
Abstract
We evaluated the antimicrobial resistance of Salmonella isolated in 2008 from a chicken hatchery, chicken farms, and chicken slaughterhouses in China. A total of 311 Salmonella isolates were collected from the three sources, and two serogroups of Salmonella were detected, of which 133 (42.8%) consisted of Salmonella indiana and 178 (57.2%) of Salmonella enteritidis. The lowest percentage of S. indiana isolates was found in the chicken hatchery (4.2%), followed by the chicken farms (54.9%) and the slaughterhouses (71.4%). More than 80% of the S. indiana isolates were highly resistant to ampicillin (97.7%), amoxicillin/clavulanic acid (87.9%), cephalothin (87.9%), ceftiofur (85.7%), chloramphenicol (84.9%), florfenicol (90.9%), tetracycline (97.7%), doxycycline (98.5%), kanamycin (90.2%), and gentamicin (92.5%). About 60% of the S. indiana isolates were resistant to enrofloxacin (65.4%), norfloxacin (78.9%), and ciprofloxacin (59.4%). Of the S. indiana isolates, 4.5% were susceptible to amikacin and 5.3% to colistin. Of the S. enteritidis isolates, 73% were resistant to ampicillin, 33.1% to amoxicillin/clavulanic acid, 66.3% to tetracycline, and 65.3% to doxycycline, whereas all of these isolates were susceptible to the other drugs used in the study. The S. indiana isolates showed resistance to 16 antimicrobial agents. Strains of Salmonella (n = 108) carrying the resistance genes floR, aac(6’)-Ib-cr, and bla TEM were most prevalent among the 133 isolates of S. indiana, at a frequency of 81.2%. The use of pulsed-field gel electrophoresis to analyze the S. indiana isolates that showed similar antimicrobial resistance patterns and carried resistance genes revealed six genotypes of these organisms. Most of these isolates had the common pulsed-field gel electrophoresis patterns found in the chicken hatchery, chicken farms, and slaughterhouses, suggesting that many multidrug-resistant isolates of S. indiana prevailed in the three sources. Some of these isolates were not derived from a specific clone, but represented a variety of genotypes of S. indiana.
Introduction
The widespread use of antimicrobial agents in food-animal production has contributed to the decreased susceptibility of Salmonella to antibiotics, and this resistance can be transmitted to humans through food products, particularly those of animal origin (Threlfall, 2002). The large volume of manure produced in areas where poultry production is concentrated can contribute to the contamination of ground and surface water (Mallin and Cahoon, 2003). In recent years, Salmonella of animal origin has shown a gradually increasing spectrum of drug resistance on a worldwide scale. The spread of multidrug-resistant Salmonella has not only decreased treatment options and drug efficacy in human salmonellosis, but also enhanced the virulence of the causative organisms, leading in some cases to large-scale outbreaks of foodborne infection (Mølbak, 2005). Foodborne infections caused by multidrug-resistant Salmonella infections have become a serious public health problem in developed countries, and some studies indicate a much greater rate of infection and mortality with such organisms than with drug-sensitive strains of Salmonella (Helms et al., 2002).
Multidrug-resistant Salmonella of animal origin constitute an even more serious problem in China than in developed countries of the world (Cui et al., 2009; Xia et al., 2009). Findings we made before conducting the present study indicated that multidrug-resistant Salmonella now contaminate 67% of domestic animals in China, with some strains resistant to 17 different antimicrobial agents (Chen et al., 2008). Yet knowledge is lacking about the sources of Salmonella contamination in chicken hatcheries, chicken farms, and chicken slaughterhouses in China. We therefore conducted the present study to evaluate the species and types of Salmonella and their prevalence in these chicken-production facilities and to characterize multidrug-resistant Salmonella isolates by serotyping, analysis of their antibiotic resistance patterns, and investigation of their genomes for genes known to confer antibiotic resistance through use of the polymerase chain reaction (PCR) and pulsed-field gel electrophoresis (PFGE).
Materials and Methods
Isolation, identification, and serotyping of Salmonella
In 2008, we collected a total of 992 strains from one chicken hatchery, two chicken farms, and two chicken slaughterhouses in Shandong, China. Fecal samples were taken with disposable sterile swabs from one chicken hatchery, two chicken farms, and two slaughterhouses. One hundred ninety-eight fecal samples from healthy 1-day-old chickens at the hatchery were collected with sterile swabs, 240 fecal samples from healthy 6-week-old chickens at the chicken farm were collected with sterile cloacal swabs, and 554 samples from the cutting room of slaughterhouses were collected in our laboratory from chicken meat.
After incubation in sterile selenite cystine broth (Becton-Dickinson Difco Laboratories, Detroit, MI) at 37°C for 24 h, all of the strains collected were grown on a chromogenic medium selective for Salmonella (CHROM agar; CHROMagar Microbiology, Paris, France). Only one colony per plate was selected for further study, with colonies showing purple coloration on the culture plates presumed to be Salmonella. The colonies were transferred to and grown on nutrient agar (Becton-Dickinson Difco Laboratories) and identified by transfer to tubes with triple sugar iron agar, motility indole–lysine semisolid agar, Voges-Proskauer semisolid medium, urease test broth, and Simmons citrate agar (Becton-Dickinson Difco Laboratories). The isolates examined with these media were simultaneously investigated for presence of the invA gene with the PCR (Wang et al., 2008).
Salmonella isolates were serotyped by the slide agglutination method for their O and H antigens, using the method described by Kauffmann and White for differentiating serovars of the organism (Popoff and Minor, 1992), with antisera produced at the National Institute of Biological Sciences in Beijing.
Susceptibility testing
The minimum inhibitory concentrations (MICs) of Salmonella isolates were determined with the broth microdilution method according to guidelines of the Clinical and Laboratory Standards Institute (CLSI, 2007, 2008). Escherichia coli ATCC 25922 was used as the quality-control strain in determining MIC values. The following antimicrobial agents were tested (concentration range in μg/mL): ampicillin (0.5–128), amoxicillin/clavulanic acid (4/1024–2/512), cefazolin (0.25–256), ceftiofur (0.125–128), tetracycline (0.25–512), doxycycline (0.125–256), chloramphenicol (0.5–512), florfenicol (0.25–512), kanamycin (0.125–256), gentamicin (0.125–256), sulfisoxazole (2–1024), trimethoprim (0.25–512), nalidixic acid (0.25–512), enrofloxacin (0.015–32), norfloxacin (0.03–64), ciprofloxacin (0.015–32), and amikacin (0.125–256). When the distribution of MICs for an antimicrobial agent was bimodal, the breakpoint of susceptibility was set as the midpoint between the two MIC peaks.
PCR amplification and DNA sequencing
On the basis of the MIC data, a total of 133 isolates of Salmonella indiana and 178 isolates of Salmonella enteritidis were analyzed by PCR for genes conferring multidrug resistance. Such genes were detected for isolates resistant to ampicillin (bla TEM and bla PSE), chloramphenicol/florfenicol (catA1, catA2, catA3, cmlA, and floR), tetracycline [tet(A), tet(B), tet(C), tet(D), and tet(G)], streptomycin (aadA, strA, and strB), and fluoroquinolone [qnrA, qnrB, qnrS, aac(6’)-Ib-cr, and qepA]. The primer sequences used in PCR and their predicted sizes are given in Table 4.
PCR was conducted in a final volume of 20 μL containing 1 μL of template DNA, 0.5 μL of each primer at 10 nmol/L, 10 μL of 2 × PCR Mix (TianGen; Beijing, China), and 8 μL of double-distilled water. The variant gene aac(6’)-Ib-cr was further identified by digestion with BstF5I (New England Biolabs, Ipswich, MA). The DNA sequences obtained were compared with those in the GenBank (National Center for Biotechnology Information, Bethesda, MD) using the Basic Logical Alignment Search Tool program to find regions of local sequence similarity.
Pulsed-field gel electrophoresis
The chromosomal DNA of 108 S. indiana isolates carrying the bla TEM, aac(6’)-Ib-cr, and floR genes was digested with the restriction enzyme XbaI and subjected to PFGE according to the Pulse Net Standardized Laboratory Protocol (U.S. Centers for Disease Control and Prevention, Atlanta, GA) using the contour-clamped homogeneous electrical field MAPPER™ System (Bio-Rad Laboratories, Hercules, CA) for PFGE. The agarose gels used in the procedure were run at 14°C for 18 h at a field strength of 6.0 V/cm applied at an angle of 120 degrees, with initial and final switch times of 2.16 and 54.17 s, respectively. Salmonella serovar Braenderup H9812 was used as a standard size marker in the PGFE analysis. Cluster analysis of pulsotypes was done according to the Dice coefficient method (Xia et al., 2009) and using InfoQuest FP Software/Version 4.5 (Bio-Rad).
Results
Identification and susceptibility testing
A total of 311 Salmonella isolates were collected, and two serogroups of Salmonella were detected at the three sources of isolates included in the study (Table 1), consisting of 133 isolates of S. indiana (42.8%) and 178 isolates of S. enteritidis (57.2%). The lowest frequency of S. indiana–positive isolates was at the chicken hatchery (4.2%), followed in increasing order of frequency at the chicken farms (54.9%) and chicken slaughterhouses (71.4%). The lowest frequency of S. enteritidis–positive isolates was at the slaughterhouses (28.6%), followed by the chicken farms (45.1%) and chicken hatchery (95.8%). Among the isolates investigated, 5 of S. indiana and 115 of S. enteritidis came from the chicken hatchery, 28 of S. indiana and 23 of S. enteritidis came from the chicken farms, and 100 of S. indiana and 40 of S. enteritidis came from the slaughterhouses, respectively.
Table 2 shows the distributions of the MICs of 18 antimicrobial agents for the 311 isolates of Salmonella collected in the study. These included several antimicrobial agents of great importance in both human medicine and veterinary practice. The serotypes of Salmonella identified in the study showed significant differences in antimicrobial resistance. Most of the isolates were resistant to sulfamethoxazole (S. indiana, 100%; S. enteritidis, 100%), trimethoprim (S. indiana, 99.2%; S. enteritidis, 62.4%), and nalidixic acid (S. indiana, 100%; S. enteritidis, 99.4%), but the ratios of resistance of S. indiana to trimethoprim were higher than those of S. enteritidis. All of the isolates of S. indiana collected in the study had multidrug resistance. More than 80% of the isolates of S. indiana were highly resistant to ampicillin (97.7%), amoxicillin/clavulanic acid (87.9%), cefalotin (87.9%), ceftiofur (85.7%), chloramphenicol (84.9%), florfenicol (90.9%), tetracycline (97.7%), doxycycline (98.5%), kanamycin (90.2%), and gentamicin (92.5%), and about 60% of the isolates of S. indiana were resistant to enrofloxacin (65.4%), norfloxacin (78.9%), and ciprofloxacin (59.4%). The isolates of S. indiana were susceptible only to amikacin (4.5%) and colistin (5.3%). The isolates of S. enteritidis were resistant to ampicillin (73%), amoxicillin/clavulanic acid (33.1%), tetracycline (66.3%), and doxycycline (63.5%) and were susceptible to cefalotin (6.7%), ceftiofur (6.7%), chloramphenicol (0.9%), florfenicol (0.6%), kanamycin (9.8%), gentamicin (4.5%), amikacin (1.1%), norfloxacin (2.8%), ciprofloxacin (2.8%), and colistin (3.4%). None of the isolates of S. enteritidis showed resistance to enrofloxacin.
AMP, ampicillin; AMC, amoxicillin/clavulanic acid; CEF, cephalothin; XNL, ceftiofur; CHL, chloramphenicol; FFN, florfenicol; TET, tetracycline; DOX, doxycycline; KAN, kanamycin; GEN, gentamicin; AMI, amikacin; SUL, sulfamethoxazole; TMP, trimethoprim; NAL, nalidixic acid; ENR, enrofloxacin; NOR, norfloxacin; CIP, ciprofloxacin; COL, colistin.
The rates of antimicrobial resistance of S. indiana and S. enteritidis did not differ statistically among the three sources of isolates examined in the study (t = 0.05).
Antimicrobial resistance to penicillin, fluoroquinolone, and chloramphenicol
As shown in Table 3, bla TEM, catA1, floR, tetA, strA, and aac(6’)-Ib-cr were the most prevalent resistance genes in the Salmonella isolates from the study sites and were present in S. indiana isolates from all of the sites. Digestion with BstF5I and sequence analysis revealed the variant aac(6’)-Ib-cr gene, which confers resistance to fluoroquinolones in 95 (95%) of the 100 S. indiana isolates from the slaughterhouses, 25 (89.3%) of the 28 S. indiana isolates from the chicken farms, and 3 (60%) of the 5 S. indiana isolates from the chicken hatchery examined in the study. The floR gene, which confers resistance to chloramphenicol and florfenicol, was highly prevalent in S. indiana isolates, being detected in all (100%) of the 28 isolates of this serogroup from chicken farms, 97 (97%) of the 100 isolates from slaughterhouses, and 3 (60%) of the 5 isolates from the chicken hatchery. The bla TEM gene, which confers resistance to ampicillin, was prevalent in both S. indiana (78.4%) and S. enteritidis (62.4%) isolates from all three sources. The qnrA, qnrB, qnrS, and qepA genes were not detected in any isolate.
Among the total of 133 S. indiana isolates, 108 (81.2%) had the bla TEM, floR, and aac(6’)-Ib-cr resistance genes, and this was the most frequently observed combination of resistance genes in the study.
Pulsed-field gel electrophoresis
The S. indiana isolates positive for bla TEM floR and aac(6')-Ib-cr showed six major patterns by PFGE (Fig. 1). Two of these patterns were observed among 3 of the 5 isolates obtained from the chicken hatchery (Fig. 1), and 4 of the patterns were observed among 21 of the 28 isolates obtained from the chicken farms (Fig. 2). Three patterns of combination of the bla TEM floR and aac(6')-Ib-cr genes were observed in 84 of the 100 isolates from the slaughterhouses (Fig. 2). Most of the isolates from the chicken hatchery, chicken farms, and slaughterhouses had the common PFGE patterns observed for the Salmonella isolates from all three of these sources, suggesting that many multidrug-resistant isolates of S. indiana prevailed at the three sources. Some were not derived from a specific clone, but represented a variety of genotypes.
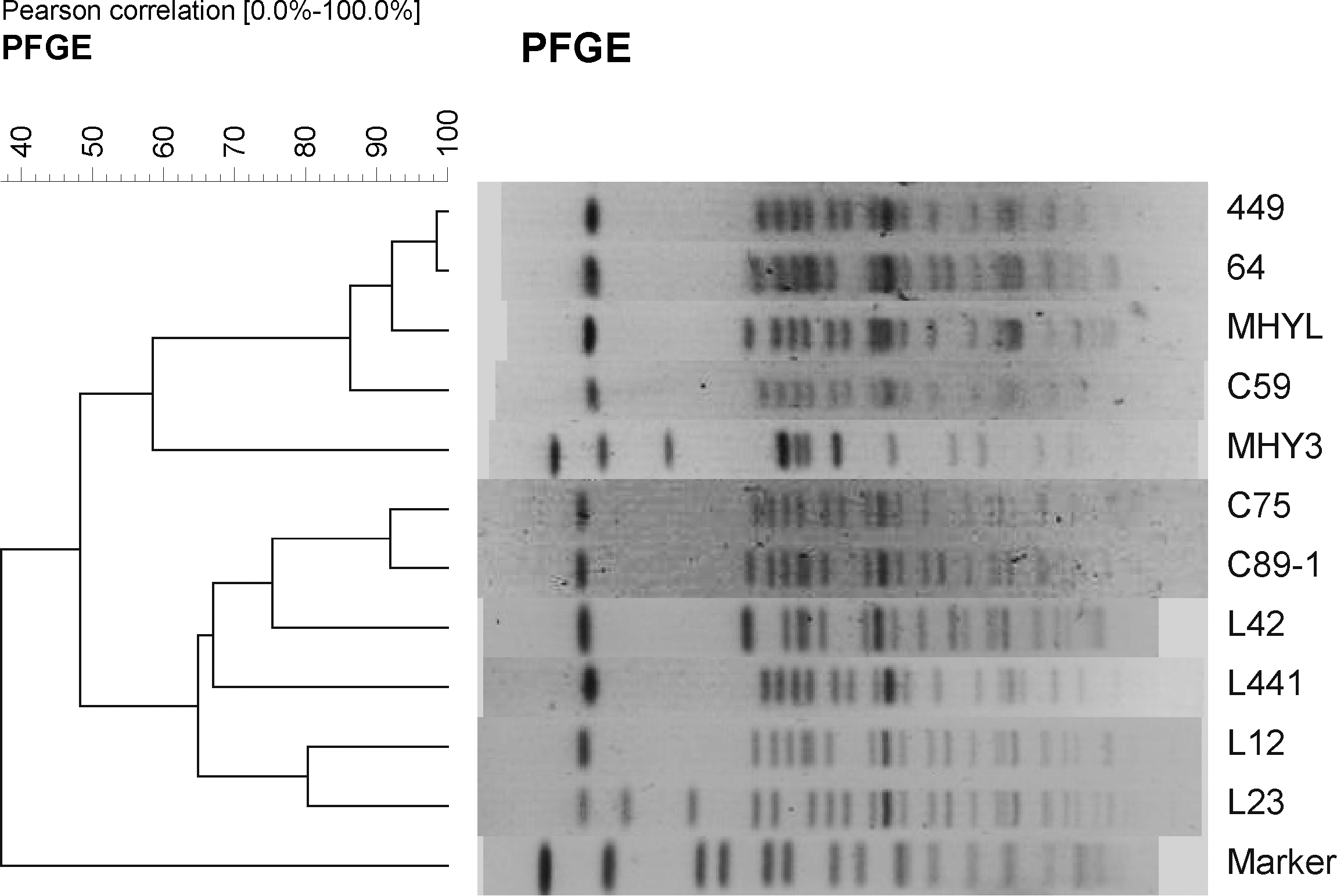
Pulsed-field gel electrophoresis of chromosomal DNA of Salmonella indiana isolates containing antimicrobial resistance genes from poultry production sources in Shandong Province, China. C59, C75, and C89-1 came from a chicken hatchery; L42, L441, L12, L23, and 64 from two chicken farms; 449, MHYL, and MHY3 from two slaughterhouses.
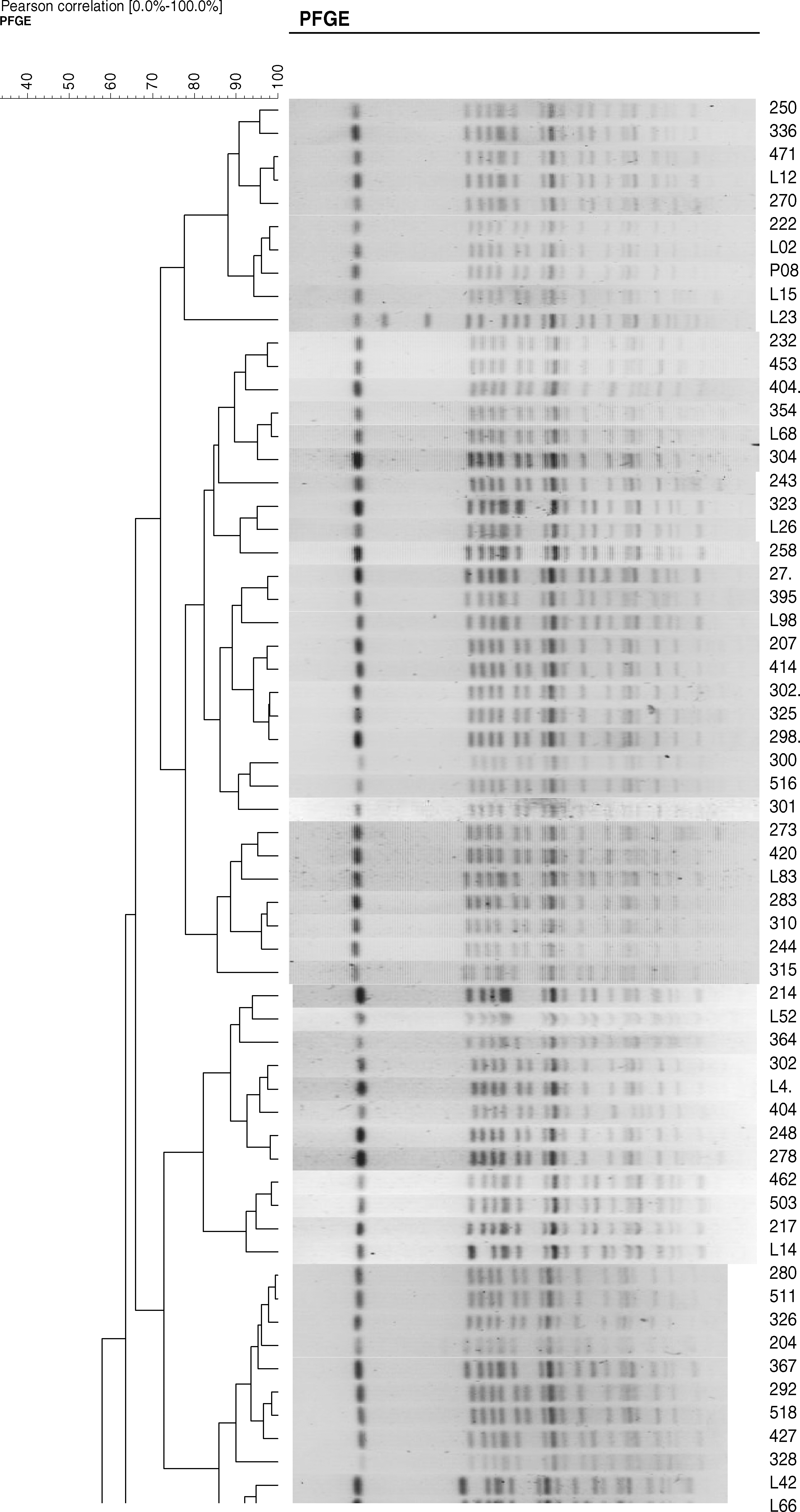
Pulsed-field gel electrophoresis of chromosomal DNA of S. indiana isolates containing antimicrobial resistance genes from two chicken farms and two slaughterhouses in Shandong Province, China. L, chicken farm; Ma, marker.

Discussion
In this study, we collected a total of 311 isolates of Salmonella and detected two serogroups of these organisms (S. indiana and S. enteritidis) at the chicken hatchery, chicken farms, and slaughterhouses that we examined in Shandong Province. No other Salmonella serotypes were found at any of the sites examined in the study, perhaps because S. indiana and S. enteritidis are the prevailing serotypes of Salmonella at chicken farms in Shandong Province. S. enteritidis is widely present in the intestines of poultry and could easily develop antimicrobial resistance under drug pressure.
For S. indiana, however, almost no information is available about antimicrobial drug resistance. Yet the recent emergence of multidrug-resistant Salmonella Typhimurium, Salmonella Paratyphi, and Salmonella Agona suggests that multidrug-resistant phenotypes may also emerge in other serotypes of Salmonella enterica (Boyd et al., 2001; Meunier et al., 2002). The few reports of drug resistance in S. indiana in the domestic Chinese and foreign literature have not found genes indicative of multidrug resistance, yet not only S. indiana, isolated in the present study, is widespread in the kinds of farms and slaughterhouses from which the isolates in the study were obtained, but also these isolates showed characteristics of multidrug resistance. These multidrug-resistant strains had a high resistance not only to streptomycin and tetracycline, but also to chloramphenicol, fluoroquinolones, and cephalosporins.
Contaminated food from animal sources is a major cause of human enteric Salmonella infections (Gordon and Tucker, 1965; Thorns, 2000). During slaughter and processing, Salmonella from the gastrointestinal tract of poultry that carry these organisms can contaminate both other carcasses and the poultry slaughter and processing line (Rostagno et al., 2006). Although efforts at controlling Salmonella have concentrated primarily on controlling contamination within abattoirs, a high proportion of carcasses are still found to be contaminated with Salmonella.
The extensive and unwarranted use of antibiotics to prevent and control bacterial contamination of poultry has resulted in a gradual increase in antimicrobial drug resistance among Salmonella in chicken. Such resistance arises not only in pathogenic but also among symbiotic bacteria in vivo and can be passed through the food chain to the human population. This can in turn lead to microbial cross-resistance and a threat to human health.
As shown in Table 4, bla TEM, catA1, floR, tetA, strA, and aac(6’)-Ib-cr were the most prevalent resistance genes found in our study, being present in S. indiana isolates from all of the poultry sources examined in the study. The aac(6’)-Ib-cr gene, like the other resistance genes named above, was identified by digestion with BstF5I and sequence analysis. The aac(6’)-Ib-cr is a variant of the aac(6’)-Ib gene and is responsible for reduced susceptibility to ciprofloxacin and norfloxacin through the N-acetylation of a piperazinyl amine (Robicsek et al., 2006; Jiang et al., 2008). The gene was present in 95 (95%) of the 100 isolates from the slaughterhouses, 25 (89.3%) of the 28 isolates from the chicken farms, and 3 (60%) of the 5 isolates from the chicken hatchery. Ciprofloxacin is often used to treat salmonellosis. However, the incidence of resistance or reduced susceptibility to fluoroquinolones among Salmonella species has increased worldwide over the past decade, and there have been reports of failed treatment of salmonellosis with ciprofloxacin (Aarestrup et al., 2003). In the United States, the aac(6’)-Ib-cr gene has been found to be widely distributed geographically and stable over time (Park et al., 2006). Our findings suggested that this gene may have been already prevalent among bacteria before the emergence of the qnr genes for fluoroquinolone resistance. The floR gene, also conferring resistance to fluoroquinolones, was, like the aac(6’)-Ib-cr gene, highly prevalent in the S. indiana isolates in our study, being detected in all 28 (100%) of the isolates from the two chicken farms, 97 (97%) of the 100 isolates from the two chicken slaughterhouses, and 3 (60%) of the 5 isolates from the chicken hatchery. The bla TEM gene, conferring resistance to ampicillin, was prevalent in both S. indiana (78.4%) and S. enteritidis (62.4%) isolates, collectively, from the three study sources.
The aac(6’)-Ib-cr variant enzyme was the first reported enzyme to confer resistance to two structurally different families of antibiotics (Courvalin, 2008). The expression of aac(6’)-Ib-cr may facilitate the survival of DNA gyrase and topoisomerase IV mutants (Robicsek et al., 2006). Surveys of antibiotic-resistant Enterobacteriaceae found that the aac(6’)-lb-cr variant enzyme was frequently associated with the expression of an expanded spectrum of β-lactamase enzymes in community-acquired enterobacteria (Karisik et al., 2006; Coque et al., 2008). Chloramphenicol and florfenicol are both members of the molecular family of N-substituted acetamides. In August 1994, chloramphenicol was prohibited in the European Union for use in food animals. Florfenicol, a fluorinated derivative of chloramphenicol, has been introduced to treat respiratory infectious diseases in cattle and pigs and is expected in the near future to see wider use in animal production (Martine et al., 2005).
Multidrug-resistant Salmonella enterica serovar Typhimurium, mainly of definitive phage type 104 (DT104), has emerged during the past decade as a world health problem (Cloeckaert et al., 2001). This bacterium, which causes diseases in humans and animals, is resistant to ampicillin, chloramphenicol, florfenicol, spectinomycin, streptomycin, sulfonamides, and tetracyclines. A chromosomal locus of about 12.5 kb, carrying genes for resistance to all of these antimicrobial agents as well as the chloramphenicol–florfenicol cross-resistance gene floR and included in a 43-kb genomic island called Salmonella genomic island 1, was identified in 1999 (Arcangioli et al., 1999; Briggs and Fratamico, 1999; Boyd et al., 2001). Interestingly, floR was also recently found in other serovars of S. enterica, such as Agona, Albany, Newport, and Paratyphi B, as well as in other Gram-negative bacteria such as E. coli, Klebsiella pneumoniae, and Vibrio cholerae (Schwarz et al., 2004).
Although the six different patterns of bla TEM floR and aac(6’)-lb-cr positivity found by PFGE among the S. indiana isolates in our study were detected at poultry sources, they carried similar resistance genes. Drug-susceptibility tests show that these strains are sensitive only to amikacin and colistin but have high resistance to other drugs including ciprofloxacin and cephalosporins produced for human therapeutic use. The existence and wide distribution of these strains put human health at risk. The application of antibiotics to control and prevent salmonellosis has made drug-resistant salmonellosis an increasingly serious problem and increases the urgency of strengthening the monitoring of resistance to antimicrobial agents.
Southern hybridization confirmed that floR and aac(6')-Ib-cr alleles were located on the chromosome. The alleles of floR and aac(6')-Ib-cr in isolate MHY1 from the slaughterhouses examined in our study were found in the same 23-kb fragment of the gene digests of this isolate; the alleles of floR and aac(6')-Ib-cr in isolate L23 from the chicken farms were found in the same 6.5-kb fragment of this isolate L23. This suggests that the floR and aac(6')-Ib-cr genes may be associated with a common identical or similar set of genes from different sources. The fact that two bacterial serotypes with different genetic backgrounds may carry the same gene fragment for multidrug resistance implies a horizontal transfer of multidrug resistance. We are further investigating the nucleic acid sequences and genetic backgrounds of the isolates in which the alleles of the floR and aac(6’)-lb-cr genes were found.
Footnotes
Acknowledgments
The authors are grateful to Dr. Yu-Qing Liu of the Institute of Animal Science and Veterinary Medicine of the Shandong Academy of Agricultural Sciences, who kindly provided assistance in isolating the Salmonella investigated in the study. The authors also thank Dr. Xiao-Lin Hou of the Beijing University of Agriculture for suggesting revisions to the manuscript. This study was supported by grants from the Program for Chang Jiang Scholars and the Innovative Research Team at the University of China (no. IRT0866), the Exclusive Research Fund for Public Welfare from Ministry of Agriculture of the People's of China (no. 200903055), the Food Safety Key Technology Project of the China National Science and Technology Pillar Program in the 11th 5-year plan period (no. 2006 BAK02A03), and the National Natural Science Foundation of China (no. 30972216).
Disclosure Statement
No competing financial interests exist.