2 mM of deoxynucleotides (GE Healthcare). Touchdown PCR was performed as follows: initial denaturation at 94°C for 2 min, followed by 9 cycles of denaturation at 94°C for 20 s, annealing at 66°C for 30 s, and extension at 72°C for 2 min. The annealing temperature for the first cycle was 66°C, and for the next eight cycles the temperature was decreased by 1°C at each cycle. The remaining 20 cycles were performed using a denaturation at 94°C for 20 s, annealing at 56°C for 30 s, and extension at 72°C for 2 min with a final extension at 60°C for 30 min. Amplifications were carried out in a Thermocycler (MiniCycler™; MJ Research). FAFLP products were separated on a MegaBace 1000 automatic sequencer (GE Healthcare). Each FAFLP reaction mixture was loaded with an internal size marker ET-550R (GE Healthcare), and the electrophoresis injection conditions were 3 kv at 150 s and 10 kv for 75 min. The AFs detected by the MegaBACE™ Fragment Profiler software were screened for detection of polymorphisms, verifying the presence or absence of fragments from 90 to 510 bp.
PFGE analysis
PFGE was performed according to the CDC PulseNet protocol (Ribot et al., 2006) using a CHEF DR II system (BioRad) at 6.0 V/cm with 0.5·TBE as running buffer. Electrophoresis conditions were an initial switch time of 2.2 s, a final switch time of 63.8 s, and a run time of 20 h. The CDC Salmonella Branderup isolate H9812 was used as the reference strain (Hunter et al., 2005). The gel was stained with ethidium bromide (1 μg/mL; Sigma) and photographed under UV-illumination. PFGE patterns were coded according to CDC PulseNet codes naming PFGE patterns (Swaminathan et al., 2001) with initials of Pontifícia Universidade Católica do Rio Grande do Sul (PUCRS); for example, “PUCRS.JEGX01.0001” would represent PFGE pattern 1 for Salmonella Enteritidis (“JEG”) cleaved with enzyme XbaI (“X01”).
Data analysis
Data were scored in a binary matrix format in Microsoft Excel (Microsoft Inc.) and exported to SPSS program (PASW Statistics 17) for the analysis of the similarity index between pairs of isolates. Cluster analysis was performed using neighbor-joining method based in Dice coefficient of similarity from which a dendrogram was generated. The isolates presenting >89.8% of similarity by FAFLP analysis and >84.6% of similarity by PFGE analysis were grouped in the same cluster. The discriminatory power was measured by the Simpson's index of diversity (D) (Hunter, 1990).
Results
The FAFLP analysis of 37 Salmonella Enteritidis strains and 4 strains of other Salmonella serovars consisted of 147 AFs in the size range of 61 to 510 bp. Only fragments with sizes between 90 and 510 bp were included in the final analysis based on a minimal sizing accuracy estimation of ∼0.5 bp. The number of AF differences between individual profiles ranged from 1 to 116. Nine FAFLP clusters were generated based in Dice similarity coefficient, which can be visualized in the neighbor-joining dendrogram (Fig. 1). Among the 40 profiles obtained, 96 AFs were shown to be polymorphic, resulting in a Simpson's index of diversity of 0.99.
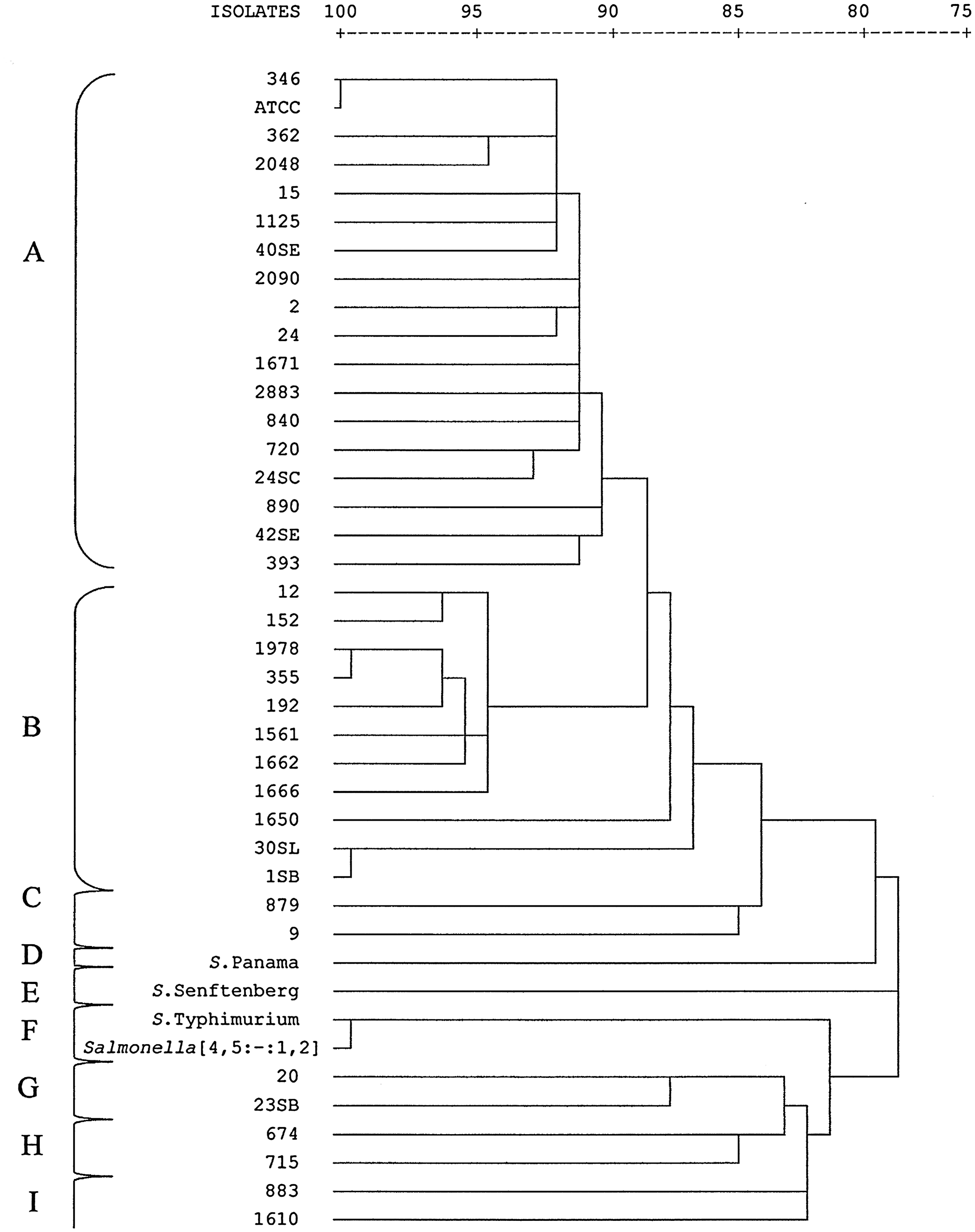
Dendrogram showing the genetic relationships between 37 isolates of Salmonella Enteritidis and 4 isolates of other Salmonella serovars. Similarity was determined by Dice coefficient and isolates clustered using neighbor-joining method on the basis of fluorescent amplified fragment length polymorphism profiles. Isolates exhibiting >89.8% similarity were grouped in the same cluster.
Cluster A was formed by the Salmonella Enteritidis ATCC 13076 and 18 strains isolated from humans, food, swine, broiler carcasses, and poultry-related samples and which belonged to phage types PT4, PT4a, PT6, PT6a, PT7, PT9, and PT11. Cluster B included 11 isolates originated from poultry, humans, food, broiler carcasses, and swine, with corresponding phage types PT4, PT4a, and one isolate of PT6a.
The strains 879 (PT4a) and 9 (PT4) isolated from broiler carcasses and poultry, respectively, shared one FAFLP cluster (cluster C) with 19 different AFs.
Isolates obtained from poultry (20) and swine (23SB) formed cluster G, presenting 92.8% of similarity and 12 divergent AFs. The 2 isolates of cluster H (674 and 715) from food and humans belong to phage type 4a and presented 91.2% of similarity, differing in 15 AFs one from each other. These two clusters showed around 88% similarity with each other.
The PFGE analysis discriminated 13 patterns (PUCRS.JEGX01.0001 to PUCRS.JEGX01.0013) that showed one to 41 different bands, resulting in a Simpson's index of diversity of 0.55. The pattern PUCRS.JEGX01.0001 was the most prevalent (31/40 isolates), being found in isolates from different sources and geographic areas (Egypt, Albania, and Zimbabwe). Eight patterns were represented by only one Salmonella Enteritidis isolate. The PFGE patterns grouped the isolates in seven clusters (Fig. 2), the majority of isolates being grouped in cluster I (≥88.9% of similarity).
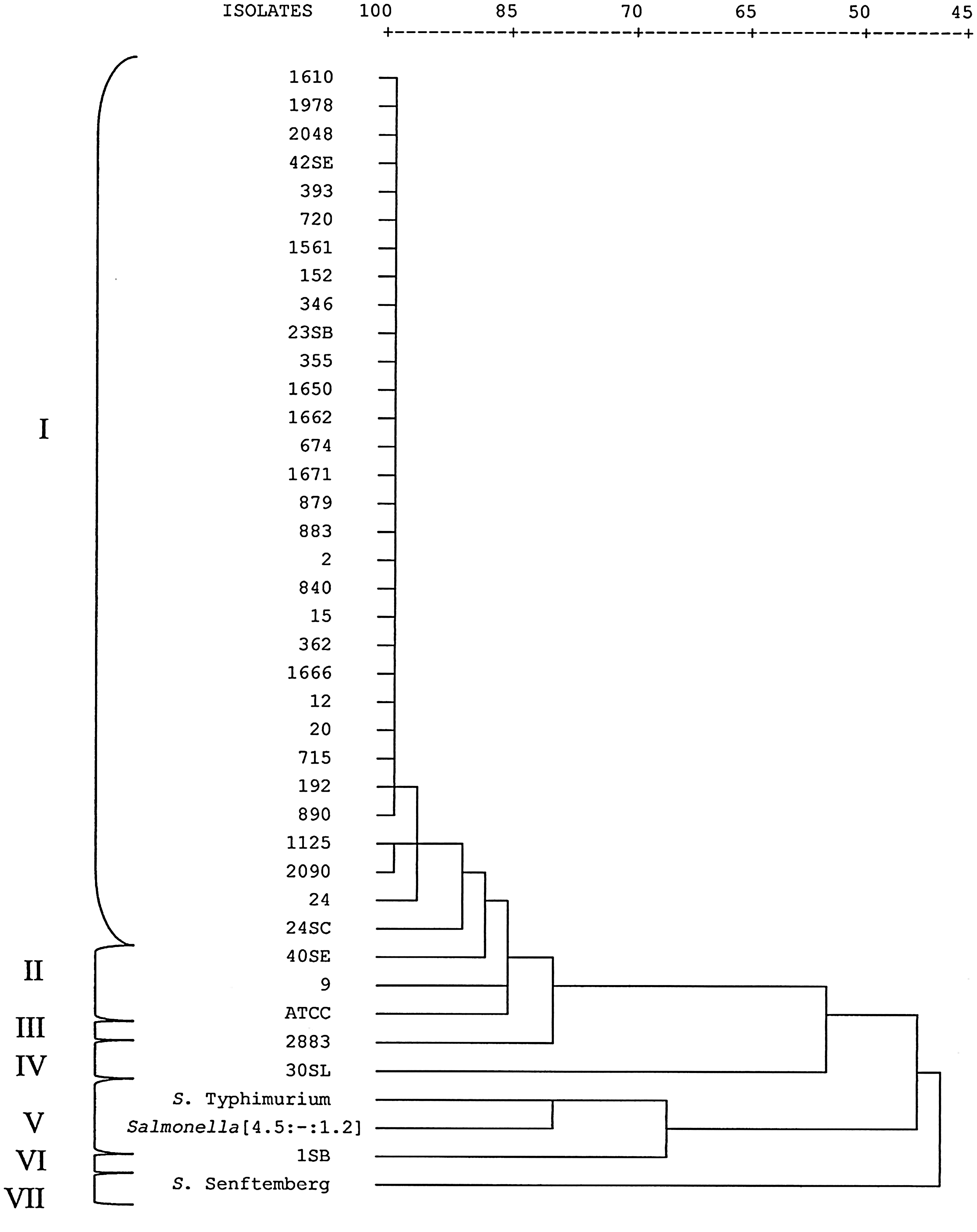
Dendrogram showing the genetic relationships between 37 isolates of Salmonella Enteritidis and 3 isolates of other Salmonella serovars. Similarity was determined by Dice coefficient and isolates clustered using neighbor-joining method on the basis of pulsed-field gel electrophoresis patterns. Isolates exhibiting >88.9% similarity were grouped in the same cluster.
Three strains of Salmonella Enteritidis from different geographic locations (Albania, Tanzania, and Italy) were clustered together (cluster A) by FAFLP. Egypt isolate constituted cluster B, and Zimbabwe isolate formed a cluster with the isolate 883 (cluster I). Differently, PFGE analysis grouped isolates from Italy, Egypt, Albania, and Zimbabwe in cluster I, whereas the isolate from Tanzania belonged to cluster III.
The outgroup, constituted by four isolates of distinct serovars, was discriminated by FAFLP and PFGE. The Salmonella Typhimurium and Salmonella [4,5:-:1,2] isolates were grouped in cluster F and cluster V by FAFLP and PFGE, respectively. The differences between these serovars were one AF (137 bp) in FAFLP and five bands in PFGE. Salmonella Senftenberg was discriminated from the other isolates and formed single clusters (cluster E and VII in FAFLP and PFGE, respectively). Salmonella Panama also formed a single cluster (cluster D) in FAFLP; however, this isolate could not be typed by PFGE using the standard CDC PulseNet protocol.
Discussion
FAFLP is based on the detection of genomic restriction fragments by PCR and has been shown to be efficient for the discrimination of subspecies of Salmonella (Scott et al., 2002) and between Salmonella serovars (Aarts et al., 1998; Lindstedt et al., 2000). This method was used in this study for the genotypic characterization of 37 isolates of Salmonella Enteritidis from different sources, as well as 4 other Salmonella serovar isolates, and compared with PFGE analysis, which is considered the “gold standard” method for Salmonella typing. The FAFLP analysis grouped the isolates in nine clusters and presented an index of discrimination of 0.99. These results were in agreement to those found by Desai et al. (2001) using the same primers and adapters, which obtained a discriminatory index of 0.98 to subtype Salmonella Enteritidis PT4. These authors found association between the FAFLP analysis and epidemiological data. Conversely, our data show that probably nonrelated isolates from different geographical areas were grouped in the same cluster, contrary to what the epidemiological data suggest. Similar results were obtained by PFGE, as Italy, Egypt, Albania, and Zimbabwe isolates were grouped together in cluster I. Indeed, clusters A and B harbor most isolates tested by FAFLP (43.9% and 26.8%, respectively), which were clustered in a single cluster (cluster I) in the PFGE analysis, grouping together isolates obtained from different sources and displaying various phage types, including type strain ATCC 13076. This result could be compared with data from Torpdahl et al. (2005), which by using three different molecular techniques (PFGE, multilocus sequence typing, and AFLP) did not find significant diversity between isolates of Salmonella enterica from human and veterinary sources, as well as with data by Lawson et al. (2004), which found 93% of outbreak-associated isolates and 82% of sporadic isolates of Salmonella Typhimurium in the same FAFLP profile. So, since Salmonella Enteritidis displays great genetic similarity (Olsen et al., 1994; Saeed et al., 2006; Boxrud et al., 2007), little discrimination between isolates could be expected, as the same isolates used in this study, when analyzed by rep-PCR or RAPD, showed high genetic homogeneity and low discriminatory power (Oliveira et al., 2007; Santos et al., 2008). However, the FAFLP results presented in this work showed a higher discrimination index than the other techniques, including the “gold standard” Salmonella typing method (PFGE), showing unique profiles for almost all isolates.
Although FAFLP clustering was not able to discriminate different phage types, it was already described that FAFLP was able to discriminate 25 PT reference strains of Salmonella Enteritidis into distinct profiles, six phage types being also analyzed in this study (Scott et al., 2001). Therefore, since phage typing still corresponds to an important tool for epidemiological typing of Salmonella Enteritidis, its association with molecular methods may greatly improve the discriminatory power between isolates of this serovar. Also, since isolates sharing the same phage type may present highly divergent FAFLP profiles and, therefore, present considerable genomic diversity, clustering analyses entirely based on single phenotypic markers should be taken with caution.
This study did not intend to subtype different Salmonella serovars by FAFLP, but four different serovars were included as outgroup. Among them, Salmonella Senftenberg and Salmonella Panama were grouped in distinct clusters, E and D, by FAFLP, respectively. Salmonella Typhimurium and Salmonella [4,5:-:1,2] isolates showed 99.5% similarity and were grouped in cluster F and cluster V by FAFLP and PFGE, respectively. Possibly, Salmonella [4,5:-:1,2] could be a monophasic variant originated from serovar Typhimurium (4,5,12:i:1,2), similar to what was suggested by others studies using FAFLP and other typing methods in relation to Salmonella [4,5,12:i:-] (Guerra et al., 2000; de la Torre et al., 2003; Alcaine et al., 2006; Soyer et al., 2009).
The context described here indicates that besides FAFLP presenting a high discriminatory power, this method also proves to be less laborious and to possess a higher resolving power than PFGE (Desai et al., 2001; Scott et al., 2002). Further, FAFLP is easily reproducible (Torpdahl and Ahrens, 2004) and requires only a small amount of DNA for analysis (Vos et al., 1995).
Conclusion
We conclude that FAFLP is a rapid method for epidemiological investigations of Salmonella outbreaks, presenting a high discriminatory power, although it was not able to distinguish in the clustering analysis putative epidemiologically nonrelated isolates.
Footnotes
Acknowledgments
The authors are grateful for the financial support provided by Conselho Nacional de Pesquisa (CNPq) and Laboratório de Medicina Veterinária Preventiva of Universidade Federal do Rio Grande do Sul for helpful assistance in the development of the PFGE protocol.
Disclosure Statement
No competing financial interests exist.