
Abstract
This study assessed the distribution of class 1 integrons in commensal bacteria isolated from agricultural and nonfarm environments, and the transferability of class 1 integrons to pathogenic bacteria. A total of 26 class 1 integron-positive isolates were detected in fecal samples from cattle operations and a city park, water samples from a beef ranch and city lakes, and soil, feed (unused), manure, and compost samples from a dairy farm. Antimicrobial susceptibility testing of class 1 integron-positive Enterobacteriaceae isolates from city locations displayed multi-resistance to 12–13 out of the 22 antibiotics tested, whereas class 1 integron-positive Enterobacteriaceae isolates from cattle operations only displayed tetracycline resistance. Most class 1 integrons had one gene cassette belonging to the aadA family that confers resistance to streptomycin and spectinomycin. One isolate from a dog fecal sample collected from a city dog park transferred its class 1 integron to a strain of Escherichia coli O157:H7 at a frequency of 10−7 transconjugants/donor by in vitro filter mating experiments under the stated laboratory conditions. Due to the numerous factors that may affect the transferability testing, further investigation using different methodologies may be helpful to reveal the transferability of the integrons from other isolates. The presence of class 1 integrons among diverse commensal bacteria from agricultural and nonfarm environments strengthens the possible role of environmental commensals in serving as reservoirs of antibiotic resistance genes.
Introduction
A
Studies have reported the presence of class 1 integrons in environmental commensal microorganisms (Rosser and Young, 1999; Nandi et al., 2004; Agerso and Sandvang, 2005; Gaze et al., 2005; Moura et al., 2007). Gram-positive bacteria such as Arthrobacter spp. and Corynebacterium spp. were found to be reservoirs for class 1 integrons in poultry litter (Nandi et al., 2004). Class 1 integrons and tetracycline resistance genes present on the same plasmids were found in Alcaligenes spp. and Pseudomonas spp. isolated from pigsties and manured soil (Agerso and Sandvang, 2005). Out of 3000 Gram-negative isolates examined from an aquatic environment, 3.6% of the isolates contained class 1 integrons (Rosser and Young, 1999). Both class 1 and 2 integrons were found in bacteria isolated from a slaughterhouse waste water treatment plant (Moura et al., 2007). Class 1 integrons were present in 3.8% of Gram-positive and Gram-negative isolates from environments polluted with quaternary ammonium compounds (Gaze et al., 2005). Commensal bacteria are present numerously and diversely in the environment, and those carrying class 1 integrons may serve as reservoirs of antibiotic resistance genes. In vitro conjugation experiments revealed that class 1 integrons present in environmental commensal bacteria could be transferred among commensals, and between commensal and pathogenic bacteria under antibiotic selective pressure (Agerso and Sandvang, 2005; Jiang et al., 2006).
It is generally believed that human and veterinary medicine and agricultural use of antibiotics exert selective pressures for presence of class 1 integrons in bacteria (Smith et al., 2007). Nevertheless, the presence of integrons in bacteria recovered from waste water and sediments in urban areas and even in environments relatively remote from human and agricultural activities has drawn attention to the impact of activities other than antibiotic use on the transfer of resistance genes between integron-associated settings (Roe et al., 2003; Elsaied et al., 2007; Gillings et al., 2008; Hardwick et al., 2008; Baker-Austin et al., 2009). Investigating class 1 integrons in various environments should help us to understand how environmental conditions may influence reservoirs of class 1 integrons in commensal bacteria and influence movement of class 1 integrons between commensals and pathogens. In the present study, we examined the distribution of class 1 integrons in commensal bacteria isolated from areas heavily influenced by cattle production, areas influenced by urban activities and indirectly influenced by agriculture, and areas not influenced by agriculture. Class 1 integron-positive commensal bacteria were provisionally identified and compared for gene cassette diversity through sequence analysis. Class 1 integron-positive commensal bacteria were also tested for their conjugative transferability to pathogenic bacteria. The resulting data should be helpful in understanding the presence and function of class 1 integrons in commensal bacteria under varying environmental pressures, and in predicting their role in spreading antibiotic resistance.
Materials and Methods
Isolation of commensal bacterial strains
Soil, fecal floor, and water samples were collected from cattle operations (144 samples) and nonfarm environments (144 samples), the latter including the city of Fort Collins (96 samples) and the Rocky Mountain National Park (48 samples), in Colorado. The collection of soil, fecal floor, and water samples followed the methods described by Srinivasan et al. (2008), Yang et al. (2006), and Esiobu et al. (2002), respectively. In addition, a total of 42 samples of manure, compost, and unused feed and bedding were collected from cattle operations following the method described by Nandi et al. (2004). Cattle operations, including a beef ranch and a dairy farm, represented areas heavily influenced by agricultural activities. City locations, including city parks, hospital areas, and water reclamation facilities, represented areas influenced by urban activities and indirectly influenced by agriculture (Pei et al., 2006). The Rocky Mountain National Park represented an area not influenced by agriculture.
Each sample (10 g or 10 mL) was serially diluted in 0.1% buffered peptone water (Difco, Becton Dickinson, Sparks, MD) and spread-plated onto tryptic soy agar (Difco) supplemented with 400 μg/mL delvocid salt (DSM Food Specialties, Delft, The Netherlands) (TSA/DS) and tetracycline (8 μg/mL; Sigma-Aldrich, St. Louis, MO) (TSA/DS/TET), and TSA/DS supplemented with ceftiofur (4 μg/mL; Sigma-Aldrich) (TSA/DS/CEF). Delvocid salt is a fungicide containing 50% (w/w) natamycin (Ghosh and LaPara, 2007). On the basis of breakpoint minimum inhibitory concentrations (MICs) established by the Clinical Laboratory Standard Institute (CLSI), the breakpoint MIC of ceftiofur for Salmonella and Escherichia coli is ≥8 μg/mL, and the breakpoint MIC of tetracycline for organisms other than streptococci is ≥16 μg/mL (CLSI, 2002). TSA/DS/CEF and TSA/DS/TET were used in this study for enumeration of bacterial populations with a MIC of ≥8 μg/mL for ceftiofur and with a MIC of ≥16 μg/mL for tetracycline, respectively.
Colonies with distinct morphologies (one of each morphology) were picked from the plates supplemented with antibiotics after incubation at 25°C for 72 h. For each sample, the numbers of colonies picked were based on their morphological diversities. All the colonies were streaked twice on TSA/DS/TET or TSA/DS/CEF plates for purity and were then stored at −80°C in tryptic soy broth (Difco) supplemented with 20% glycerol. Each isolate was activated by two successive transfers in tryptic soy broth at 30°C for 24 h before use for the following tests.
Polymerase chain reaction detection of class 1 integrons in isolates resistant to tetracycline or ceftiofur
Total bacterial DNA of each isolate was extracted by boiling in water for 10 min. The extracted DNA from each isolate was subjected to polymerase chain reaction (PCR) screening for class 1 integrons by using the primer set of 5′-CS (5′-GGC ATC CAA GCA GCA AG-3′) and 3′-CS (5′-AAG CAG ACT TGA CCT GA-3′) (Jiang et al., 2006). The PCR was carried out in a total volume of 50 μL, containing 3 μL of bacterial DNA template, 25 μL of iQ™SYBR Green supermix (Bio-Rad, Hercules, CA), and 2 μL of each primer (25 μM). The amplification conditions were 95°C for 10 min, 40 cycles of 95°C for 1 min, 54°C for 1 min, and 72°C for 2 min. Distilled water was used as a negative control, and E. coli O11:NM CVM 1884, kindly provided by Dr. Shaohua Zhao (U.S. Food and Drug Administration, Laurel, MD), was used as a positive control (Zhao et al., 2001). The reaction products were observed by electrophoresis in 0.9% agarose gels stained with ethidium bromide under ultraviolet light (312 nm).
DNA sequencing for antibiotic resistance gene cassettes within class 1 integrons
Purified PCR products were presequenced with an ABI 3130 Genetic Analyzer (Applied Biosystems Inc., Foster City, CA) at the Proteomics and Metabolomics Facility of Colorado State University by using the above-described primer sets of 5′-CS and 3′-CS. DNA sequences were compared for homology with the GenBank sequence database by using the BLAST network service available from the National Center for Biotechnology Information. PCR products with confirmed conservative 5′- and 3′-regions were sent to a commercial Primer Walking service provided by Eton Bioscience, Inc. (San Diego, CA) for the determination of antibiotic resistance gene cassettes between the conserved regions.
Provisional identification of class 1 integron-positive isolates
Integron-positive isolates were subjected to PCR for 16S rRNA genes using the primer set of 16SF (GGT GAA TAC GTT CYC GG) and 16SR (GGW TAC CTT GTT ACG ACT T) (Nandi et al., 2004). The PCR was carried out in a total volume of 25 μL, containing 3 μL of bacterial DNA template, 12.5 μL of iQ™SYBR Green supermix (Bio-Rad), and 1 μL of each primer (25 μM). The amplification conditions were 95°C for 10 min, and 40 cycles of 94°C for 30 sec, 50°C for 30 sec, and 72°C for 1 min. The PCR-generated DNA fragments were then purified using Ultrafree-DA centrifugal filter units (Millipore, Bedford, MA). Purified PCR products were sequenced with the ABI 3130 Genetic Analyzer at the Proteomics and Metabolomics Facility (Colorado State University) by using the above-described primers. DNA sequences were analyzed by searching the GenBank database of the National Center for Biotechnology Information via the BLAST network service. The resulting gene sequences were aligned in ClustalW (
Antimicrobial susceptibility testing
Antimicrobial MICs of the class 1 integron-positive isolates were determined with the Sensititre MIC Susceptibility system (Trek Diagnostic Systems, Westlake, OH) according to the manufacturer's instructions (Zhao et al., 2001). The results were interpreted according to the CLSI guidelines for broth microdilution methods (CLSI, 2000). Gram-positive isolates were tested for the following 21 antibiotics (breakpoints of each of the tested antibiotics are given in parentheses; μg/mL): ampicillin (16), chloramphenicol (32), ciprofloxacin (4), clindamycin (4), daptomycin (8), erythromycin (8), gemifloxacin (0.5), gentamicin (16), linezolid (8), moxifloxacin (8), nitrofurantoin (16), oxacillin + 2% NaCl (0.5), penicillin (16), quinupristin/dalfopristin (4), rifampin (4), streptomycin (1000), telithromycin (4), tetracycline (16), tigecycline (0.5), trimethoprim/sulfamethoxazole (4/76), and vancomycin (32). Gram-negative isolates were tested for the following 22 antibiotics (breakpoints of each of the tested antibiotics are given in parentheses; μg/mL): amikacin (64), ampicillin (32), ampicillin/sulbactam 2:1 (32/16), aztreonam (32), cefazolin (32), cefepime (32), cefoxitin (32), cefpodoxime (8), ceftazidime (32), ceftriaxone (64), cefuroxime (32), cephalothin (32), ciprofloxacin (4), ertapenem (8), gentamicin (16), meropenem (16), piperacillin/tazobactam (128/4), tetracycline (16), ticarcillin/clavulanic acid (128/2), tigecycline (8), tobramycin (16), and trimethoprim/sulfamethoxazole (4/76). E. coli ATCC 25922 and Enterococcus faecalis ATCC 29212 were used as controls in antimicrobial MIC determinations.
Conjugation studies
The class 1 integron-positive isolates were used as donor strains. Green fluorescent protein (GFP)-expressing E. coli O157:H7 ATCC 43895/ISEHGFP (Noah et al., 2005); Salmonella Typhimurium AM 04423, kindly provided by Dr. Jean M. Whichard (Centers for Disease Control and Prevention, Atlanta, GA) (Whichard et al., 2005); and E. faecalis ATCC 29212 were used as recipient strains. Conjugation was performed by mating a donor strain with a recipient strain at the ratio of 0.1:0.1 mL (109 CFU/mL) on a 0.20 μm sterile syringe filter (Nalge Nunc, Rochester, NY) over TSA (Difco) plates (Jiang et al., 2006). After incubation at 35°C for 18 h, the filter was carefully removed with sterile forceps and transferred into a 50-mL sterile tube (Falcon, BD Biosciences, Franklin Lakes, NJ) containing 20 mL of 0.1% buffered peptone water. The tubes were vortexed vigorously for 1 min, serially diluted, and plated. Cell numbers of recipients of E. coli O157:H7 ATCC 43895/ISEHGFP, Salmonella Typhimurium AM 04423, and E. faecalis ATCC 29212 were determined on TSA supplemented with 4 μg/mL gentamicin (Sigma-Aldrich) (TSA/GEN), XLT4 agar (Difco), and TSA/GEN, respectively. The selective media for transconjugates are listed in Table 1. The transconjugates with E. coli O157:H7 ATCC 43895/ISEHGFP as recipients were confirmed both by observation of green fluorescence under ultraviolet light and PCR amplification of class 1 integron genes as described above.
The detection limit was 10−9.
Resistant to gentamicin out of the 22 antibiotics tested in Table 5.
Susceptible to all of the 22 antibiotics tested in Table 5.
MIC ranges for E. faecalis ATCC 29212 are available in CLSI (2002).
Isolate number.
Provisional identification of isolate to genus or species level based on 16S rRNA gene sequencing results (Table 3).
TSA/GEN/TET: TSA supplemented with gentamicin (4 μg/mL) and tetracycline (8 μg/mL).
SMAC-CT/TET: SMAC-CT supplemented with tetracycline (8 μg/mL).
XLT4/TET: XLT4 supplemented with tetracycline (8 μg/mL).
TSA/GEN/ERY: TSA supplemented with gentamicin (4 μg/mL) and erythromycin (4 μg/mL).
SMAC-CT/SXT: SMAC-CT supplemented with trimethoprim/sulfamethoxazole (2/38 μg/mL).
TSA/GEN/AMP: TSA supplemented with gentamicin (4 μg/mL) and ampicillin (8 μg/mL).
XLT4/SXT: XLT4 supplemented with trimethoprim/sulfamethoxazole (2/38 μg/mL).
MICs, minimum inhibitory concentrations.
Results
PCR detection of class 1 integrons in commensal bacteria resistant to tetracycline or ceftiofur
A total of 1870 isolates resistant to tetracycline or ceftiofur were obtained, including 538 isolates from the beef ranch, 573 isolates from the dairy farm, 543 isolates from the city locations, and 216 isolates from the national park (Table 2). A total of 26 isolates from cattle operations and city locations were found to be class 1 integron positive. Class 1 integrons were not found in any of the isolates from the Rocky Mountain National Park. The percentages of class 1 integron-positive isolates were 1.86% (10/538), 1.75% (10/573), and 1.10% (6/543) for the beef ranch, dairy farm, and city locations, respectively. These class 1 integron-positive isolates were detected from fecal floor samples from cattle operations and the city dog park, water samples from the beef ranch and city lakes, and soil, feed (unused), manure, and compost samples from the dairy farm (Table 2).
Number of class 1 integron-positive isolates/number of bacterial isolates with tetracycline MIC ≥16 μg/mL or ceftiofur MIC ≥8 μg/mL from this type of sample.
Three from animal drinking water and one from waste water.
All from lakes in city.
Two from nursing calves and four from young heifers.
One from nursing calf, one from young heifer, and one from stocker cattle.
From pet dogs.
Samples were not collected from this location.
DNA sequencing of antibiotic resistance gene cassettes of class 1 integrons
Most of the class 1 integron-positive commensal bacteria had class 1 integrons carrying one gene cassette of ∼1100 bp in size. The gene cassettes within most of the integrons belonged to the aadA family of genes conferring resistance to streptomycin and spectinomycin (Table 3). Three and one class 1 integron-positive isolates from the city locations carried aadA1 and aadA2 gene cassettes, respectively. For class 1 integron-positive isolates from cattle operations, five isolates carried aadA1; two isolates carried aadA5; one isolate carried aadA7; and nine isolates carried aadA9 gene cassettes. In addition, one manure isolate from the dairy farm (336-C-3) was found to carry a dfr16 gene cassette encoding resistance to trimethoprim. Only two isolates were found to have class 1 integrons with more than one gene cassette. Specifically, one fecal isolate (199-T-3) from the beef ranch carried two gene cassettes (aadA1 and dfr1), and one pet-dog-fecal isolate (221-C-1) from the city dog park contained three gene cassettes of dfrA12, orfF (encoding a putative protein), and aadA2 (Table 3).
To all three recipients of E. coli O157:H7, Salmonella Typhimurium, and E. faecalis.
aadA1, aadA2, aadA5, aadA7, and aadA9: encoding aminoglycoside-3′′-adenylyltransferases, which confer resistance to streptomycin and spectinomycin.
dfrA12, dfr16, and dfr1: encoding dihydrofolate reductases, which confer resistance to trimethoprim.
orfF: encoding putative protein.
To the recipient of E. coli O157:H7; the transfer frequencies for Salmonella Typhimurium and E. faecalis were <10−9.
Provisional identification of class 1 integron-positive isolates
Results of the provisional identification of class 1 integron-positive isolates, through 16S rRNA gene sequencing, are shown in Table 3. A neighbor-joining distance tree (Fig. 1) was generated based on 16S rRNA gene sequence data of the 26 class 1 integron-positive isolates to explore the genetic homogeny between isolates (Gillings et al., 2008). The neighbor-joining tree revealed that the class 1 integron-positive isolates from cattle operations belonged to alpha-, beta-, and gamma-proteobacteria, Actinobacteria, and Bacilli (Fig. 1). In contrast, except for isolate 160-T-2, which belonged to beta-proteobacteria, all the class 1 integron-positive isolates from city locations belonged to gamma-proteobacteria.
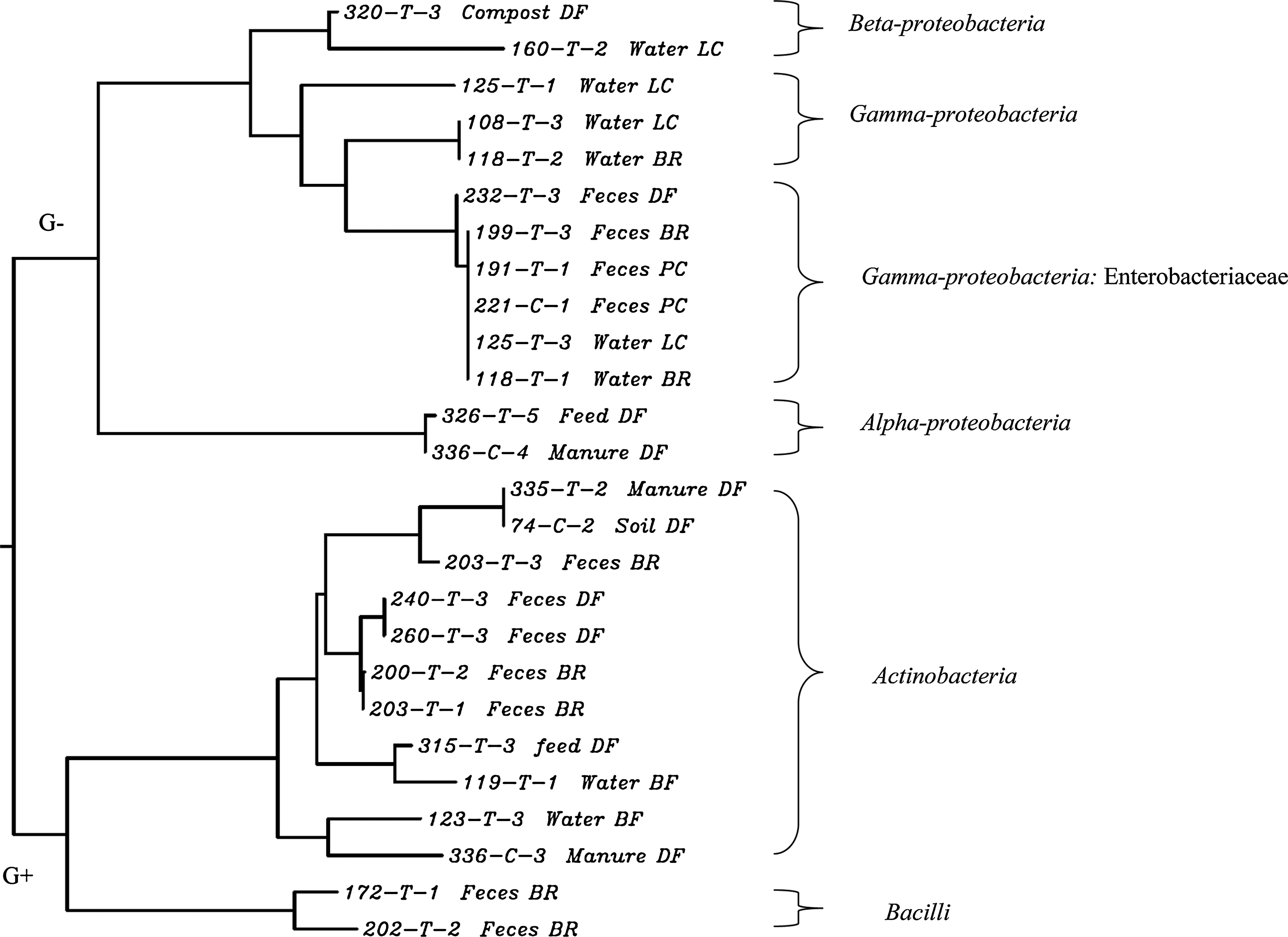
Neighbor-joining distance tree based on 16S rRNA gene sequences of class 1 integron-positive isolates. 320-T-3 was identified as Alcaligenes; 160-T-2 as Delftia; 125-T-1 as Stenotrophomonas; 191-T-1 as E. coli; 326-T-5 as Brevundimonas; 335-T-2 and 74-C-2 as Arthrobacter; 240-T-3, 260-T-3, and 203-T-1 as Dietzia; 315-T-3 and 119-T-1 as Corynebacterium; 336-C-3 as Microbacterium; 123-T-3 as Leucobacter; 172-T-1 and 202-T-2 as Bacillus. BR, beef ranch; DF, dairy farm; LC, lakes in city; PC, park in city.
Antimicrobial susceptibility testing
MIC results for Gram-positive and Gram-negative isolates against different antibiotics, which are clinically important for the treatment of corresponding infections, are presented in Tables 4 and 5, respectively. Since there are no established breakpoints for all commensal bacteria, while breakpoints for Enterobacteriaceae have been well established, only Enterobacteriaceae isolates were compared for their resistance patterns. Enterobacteriaceae isolates from the city locations (125-T-3, 221-C-1, and 191-T-1) displayed multi-resistance to 12–13 of the 22 antibiotics tested, while class 1 integron-positive Enterobacteriaceae isolates from cattle operations (118-T-1, 199-T-3, and 232-T-3) only displayed resistance to tetracycline (Table 6).
AMP, ampicillin; BR, beef ranch; CHL, chloramphenicol; CIP, ciprofloxacin; CLI, clindamycin; DAP, daptomycin; DF, dairy farm; ERY, erythromycin; GEM, gemifloxacin; GEN, gentamicin; LZD, linezolid; MXF, moxifloxacin; NIT, nitrofurantoin; OXA+, oxacillin +2% NaCl; PEN, penicillin; RIF, rifampin; STR, streptomycin; SXT, trimethoprim/sulfamethoxazole; SYN, quinupristin/dalfopristin; TEL, telithromycin; TET, tetracycline; TGC, tigecycline; VAN, vancomycin.
AMI, amikacin; A/S2, ampicillin/sulbactam 2:1; AXO, ceftriaxone; AZT, aztreonam; CEP, cephalothin; DPC, dog park in city; ETP, ertapenem; FAZ, cefazolin; FEP, cefepime; FOX, cefoxitin; FUR, cefuroxime; MERO, meropenem; POD, cefpodoxime; P/T4, piperacillin/tazobactam; TAZ, ceftazidime; TIM2, ticarcillin/clavulanic acid; TOB, tobramycin; LC, lake in city.
S: Considered as susceptible, when the MIC was less than breakpoint according to CLSI (2000) guidelines for interpretation of MIC data for Enterobacteriaceae.
R: Considered as resistant, when the MIC was greater than or equal to breakpoint according to CLSI (2000) guidelines for interpretation of MIC data for Enterobacteriaceae.
Conjugation studies
In this study, we tested the transfer of class 1 integrons from commensal bacteria to E. coli O157:H7, Salmonella Typhimurium, and E. faecalis. Only one integron-positive fecal isolate from the city dog park (221-C-1) transferred its class 1 integron to E. coli O157:H7 at a frequency of 10−7 transconjugants/donor (Table 3). The remaining transfer frequencies of class 1 integrons from commensal bacteria to pathogenic bacteria were less than the detection limit of 10−9 (Table 3).
Discussion
The significance of detection of class 1 integrons in the present study is that it targeted all the culturable commensal bacteria present in various environments with a MIC of ≥16 μg/mL for tetracycline or a MIC of ≥8 μg/mL for ceftiofur. The definition of commensal bacteria used in this study is that provided by Marshall et al. (2009): “bacterial strains deemed not actively responsible for a pathogenic process and derived from humans, animals, or plants, or recovered from environmental sources such as air, water, soil, sludge, etc.” Tetracycline and ceftiofur were used in this study to represent antibiotics that have been used for treating cattle disease for a long time and from a recent time (since 1988), respectively. In a study by Agerso and Sandvang (2005), the tetracycline resistance genes, tet(A) and tet(C) encoding the efflux pump, were found to be present with class 1 integrons on the same plasmids. Further, Jiang et al. (2006) reported that the ceftiofur resistance gene, blaCMY-2 , was cotransferred with class 1 integrons to recipients in conjugation studies. The transconjugants in the latter study obtained resistance to other antibiotics, such as chloramphenicol, streptomycin, kanamycin, gentamicin, ampicillin, and tetracycline from the donor strains as well (Jiang et al., 2006). Therefore, the use of these two antibiotics in food animals may result in the selection of integrons (Agerso and Sandvang, 2005; Jiang et al., 2006).
Since the genera of the bacteria were not known before PCR detection of class 1 integrons, a set of primers widely used for detection of class 1 integrons among commensal E. coli and Salmonella strains was applied to all of the 1870 isolates (Zhao et al., 2001). This primer set resulted in multiple bands for some bacterial isolates (data not shown). The amplicon sizes of class 1 integrons depend on the number of gene cassettes inserted between conserved regions. If no resistance gene cassette is inserted, the expected size is 153 bp (Gebreyes and Thakur, 2005). Class 1 integrons carrying one antibiotic resistance gene or one unknown protein are most times >750 bp (Zhao et al., 2001; Chen et al., 2004; Gebreyes and Thakur, 2005). In the present study, any bands >500 bp were purified and sequenced to avoid under-estimation of class 1 integrons containing gene cassettes for antibiotic resistance.
Previous studies have reported the detection of class 1 integrons in various locations, including human and animal clinical settings, agricultural environments, urban and pristine areas, food animals, pets, and wild animals (Rosser and Young, 1999; Nandi et al., 2004; Agerso and Sandvang, 2005; Gaze et al., 2005; Moura et al., 2007; Hardwick et al., 2008; Baker-Austin et al., 2009). In this study, 216, 538, 573, and 543 isolates from the Rocky Maintain National Park, beef ranch, dairy farm, and city locations, respectively, were screened for the presence of class 1 integrons by PCR. Class 1 integrons were detected in both cattle operations and in the city locations, but not in samples collected from the national park (Table 2). The fact that integrons were not detected in the national park area may be attributed to the absence of human and agricultural activity. However, the absence of class 1 integrons in the national park area needs to be confirmed because only culturable bacteria were screened for class 1 integrons in our study. Further studies with larger sample sizes, including nonculturable bacteria and using antibiotics other than tetracycline and ceftiofur for initial selection, may give a more accurate estimate of the presence of integrons in the national park.
The presence of class 1 integrons in cattle operations is not surprising as numerous studies have reported it previously (Douris et al., 2007; Scott et al., 2009). The detection of class 1 integrons in unused feed in cattle operations indicates that the feed may have been contaminated with antibiotic-resistant bacteria before arriving at the cattle operation, and may serve as a source of contamination of cattle. Of particular concern is the presence of integron-positive bacteria in the city locations. Unlike cattle operations, the city lakes and dog parks do not have direct application of antibiotics. Nevertheless, they are exposed to humans and pets that have had or are having antibiotic treatments, and are exposed to various urban activities. The presence of class 1 integrons in isolates from pet dog feces indicated that human pets could be a source and/or means of transmission of antibiotic resistance in urban environments and humans.
All class 1 integron-positive isolates from city locations except for one isolate were taxonomically close to gamma-proteobacteria, whereas the isolates from cattle operations displayed diverse taxonomic relationships, which is in agreement with the properties of class 1 integron-positive isolates recovered from diverse habitats (Elsaied et al., 2007; Gillings et al., 2008; Hardwick et al., 2008; Baker-Austin et al., 2009). All six class 1 integron-positive isolates from the city locations were Gram-negative, whereas the majority (13 out of 20) of class 1 integron-positive isolates from the cattle operations were Gram-positive (Table 3). On the basis of previous reports, Gram-negative bacteria are much more of a concern than Gram-positive bacteria in the distribution of antibiotic resistance (Stokes et al., 2006; Gillings et al., 2008). Sequencing data of 16S rRNA genes allowed provisional identification of some isolates to genus level, but not for all the isolates because the discriminatory power of 16S rRNA genes varies between genera (Janda and Abbott, 2007). Since there is lack of a harmonious set of guidelines for the use of 16S rRNA gene sequencing for definitive microbial identifications (Janda and Abbott, 2007), sequencing results were used as reference identifications for integron-positive commensal bacterial isolates in this study. Other tests, for example, conventional biochemical or rapid ID tests, may be needed for complete identification of the isolates in the future.
Sequence analysis revealed that the majority of class 1 integrons carried one antibiotic resistance gene cassette belonging to the aadA family (Table 3). By comparing the data related to resistance gene cassettes and the antibiotic resistance phenotype of Enterobacteriaceae isolates, it is interesting to find that not all the antibiotic resistance phenotypes were encoded by class 1 integrons, and not all of the isolates displayed the resistance phenotype corresponding to the resistance gene cassettes (Tables 3, 5, and 6). Bacteria containing class 1 integrons but not displaying resistance phenotypes have been reported previously. Specifically, integron-positive isolates carrying the aadA gene cassette but that were not resistant to streptomycin have been reported in several studies (Zhao et al., 2001; Roe et al., 2003; Pan et al., 2006; Chang et al., 2007). A class 1 integron-positive human commensal, Enterobacter cloacae, was sensitive to all antibiotics tested except some members of the β-lactam family (Labbate et al., 2008).
Transmission of class 1 integrons to the selected recipients of E. coli O157:H7, Salmonella Typhimurium or E. faecalis was not observed (<10−9) for all the tested isolates, except for isolate 221-C-1. Pairing of each commensal bacterial isolate with closely related bacteria as recipients for conjugation experiments, and determination of integron location, either on plasmids or on the chromosome, is needed for further confirmation of their transferability. However, as indicated, isolate 221-C-1 did show its capability of transferring its class 1 integron to pathogenic E. coli O157:H7 at a frequency of 10−7. This transfer frequency is in agreement with previously reported class 1 integron frequencies ranging from 10−5 to 10−8 (Jiang et al., 2006; Nagachinta and Chen, 2008). This fecal isolate carrying the transferable class 1 integron was from a city dog park, was resistant to 13 of the 22 tested antibiotics, and had three gene cassettes within its class 1 integron. This class 1 integron-positive bacterium may play a role in the spread of antibiotic resistance genes, in addition to serving as a reservoir of antibiotic resistance genes.
Conclusion
In the present study, class 1 integrons in commensal bacteria from cattle operations and city locations were characterized and tested for their transferability. In addition to cattle operations, class 1 integron-positive commensal bacteria were found in urban environments with human and pet animal activities. One isolate from a dog fecal sample collected from a city dog park transferred its class 1 integron to a strain of E. coli O157:H7 at a frequency of 10−7. The presence of class 1 integrons among diverse commensal bacteria from agricultural and nonfarm environments strengthens the possible role of environmental commensals in serving as reservoirs of antibiotic resistance genes. It should be noted, however, that this study was based on culturable bacterial populations and using tetracycline and ceftiofur for initial selection. It will require more studies of microbial communities in various environments for class 1 integrons to confirm and expand the findings.
Footnotes
Acknowledgment
This work was funded by The Beef Checkoff and by the Colorado State University Agricultural Experiment Station.
Disclosure Statement
No competing financial interests exist.