
Abstract
Bovine vaccinia is an emergent zoonosis caused by the Vaccinia virus (VACV). The disease is characterized by the appearance of exanthematic lesions that occur in humans and dairy cows. Previous studies have revealed the presence of infectious viral particles in milk samples during an outbreak of bovine vaccinia in Brazil, indicating the possibility of disease transmission through raw milk. To assess the viability of the virus in milk after thermal treatment and processing procedures, milk samples were experimentally contaminated with 103 plaque forming units (PFU)/mL (group I) and 105 PFU/mL (group II) VACV Guarani P2 virus, and the third group was not contaminated and served as a control. The samples were submitted to storage temperatures in a cold chamber, freezer for 48 hours, and to low temperature long-time treatment. Moreover, the viral viability was evaluated in cheese produced with contaminated milk using 104 PFU/mL VACV Guarani P2. Notably, the virus remained viable in milk after storage for 48 hours in both the cold chamber and the freezer, with a reduction in viral titer of 14.49% and 25.86%, respectively. Group II showed a viral reduction in titer of 61.88% and 75.98%, respectively. Thermal treatment 65°C for 30 minutes showed a reduction of viral titer of 94.83% and 99.99%, respectively, in group I and group II, but still showed remaining viable virus particles. In addition, it was possible to recover infectious viral particles from both the solid curds and the whey of the cheese produced with experimentally contaminated milk. The cheese shows a reduction in viral titer of 84.87% after storage at 4°C for 24 hours. The presence of viable viral particles in milk after both thermal treatment and cheese production indicates a potential public health risk.
Introduction
Bovine vaccinia (BV) is an emergent zoonosis caused by the Vaccinia virus (VACV), which is a virus of the Poxviridae family and the Orthopoxvirus (OPV) genus (Damaso et al., 2000; Lobato et al., 2005; Trindade et al., 2006). The infection is characterized by the development of lesions in the cows' teats and on the hands of the workers milking them. Infection begins in both cows and humans with the formation of macules followed by edema and local pain that evolves into pustules, ulcers, and crusts until complete wound healing occurs (Trindade et al., 2007). The disease is responsible for significant socioeconomic damage that includes temporary removal of the individual from the work environment. Moreover, the disease causes a significant reduction in milk production in the cows as well as medical expenses and secondary bacterial infections in cows and humans (de Souza Trindade et al., 2003; Leite et al., 2005).
Using polymerase chain reaction (PCR) assays and cell culture inoculation techniques, previous studies by our group (Abrahão et al., 2009a) have revealed the presence of VACV DNA as well as viable viral particles in milk from diseased cows. These results have raised awareness of the possibility that milk and its derivates may serve as a vehicle for the spread of viruses, a situation which can represent a potential and significant public health risk due mainly to the consumption of cheese made from raw milk. Although human infection of VACV through the consumption of milk from diseased cows has not been described in Brazil, Buffalopox, a strain of VACV that afflicts buffalos in India, was identified as the etiological agent of oropharyngeal lesions in humans who consumed milk that had not been previously pasteurized (review by Berkelman, 2003). Despite the possibility that milk serves as a vehicle for VACV and other viruses, resistance to infectious VACV particles in milk subjected to different thermal treatments and to different techniques for the production of cheese and yogurts remains poorly studied (Tomasula and Konstance, 2004; Bona et al., 2005). For the most part, the criteria for milk quality control involve only the determination of bacterial and somatic cell counts and research on antibacterial residues.
Recently, the Serro region of Minas Gerais, Brazil, suffered an outbreak of BV. This region is noteworthy due to its tradition of producing “Minas” artisanal cheese, mainly made from raw milk by cheese-making families. In these facilities and others affected by BV, milk continues to be normally collected and commercialized, being forwarded to individual or communal expansion tanks and/or subsequently to the milk industry where it is processed and commercialized (Madureira, 2009). However, some facilities use a pasteurization method in which the milk remains in special vats under constant mechanical agitation and temperatures varying from 62°C to 65°C according to Brazilian law (Brazil, 1952). Although not adapted to an industrial scale, this process can be used in small dairies for public supply or for the production of milk derivatives according to the Normative Instruction number 5 (Brazil, 2002). Therefore, considering the lack of information regarding the risks involved in the consumption of milk contaminated with VACV, the present study aims at evaluating the viability of the VACV Guarani P2 virus (GP2V) in samples of milk experimentally contaminated and subjected to different thermal treatments. The thermal conditions selected in this work were based on common milk handling practices such as storage at 4°C for until 48 hours and slow pasteurization in small farms and cheese producing industries. Further, the viability of GP2V was also evaluated in milk after the production of cheese. This is the first work that investigates the viability of VACV in milk and its derivatives after thermal treatments.
Materials and Methods
Cells and viruses
Vero cells, a continuous cell line derived from the kidney epithelia of the African green monkey (Cercopithecus aethiops), were multiplied in minimum essential media (MEM) eagle (Gibco) supplemented with 5% fetal bovine serum (FBS) (Cultilab), gentamycin (50 μg/mL), penicillin (200 U/mL), and fungizone (2.5 μg/mL) under incubation with 5% atmospheric CO2 at 37°C. The GP2V sample was isolated from a lesion in a cow's teat from a 2001 outbreak of BV in the municipality of Guarani located in the state of Minas Gerais, Brazil (Trindade et al., 2006).
Multiplication and purification of GP2V
Vero cells were contaminated with a multiplicity of infection of 0.01 plaque forming units (PFU)/cell. After 90% of cytopathic effect was achieved, cells were washed with phosphate-buffered saline (PBS) and scraped off the culture vial. The viral suspension was purified as previously described by Joklik (1962) and titrated in Vero cells with the titer represented in PFU/mL (Campos and Kroon, 1993). These viruses were used to contaminate the milk samples.
GP2V viability in contaminated milk after thermal treatments
Samples from standard ultra high temperature pasteurized whole milk were aseptically divided into 30 mL aliquots and contaminated with GP2V in three groups. Group I was inoculated with 103 PFU/mL, group II with 105 PFU/mL, and the third group was not contaminated and served as a control.
After viral contamination, the samples were subjected to storage conditions at 4°C and −20°C, both of which were performed for a period of 48 hours. The samples that were subjected to a temperature of 65°C for 30 minutes (low temperature long time treatment, LTLT) were maintained in a microcontrolled water bath equipped with a digital temperature controller and were maintained under constant agitation during the experiment. Each experiment was repeated eight times. Before and after each thermal treatment, 1 mL aliquots were withdrawn for the determination of viral titer reduction.
GP2V viability in contaminated milk after cheese production
For the production of cheese, 500 mL of type “C” pasteurized milk were produced using the rapid pasteurization system (high temperature short time), in which the milk was subjected to binomial plates at 72°C–75°C for a period of 15–20 seconds. Eight samples from different producers of milk were used. The samples were refrigerated at 5°C and subsequently warmed to a temperature of 36°C for the addition of 0.05% calcium chloride (50% w/v), 0.07% curd, and 104 PFU/mL of virus. The samples were then left to rest for 30–40 minutes to facilitate the formation of a firm, smooth curd. After coagulation, the resulting solid curd was cut and left unperturbed for 5 minutes. Slow, circular movements were then applied to release the whey. After the whey had been drained, one aliquot was collected to perform the viral titration. The solid curd was collected, deposited in a bowl, maintained at 4°C for ∼24 hours, and subsequently divided into four aliquots. Each aliquot was macerated and diluted in PBS to a ratio of 1:10. The viral titration was performed before and after the collection of the solid curd. A control test was performed by creating cheese from milk in the absence of the virus.
Calculation of the reductions in viral titer
Before and after each thermal treatment and the cheese production, samples were withdrawn from each of the eight replicates. Vero cells were incubated overnight at 37°C and with 5% CO2 in a six-well plate at a concentration of 1.5 × 106 cells per well. The serial dilution of the samples were made in MEM (10−2–10−4); and 300 μL of each serial dilution was adsorbed for 1 hour to the cells and filled out with 2 mL of MEM supplemented with 1% FBS. Cell control wells were maintained without virus. The plates were incubated at 37°C with 5% CO2 and after 72 hours, the medium was discarded and the cells were fixed for 15 minutes in 10% formaldehyde solution in PBS and stained with a 1% crystal violet per 10 minutes. The titer was obtained by counting the number of plaques in the well that showed between 30 and 300 plaques, multiplied by the inverse at the dilution per mL. The titer was represented in PFU/mL (Campos and Kroon, 1993). Subsequently, the mean viral titer and the standard deviation of the viral titer reduction were calculated for each of the eight repetitions. The percentage of viral titer reduction was calculated by counting the number of plaques in the contaminated samples before the treatment (control), subtracting the number of plaques obtained after treatment (mean titer of eight repetitions), divided by the number of plaques in the control, and multiplied by 100 (Malik et al., 2006).
Statistical analyses
In the experimental design, a split-plot design was used for testing a 2 × 3 factorial (two occasions—before and after thermal treatments and three treatments located to subparcels) with eight repetitions (Sampaio, 2007). Each of the repetitions was considered to be a block, and group I (inoculated with 103 PFU/mL), group II (inoculated with 105 PFU/mL), and group III (without viral inoculation) were the parcels. Statistical analyses were performed with the help of Minitab 14 software. The significance analysis was performed through the use of a generalized linear model with the main objective of testing defined as the determination of the significance between the factors, which included the temperature, viral concentration, and the possible effect of interaction between the factors, tested in each assay. Factors and interactions were not considered to be statistically significant unless p-values of <0.05 were observed. Finally, the effect of the eight samples, or blocks, was tested. The analysis of viral viability in cheese was the only effect tested using a paired t-test.
Results
Viability of GP2V in contaminated milk after different thermal treatments
Experimental contaminated milk was used to test the viability of GP2V in milk after thermal treatments. In group I, samples were contaminated with 103 PFU/mL GP2V, group II with 105 PFU/mL, and the third group was not contaminated and served as a control. It was possible to recover infectious viral particles in contaminated milk after storage at −20°C and 4°C and after heat treatment at 65°C in both groups. In group I, the reductions in viral titer at −20°C and 4°C were, respectively, 14.49% (average 0.04 and ±standard deviation [SD] 0.08 log10) and 25.86% (average 0.12 and ±SD 0.12 log10). In group II, the reduction of viral titer for the temperatures −20°C and 4°C was, respectively, 61.88% (average 0.48 and ±SD 0.32 log10) and 75.98% (average 0.61 and ±SD 0.22 log10). Although a smaller reduction in viral titer of samples stored at −20°C in both evaluated groups was observed, results show that the mean reduction of viral titer for storage temperatures was similar (p > 0.05) (Fig. 1).

Reduction of the viral titer in GP2V contaminated milk after thermal treatments. Milk contaminated with GP2V was conserved at temperatures of −20°C and 4°C for 48 hours and at 65°C for 30 minutes, after that viral titers were determined. The error bars represent the standard deviation. Asterisks indicate that viral titer reduction is statistically significant (mean of initial viral titer–mean of final viral titer p < 0.05). GP2V, Guarani P2 virus.
The samples submitted to 65°C temperature for 30 minutes (LTLT) showed significant reduction in viral titers in both evaluated viral concentrations (p < 0.05). The mean reductions in viral titers of samples from group I and group II were, respectively, 94.83% (average 2.65 and ±SD 1.15 log10) and 99.99% (average 4.0 and ±SD 0.48 log10). Moreover, a larger absolute reduction in viral titers in samples of group II compared with group I can be observed (p > 0.05) (Fig. 1). There was no recovery of viral particles in the control group.
GP2V viability in cheese made with contaminated milk: solid curd and whey
After the production of cheese from milk experimentally inoculated, it was possible to recover infectious viral particles in solid curd and whey. The weight of cheese produced was 50 g (5 cm of the height and 6 cm the diameter). Comparing the viral titers of solid curd before and after storage at 4°C for 24 hours, a reduction of 84.87% (average 0.79 ±SD 0.2 log10) of initial viral titer was observed. Results using a logarithmic scale and the differences in titer before and after cheese processing and incubation were significant (p-value = 0.000) for the bilateral test (Fig. 2). The recovered viral titer from whey drained from solid curd was 2.62 log10. There was no recovery of viral particles in the control group.
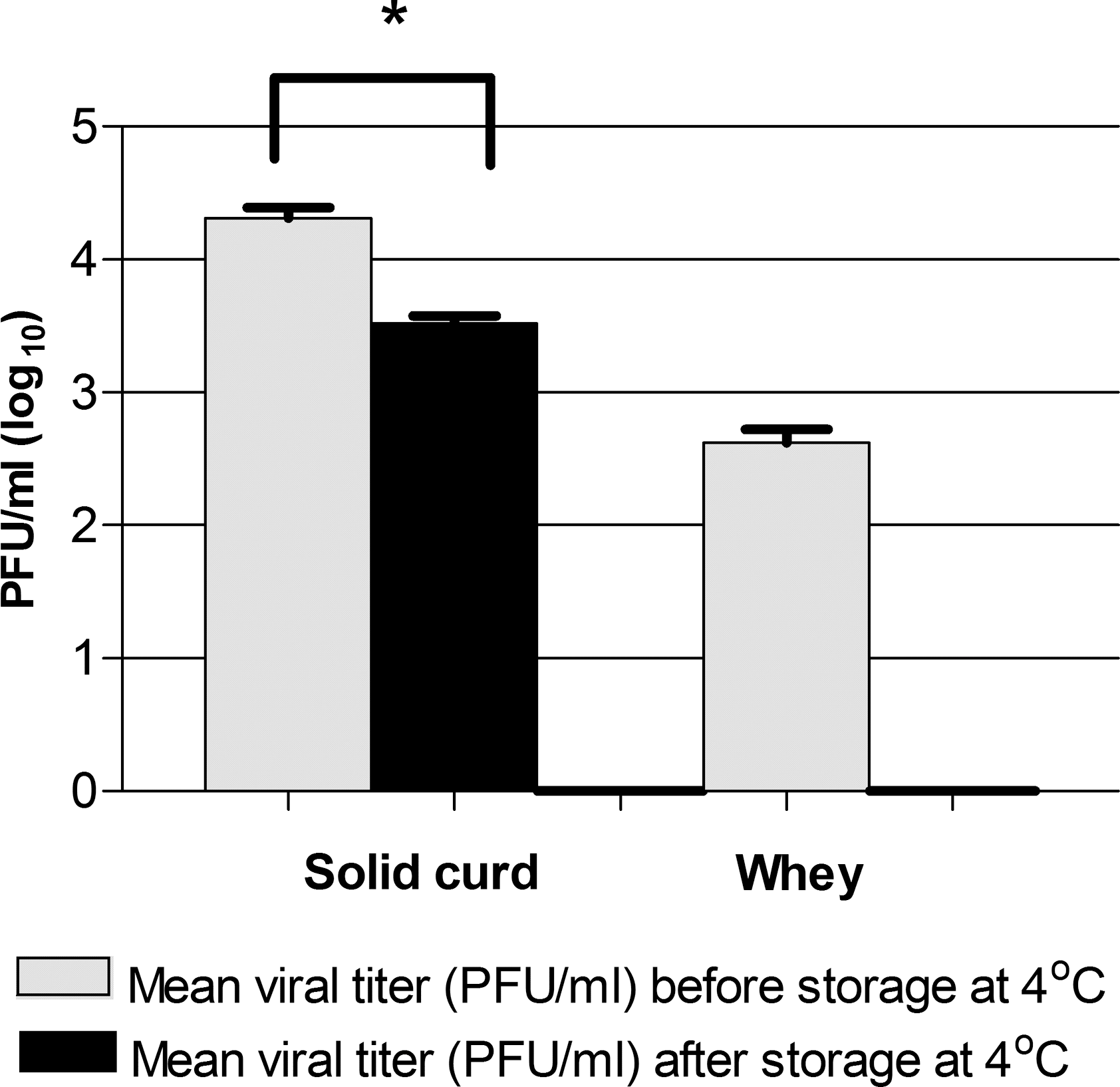
Viral viability in cheese produced with GP2V contaminated milk. The bars represent the mean of virus titers in solid cards before and after storage at 4°C. The error bars show the standard deviation. Significance variations were calculated using the Student's t-test (p < 0.05: statistical significance). Asterisks indicate that viral titer reduction is statistically significant (mean of initial viral titer–mean of final viral titer p < 0.05).
Discussion
Our results showed that it was possible to recover viable virus particles in experimentally inoculated milk that had been subjected to certain thermal treatments. The time interval of 48 hours selected for this experiment had the objective of approximating the time for which milk usually remains in the expansion tanks under refrigeration at 4°C until it is sent to the receiving establishment, as outlined by the agreement set forth in the Normative Instruction Number 51 of MAPA (Brazil, 2002). This instruction indicates that C-type raw milk can be stored at 4°C for up to 24 hours, a timeframe which can be extended to 48 hours when the milk originates from a facility that performs the second milking at the end of the afternoon. The presence of infectious viral particles in the milk after 48 hours of storage at 4°C highlights the possibility of viral spread under these parameters.
Essbauer et al. (2007) evaluated the viability of VACV (Munich 1 sample) in rainwater that was stored at 4°C. The virus remained viable without a significant reduction in the viral titer during the entire experiment, which lasted for 2 weeks. The Variola virus, a virus of the OPV genus, also remained viable for up to 8 weeks in scabs maintained at 4°C or at −20°C (Huq, 1976). Notably, it was possible to detect infectious viral particles in addition to viral DNA. These results are in agreement with the findings presented herein, in which GP2V also remained viable when exposed to temperatures of 4°C or −20°C for up to 48 hours (Fig. 1).
GP2V remained viable in milk samples even after the samples were subjected to a temperature of 65°C for 30 minutes. According to the Normative Instruction Number 51 of MAPA (Brazil, 2002), the LTLT can be adopted by small milk facilities for public supply or for the production of milk-based products. However, the survival of viable viral particles in milk even after LTLT calls attention to the possibility that these products serve as vehicles for viral transmission, thus posing a significant public health risk.
The preservation of GP2V viability in milk even after thermal treatment could be associated with the fat found in ultra high temperature whole milk. In research on the viability of the foot and mouth disease virus in milk, Callis et al. (1975) observed that this virus remained viable in whole milk after heating at 72°C for 3 minutes. However, in skim milk, which has a low fat content, it was not possible to obtain viable viral particles after 15 seconds at the same temperature. According to these authors, the presence of higher concentrations of fat in whole milk diminished the effect of heat on the viral particles.
The results of this study highlight the resistance of GP2V in milk to certain temperatures. Interestingly, previous studies have reported that VACV also displayed resistance to dry heat after 1 hour of exposure to temperatures of 40°C, 75°C, 85°C, and 95°C (Sauerbrei and Wutzler, 2009). This same work demonstrated that a significant reduction in the viral titer (reduction of at least 4 log10 of the original titer) was only observed when samples were exposed to a temperature of 95°C for 2 hours.
After the production of cheese from experimentally contaminated milk, it was also possible to recover infectious particles from both the solid curd and the whey (Fig. 2).
The reason that the viral titer of GP2V recovered in the whey was lower than the titer recovered from the solid curd might be explained by the large volume of whey produced (∼500 mL) before processing and storage. This fact could favor the dispersal of the particles. In the solid cheese curd, the viral particles were concentrated in a small volume, with an average weight of 50 g. The viral viability in solid curd might be mainly attributed to the presence of proteins capable of offering protection to the virus. According to Machado et al. (2004), the protein concentration in Minas artisanal cheese is roughly 17 g/100 g. Some authors highlight the viability of VACV and other OPVs in protein-rich environments. Essbauer et al. (2007) also observed the influence of the presence of proteins in the viability of VACV. The virus remained viable for >166 days in rainwater supplemented with FBS, a protein-rich medium, when stored at 4.5°C. In rainwater without FBS, the virus remained viable for only 56 days.
Additionally, the slightly acidic pH of unripened Minas cheese, which varies from 5.0 to 6.2 according to Silva et al. (2003), might favor the viability of the virus. While evaluating the viability of VACV in a slightly acidic medium (pH 4.5–5.7), Essbauer et al. (2007) indicated that the viral titer remained the same for up to 56 days in samples that were experimentally inoculated and incubated at 4.5°C.
In Minas Gerais (Brazil), the State Law Number 14.185 of 31/01/2002 states that the beginning of artisanal Minas cheese processing must occur within 90 minutes after the initial milking without any previous thermal treatment of the milk (Minas Gerais, 2002). In disease hotspots, the utilization of raw milk for the production of cheese can present risks for both the cheese-maker, who typically manipulates the curd without any type of protection (such as gloves), and the individuals consuming these products.
Raw milk used in the production of cheese and its derivatives represents a potential risk for the dissemination of diseases arising from infectious agents excreted with the milk as well as those acquired from the use of contaminated utensils at the time of milking (Bona et al., 2005; Oliver et al., 2005).
Conclusions
The GP2V remained viable in the milk after storage and LTLT. Further, infectious viral particles were recovered from cheese and whey produced with experimentally contaminated milk. These findings point to potential public health concerns and the need to consider reexamination of current milk handling regulations.
Footnotes
Acknowledgments
We thank Grazielle Gallinari, Fábia Campos, and colleagues from Laboratório de Vírus (ICB-UFMG/Escola de Veterinária) for their excellent technical support. Financial support was provided by the CNPq, CAPES, and FAPEMIG. E.G. Kroon and Z.I.P. Lobato receive fellowships from CNPq.
Disclosure Statement
No competing financial interests exist.