
Abstract
Pomegranate juice (PJ) has gained popularity because of its associated antioxidant, antimicrobial, anticancer, and anti-inflammatory properties. However, its effects against epidemiologically significant foodborne viruses have not been investigated. In the absence of culturable human noroviruses, feline calicivirus (FCV-F9), murine norovirus (MNV-1), and MS2 (ssRNA) bacteriophage were used as foodborne viral surrogates. The aim of this research was to study the effects of PJ and pomegranate polyphenols (PP) on foodborne viral infectivity. Viruses at high (∼7 log10 PFU/mL) or low (∼5 log10 PFU/mL) titers were mixed with equal volumes of PJ, 8, 16, and 32 mg/mL of PP, or water (control) and incubated for 1 h at room temperature. Viral infectivity after treatments was evaluated using standardized plaque assays. PJ decreased the titer of FCV-F9, MNV-1, and MS2 by 2.56, 1.32, and 0.32 log10 PFU/mL, respectively, for low titers and 1.20, 0.06, and 0.63 log10 PFU/mL, respectively, for high titers. Interestingly, FCV-F9 was undetectable after exposure to the three tested PP solutions using both low and high titers. MNV-1 at low initial titers was reduced by 1.30, 2.11, and 3.61 log10 PFU/mL and at high initial titers by 1.56, 1.48, and 1.54 log10 PFU/mL with 4, 8, and 16 mg/mL of PP treatment, respectively. MS2 at low initial titers was reduced by 0.41, 0.45, and 0.93 log10 PFU/mL and at high initial titers by 0.32, 0.41, and 0.72 log10 PFU/mL after 4, 8, and 16 mg/mL of PP treatment, respectively. PJ and PP resulted in titer reductions of foodborne virus surrogates after 1 h exposure, showing promise for use in hurdle technologies and/or for therapeutic or preventive use. To suggest the use of PJ and PP as natural remedies for foodborne viral illness prevention, their mechanism of action against viral infectivity needs to be further investigated.
Introduction
Human noroviruses remain the leading cause of viral gastroenteritis outbreaks worldwide (Widdowson et al., 2004; Turcios et al., 2006; Siebenga et al., 2009). They are nonenveloped round particles with a diameter of 27–40 nm and have a low infectious dose of 10–100 viral particles. Because of their nonenveloped nature, noroviruses are very stable and resistant to environmental degradation and chemical inactivation (Cheesbrough et al., 2000; Kuusi et al., 2002). Person-to-person, foodborne, and water-borne transmissions of noroviruses have been documented (Fankhauser et al., 2002). The symptoms associated with norovirus infections are nausea, vomiting, diarrhea, abdominal pain, and low-grade fever. Typically, norovirus infections are self-limiting and usually cause no complications, but the newly emergent strains have become highly virulent and found to be life threatening, especially to the elderly and immunocompromised (Siebenga et al., 2009). No effective medication is yet available, though symptoms are known to be somewhat alleviated with over-the-counter antidiarrheal agents such as bismuth subsalicylate. It is therefore intriguing to explore the effectiveness of natural remedies such as PJ and pomegranate extracts for the treatment or prevention of foodborne norovirus infections.
Human noroviruses cannot be used to evaluate the antiviral effect of PJ or PP, as these viruses cannot be grown in cell culture (Duizer et al., 2004). The lack of infectivity assays for human noroviruses necessitates the use of surrogates such as feline calicivirus (FCV) (Steinmann, 2004), bacteriophage MS2 (Dawson et al., 2005), and murine norovirus (MNV-1) (Wobus et al., 2004) to determine antiviral effects. The capsid structure, genomic organization, and replication cycle of MNV-1 (a genogroup V norovirus of the Caliciviridae family) are very similar to those of human noroviruses (Sosnovtsev et al., 2006; Wobus et al., 2006). Therefore, MNV-1 is considered to be the most appropriate surrogate for human noroviruses by some researchers.
The purpose of the present study was to evaluate the antiviral activity of PJ and PP (POM™—an extract of polyphenols from fresh pomegranate fruit) against foodborne virus surrogates FCV-F9, MNV-1, and MS2 bacteriophage. The time dependence of PJ and PP on the infectivity of MNV-1 was also studied.
Materials and Methods
Viruses, bacterial hosts, and cell lines
Crandell Reese Feline Kidney (CRFK) cells, FCV-F9, bacteriophage MS2, and E. coli B-15597 cells were purchased from ATCC (Manassas, VA). RAW 264.7 cells were obtained from the University of Tennessee at Knoxville. MNV-1 was provided as a gift by Dr. Skip Virgin (Washington University, St. Louis, MO).
Propagation of viruses
Both CRFK and RAW 264.7 cell lines were maintained at 37°C in a water-jacketed incubator supplemented with 5% CO2. Viral stocks were prepared by inoculation of FCV-F9 and MNV-1 stock to monolayers of CRFK and RAW 264.7 cells, respectively, and cultured until >90% cell lysis. MS2 coliphage was propagated in E. coli B-15597 host in 3% trypticase soy broth containing 0.1% glucose, 2 mM CaCl2, and 10 μg/mL thiamine at 37°C for 18 h. The viruses were harvested by centrifugation and filtration as described earlier (D'Souza and Su, 2010; Su et al., 2009).
Antiviral effects of PJ and PP solutions
PJ (Pure Pomegranate, Northland, Northland Products, LLC, Port Washington, NY), was sterilized by filtration through 0.22-μm filters. PP was obtained as a gift from POM Wonderful (Los Angeles, CA). Three steps are involved in PJ production: (1) pressing of pomegranates to obtain juice; (2) centrifugation of obtained fruit juice through a centrifuging separator to remove suspended material; and (3) clarification of juice by heating the clear juice to inactivate enzymes and destroy any vegetative microorganisms. PP were reportedly produced by aqueous extraction of fruit residue after pressing for juice and removal of seeds and then solid-phase extraction with an aqueous ethanol solution to produce a powder containing >90% of polyphenols.
To prepare PP solution, the PP were first dissolved in methanol and sterile deionized distilled water, filter sterilized (0.2-μm filter), and further diluted aseptically to 8, 16, and 32 mg/mL in sterile deionized distilled water. PJ or PP was mixed with equal volume of each virus to obtain titers of ∼7 or ∼5 log10 PFU/mL and incubated at room temperature for 1 h. For the time-dependence study, PJ or 8 mg/mL PP solution was mixed with equal volumes of FCV-F9 or MNV-1 and incubated for 0, 10, 20, 30, 45, and 60 min, respectively. Individual viruses mixed with sterile deionized distilled water were used as the untreated controls. After incubation, treatments were neutralized by 10-fold serial dilution of virus in Dulbecco's modified Eagle's medium (DMEM-F12) containing fetal bovine serum (FBS).
Cytotoxicity assays were carried out by adding 10-fold dilutions of PJ or PP samples in DMEM containing FBS to confluent CRFK and RAW 264.7 cell lines and incubated for up to 3 days. Cells treated with PJ or PP were also overlaid with DMEM containing 0.75% agarose, with continued incubation for 2 and 3 days for CRFK and RAW 264.7 cells, respectively, and then stained with neutral red solution as described in detail below. Cytopathic effects were determined by both visual inspection under an optical light microscope and neutral red staining.
All treatments were performed at least twice in independent experiments and plaque assays for evaluating the infectivity of the viruses were carried out in duplicates as described below.
Infectious plaque assays
The MNV-1 plaque assay previously described by Wobus et al. (2004) was followed with minor changes. Briefly, confluent RAW 264.7 cells were inoculated with 10-fold serial dilutions of viral samples in DMEM containing 10% FBS. Viruses were adsorbed for 2 h at 37°C in a CO2 incubator and were then aspirated. Cells were overlaid with 2 mL of DMEM-F12 containing 0.75% agarose (Sigma, Milwaukee, WI), 10% FBS, and 1% penicillin–streptomycin (Hyclone Laboratories, Logan, UT). The plates were incubated for 3 days and stained with neutral red. Plaques were counted after incubation at 37°C in CO2 for 3–5 h.
FCV-F9 plaque assays were conducted using confluent CRFK cells in six-well plates as described earlier (D'Souza et al., 2006). Confluent CRFK cells were inoculated with 10-fold serial dilutions of viral samples in DMEM-F12 containing 2% FBS. Viruses were adsorbed for 2 h at 37°C in a CO2 incubator. Cells were then overlaid with 2 mL of DMEM-F12 containing 0.75% agarose, 2% FBS, and 1% penicillin–streptomycin. The plates were incubated for 2 days and stained with neutral red. Plaques were counted after incubation at 37°C in CO2 overnight.
MS2 plaque assays were conducted by adding 0.7 mL of 10-fold serially diluted MS2 and 0.3 mL of 6-h E. coli B-15597 culture to 0.6% molten agar, mixing, and pouring on tryptic soy agar plates (Bae and Schwab, 2008). The plates were incubated at 37°C overnight and then plaques were counted.
Statistical analysis
Results from the treatments and controls were statistically analyzed using analysis of variance with SAS software (version 9.2; SAS Institute, Cary, NC) and Tukey's test on a completely randomized design with two or more replications.
Results
The effect of PJ and PP on cell lines
The cytotoxicity assays of PJ or PP were performed by incubation of PJ or PP at various concentrations on confluent CRFK or RAW 264.7 cells for 2 and 3 days, respectively, and then stained with neutral red solution. Cytopathic effects were also determined by visual inspection under the optical microscope. For PP at 8 and 16 mg/mL, at least a 2-log10 serial dilution in DMEM-F12 containing FBS was required to eliminate any cytotoxic effects on both cell lines. For PJ and PP at 4 mg/mL, a 1-log10 dilution was required to perform plaque assays and to determine their antiviral effects.
Antiviral activity of PJ
The antiviral activity of PJ against FCV-F9, MNV-1, and MS2 at a titer of ∼7 and ∼5 log10 PFU/mL was evaluated. When viruses at a titer of ∼7 log10 PFU/mL were treated with PJ, the viral titers of FCV-F9 and MS2 were found to decrease by 1.20 and 0.63 log10 PFU/mL, respectively, whereas MNV-1 titer was not significantly different from untreated controls (Table 1). When viruses at lower titers (∼5 log10 PFU/mL) were treated with PJ, the viral titers of FCV-F9, MNV-1, and MS2 were decreased by 2.56, 1.32, and 0.32 log10 PFU/mL, respectively (Table 1).
Within each column, means followed by the same superscript letter are not significantly different using the Tukey's test (α = 0.05).
FCV, feline calicivirus; MNV, murine norovirus; PJ, pomegranate juice; PP, pomegranate polyphenols.
To understand if the antiviral effect of PJ was solely due to low pH (pH 3.4), the pH of PJ was adjusted to pH 7.0 before treating the viruses. Our results show that neutralizing PJ did not diminish the antiviral activity of PJ. For low titers, the antiviral effects of PJ at pH 7.0 were similar to PJ at pH 3.4 for FCV-F9, MNV-1, and MS2 (Table 1). For high titers, the reduction in MS2 viral titer by neutralized PJ was less than that by PJ at pH 3.4. Surprisingly, neutralized PJ had increased antiviral activity on high-titer FCV-F9 than PJ at pH 3.4. These results suggest that pH is not the main factor that contributes to the antiviral activity of PJ and that the bioactive compounds in PJ also play an important role in reducing the infectivity of the tested viruses.
Antiviral activity of PP
PP was found to have antiviral effects against the three tested viruses. Reduced viral titers were observed for FCV-F9, MNV-1, and MS2 after treatment with PP at concentrations of 4, 8, and 16 mg/mL, respectively. PP at all three concentrations decreased the FCV-F9 titers to undetectable levels for both high and low titers of FCV-F9. For low-titer experiments, MNV-1 titers were decreased by 1.30, 2.11, and 3.61 log10 PFU/mL and MS2 titers were reduced by 0.41, 0.45, and 0.93 log10 PFU/mL after treating the viruses with 4, 8, and 16 mg/mL PP, respectively. For high-titer experiments, MNV-1 titers were found to be decreased by ∼1.50 log10 PFU/mL with all three concentrations of PP, and MS2 titers were reduced by 0.32, 0.41, and 0.72 after treating the viruses with 4, 8, and 16 mg/mL PP, respectively (Table 1). MS2 appeared to be more resistant to PP treatment than MNV-1 and FCV-F9.
Time-dependence study of PJ and PP on MNV-1 titer reduction
Time dependence of the antiviral activity of PJ and PP on MNV-1 was also studied. Incubation of PJ or 4 mg/mL PP with low titers of MNV-1 for up to 60 min was carried out and the infectivity of MNV-1 was determined and correlated with incubation time, as shown in Fig. 1. The titer of MNV-1 in water did not change over the duration of 60 min, whereas MNV-1 in PJ and 4 mg/mL PP dropped significantly. The antiviral action of the PJ and PP appeared to be rapid during the first 20–30 min and then slowed down. Within 20 min of mixing, a marked reduction, 0.79 log10 PFU/mL for PJ or 1.15 log10 PFU/mL for 4 mg/mL PP in MNV-1 titer, was observed. A total reduction in MNV-1 titer of ∼1.02 log10 PFU/mL for PJ and ∼1.32 log10 PFU/mL for 4 mg/mL PP treatment were achieved over 60 min of incubation.
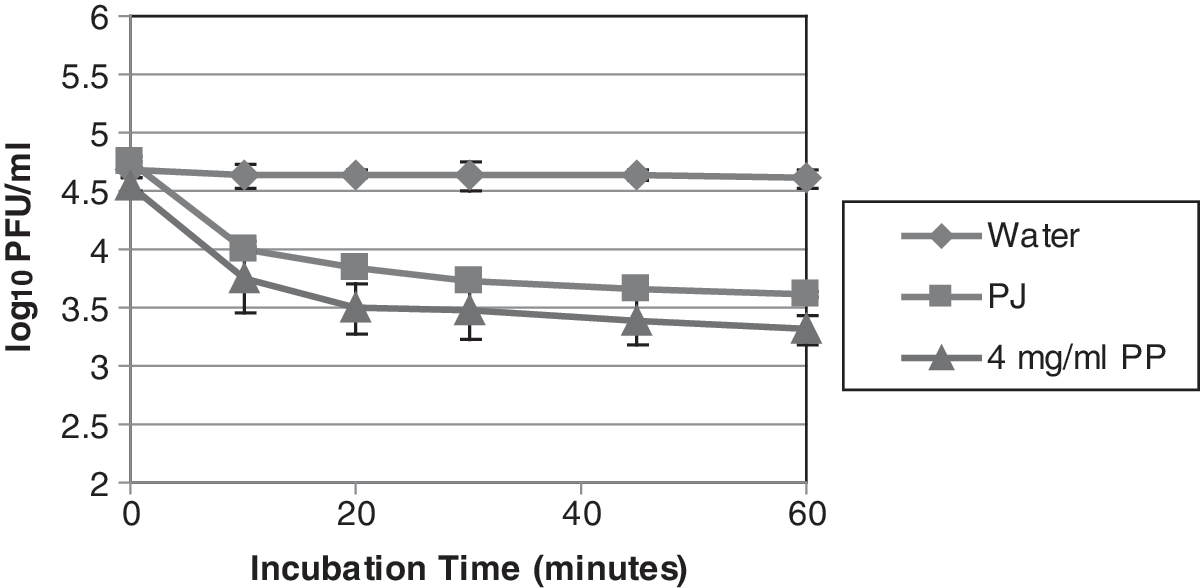
Change in MNV-1 titers after exposure to PJ or 4 mg/mL PP (final concentration after mixing with the viruses) over an incubation time of 1 h. PJ or PP (4 mg/mL final concentration) was added to equal volume of MNV-1 at titers of ∼5 log10 PFU/mL and incubated for up to 60 min at room temperature. The MNV-1 was recovered at 0, 10, 20, 30, 45, and 60 min and assayed for infectivity using standardized plaque assays. MNV, murine norovirus; PJ, pomegranate juice; PP, pomegranate polyphenols.
Understanding the effect of PJ and PP on FCV-F9 and MNV-1 infectivity
To understand if PJ or PP had an effect on viral replication and virus adsorption, PJ or PP solution was added to confluent cells after or before viral infection. Direct addition of 100% PJ was cytotoxic to the cell lines after 45 min of incubation. Therefore, commercial 100% PJ was diluted in Hank's balanced salt solution to 10%–75% juice and used on the cell lines. The juice was found to be cytotoxic if concentrations exceeded 50% for RAW 264.7 cells or 30% for CRFK cells. Thus, these studies were performed with 30% PJ for MNV-1 and 20% PJ for FCV-F9. Regarding cytotoxicity with PP, 1.6 mg/mL PP was found to be cytotoxic for RAW 264.7 cells and 0.8 mg/mL for CRFK cells. Thus, subsequent studies were performed with 0.8 mg/mL PP for MNV-1 and 0.4 mg/mL PP for FCV-F9.
To study the effect of PJ or PP on virus replication, confluent CRFK or RAW 264.7 cells were treated with PJ or PP for 45 min after FCV-F9 or MNV-1 infection. FCV-F9 titers were reduced by 0.75 and 0.40 log10 PFU/mL, respectively, after treatment with 20% PJ and 0.40 mg/mL PP (Fig. 2a). MNV-1 titers were reduced by 0.41 log10 PFU/mL with 30% PJ and no significant change was observed with 0.8 mg/mL PP (Fig. 2a). Thus, 20% PJ and 0.4 mg/mL PP had minor effects on FCV-F9 replication, whereas 30% PJ had some minor effects on MNV-1 replication.
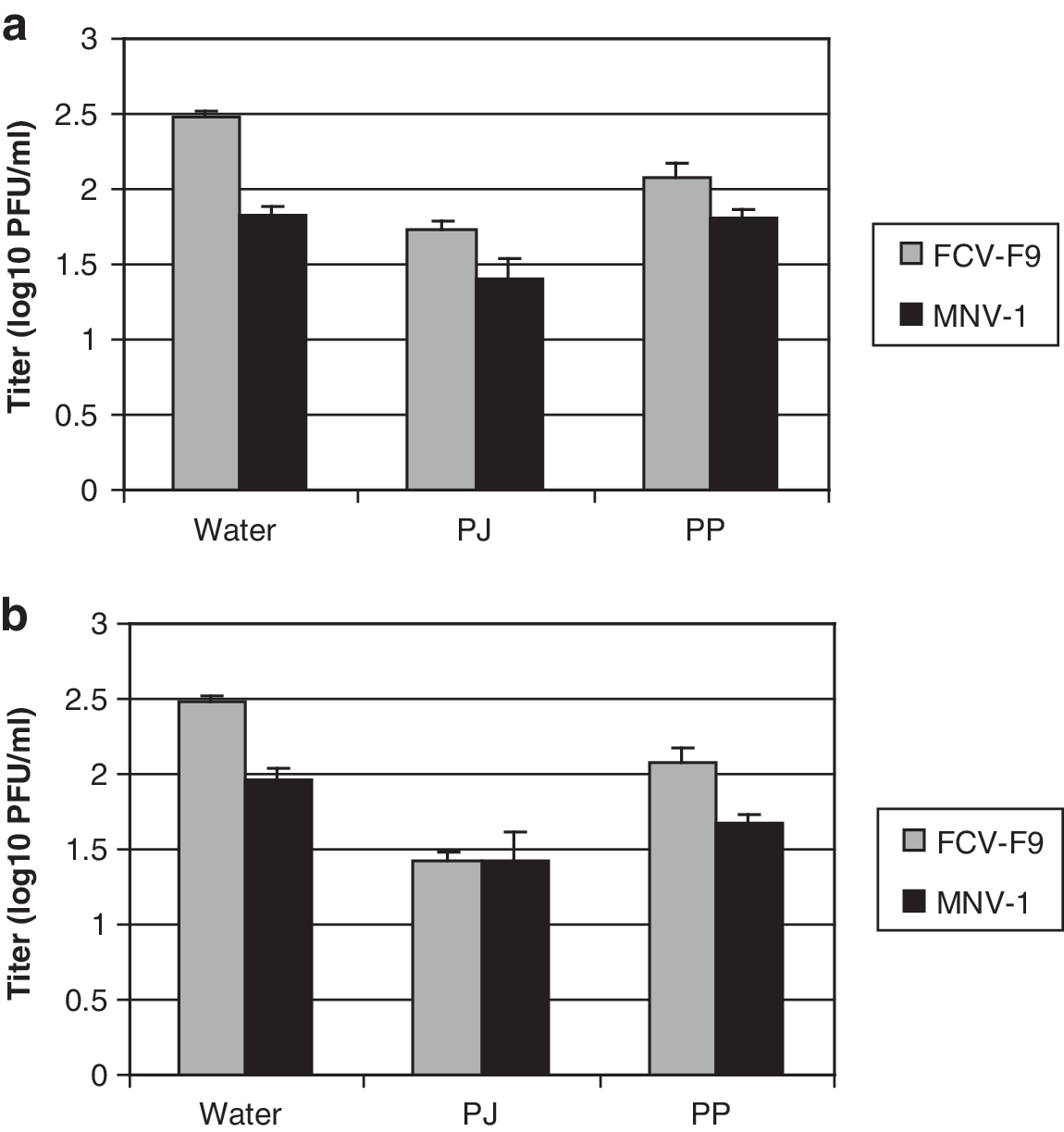
Viral titers of FCV-F9 and MNV-1 recovered after treating confluent CRFK and RAW 264.7 cells, respectively, with water, PJ, and PP. (
When CRFK or RAW 264.7 cell monolayers were pretreated with PJ or PP for 45 min before infection with FCV-F9 or MNV-1, FCV-F9 titers were reduced by 1.05 and 0.40 log10 PFU/mL with 20% PJ and 0.4 mg/mL PP, respectively, and MNV-1 titers were reduced by 0.55 and 0.29 log10 PFU/mL with 30% PJ and 0.8 mg/mL PP, respectively (Fig. 2b). These data show that slightly greater reductions in viral titers were obtained when cells were treated with PJ or PP before infection than postinfection.
Discussion
To the best of our knowledge, even though PJ and pomegranate extracts are reported to have antifungal, antibacterial, and antiviral activities, their effects on human noroviruses or foodborne viruses have not been documented. In the present study, we demonstrated that both PJ and PP have significant effects on the infectivity of foodborne viral surrogates FCV-F9, MNV-1, and MS2. The effectiveness was found to be dependent on both the virus type and the virus titer. With low-titer viruses (5 log10 PFU/mL), the effectiveness of PJ and PP on viral titer reduction followed the order of FCV-F9 > MNV-1 > MS2. When high-titer viruses (7 log10 PFU/mL) were treated, the effectiveness followed the order of FCV-F9 > MS2 > MNV-1. Adjusting the pH from 3.4 to 7.0 did not significantly decrease the antiviral effects of PJ. Thus, the bioactive compounds of PJ, not the pH alone, played a contributory role in the antiviral activity of PJ. Interestingly, PJ at pH 7.0 caused greater titer reduction on FCV-F9 than PJ at pH 3.4. The mechanism of action of PJ is unknown and needs in-depth research. One can only speculate that the neutralized pH could possibly increase the bioavailability of compounds in PJ, such as PP, to FCV-F9 in tested solutions. Alternatively, the neutralized pH could play a role in charge effects that increase the binding of FCV-F9 viral capsid to PJ or PP to cause destruction of the viral capsid or prevent binding of the virus to the host cell receptors.
Pomegranate extracts are known to have antiviral effects against HIV-1, influenza virus, herpes viruses, and poxviruses (Neurath et al., 2005; Jayaprakasha et al., 2006; Kotwal, 2008; Haidari et al., 2009). These effects are attributed to its high content of polyphenols including mainly hydrolysable tannins (ellagitannins), such as oligomers and punicalagin/punicalin, with small amounts of ellagic acid and anthocyanins (delphinidin, cyanidins, pelargonidin) and their glycosides (Aviram et al., 2008). We have not analyzed which component of PJ is responsible for the antiviral activities observed in our study. However, Haidari et al. (2009) tested four flavonoid compounds of pomegranate, namely the ellagic acid, caffeic acid, luteolin, and punicalagin, and found that only punicalagin had inhibitory effects on influenza virus. Punicalagin is a high-molecular-weight, water-soluble polyphenol responsible for >50% of the potent antioxidant activity of PJ (Gil et al., 2000). Besides antiviral activity, punicalagin also has been reported to have antifungal and antibacterial properties (Machado et al., 2002; Taguri et al., 2004; Liu et al., 2009). PJ contains total phenolics of 3.6 mg/mL (Aviram et al., 2008). Thus, mixing PJ with equal volume of viruses results in 1.8 mg/mL polyphenols. Therefore, 4 mg/mL PP (the final concentration after mixing), the lowest concentration of PP used in our study, contains twice the amount of polyphenols as in the experiments using PJ. Therefore, it is not surprising that the antiviral activity of PP at 4 mg/mL is consistently greater than that of PJ for all viruses at both high and low titers. In addition, the differences in composition and bioavailability of polyphenols in PJ to that of PP in its concentrated form could play a role in the increased reductions observed.
As MNV-1 is known to be quite resistant to most treatment conditions, including pH and heat (Cannon et al., 2006), but is susceptible to treatment with PJ and PP, our results suggest that PJ and PP appear to be promising natural alternatives for treating human norovirus infections. Recently, we also studied the effect of cranberry juice (CJ), grape juice (GJ), and orange juice (OJ) on the infectivity of human enteric viral surrogates (Su et al., 2010). For comparison purposes based on these studies, the titer reduction for FCV-F9, MNV-1, and MS2 follows the order of CJ > PJ > GJ > OJ in general, with the exception that GJ has greater effect on high-titer FCV-F9 than CJ.
Several mechanisms on the antiviral activity of polyphenols have been proposed. Haslam (1996) suggested that plant polyphenols exert a direct action on the viral particles, inhibiting the adsorption of the virus to the cell receptors. Cheng et al. (2005) studied the antiviral activity of proanthocyanidin A-1 against herpes simplex virus type 2 and found that proanthocyanidin A-1 did not reduce viral infectivity but inhibited viral attachment and penetration and also affected the late stages of herpes simplex virus type 2 infection. Liu et al. (2007) studied the inhibitory effect of tea polyphenols against HIV-1 and concluded that the inhibitory effect is through multiple mechanisms of action, including inhibiting HIV-1 reverse transcriptase and protease activity, blocking gp120-CD4 interaction by binding to cellular CD4 molecular, and destroying viral particles.
From this study, we cannot allude to which mechanism(s) play a role in the inactivation of the tested nonenveloped foodborne enteric viruses by PJ and PP. However, our results showed that treating the cell layer with PJ or PP before infection caused reduced infectivity of FCV-F9 and MNV-1. This suggests that PJ and PP may possibly play a role in inhibiting the binding of viruses to the cells by blocking the cell surface receptors. When cells were treated with PJ or PP postinfection, reduced infectivity was observed for FCV-F9 by both PJ and PP, but MNV-1 infectivity was reduced only by PP but not PJ. PJ and PP may cause structural damage to the viral capsids of FCV-F9, whereas the capsids of MNV-1 may have higher structural integrity and are more resistant to the PJ treatment. However, based on these current studies we cannot determine the cause of viral titer reduction upon treatment with PJ or PP. Further studies using transmission electron microscopy may help elucidate the mechanism of action of PJ and PP against foodborne viral surrogates.
In conclusion, the present study demonstrated that PJ and PP are capable of causing reduction in titers of foodborne viral surrogates FCV-F9, MNV-1, and MS2 (especially on low titers). However, PJ or PP alone cannot decrease the viral titers to levels that may be significant enough to protect public health, especially based on the results with MNV-1. Combination of PP with high pressure or radiation could help reach the desired levels of inactivation, without altering the sensory attributes of food products. Thus, PJ and PP show promise to be used as part of a hurdle technology for inactivating foodborne viruses. Considering the fact that PJ and pomegranate extracts have been used in folk medicine for centuries without reports of any side effects and allergic reactions, PJ and pomegranate extracts appear to be promising natural remedies for preventing or reducing human norovirus infections. However, further studies including clinical trials and FDA approvals are necessary before any recommendation or claims about PJ or PP can be made for routine human use.
Footnotes
Acknowledgment
Funding for this research was provided by the Tennessee Agricultural Experiment Station.
Disclosure Statement
No competing financial interests exist. The use of trade names in this article does imply neither endorsement by the University of Tennessee nor criticism of similar ones not mentioned.