
Abstract
Salmonella is a foodborne pathogenic bacterium that causes human illnesses and morbidity and mortality in swine. Bacteriophages are viruses that prey on bacteria and are naturally found in many microbial environments, including the gut of food animals, and have been suggested as a potential intervention strategy to reduce Salmonella levels in the live animal. The present study was designed to determine if anti-Salmonella phages isolated from the feces of commercial finishing swine could reduce gastrointestinal populations of the foodborne pathogen Salmonella Typhimurium in artificially inoculated swine. Weaned pigs (n = 48) were randomly assigned to two treatment groups (control or phage-treated). Each pig was inoculated with Salmonella Typhimurium (2 × 1010 colony forming units/pig) via oral gavage at 0 h and fecal samples were collected every 24 h. Swine were inoculated with a phage cocktail via oral gavage (3 × 109 plaque forming units) at 24 and 48 h. Pigs were humanely killed at 96 h, and cecal and rectal intestinal contents were collected for quantitative and qualitative analysis. Fecal Salmonella populations in phage-treated pigs were lower (p < 0.09) than controls after 48 h. Phage treatment reduced intestinal populations of inoculated Salmonella Typhimurium in pigs compared to controls at necropsy. Cecal populations were reduced (p = 0.07) by phage treatment >1.4 log10 colony forming units/g digesta, and rectal populations were numerically reduced. The number of pigs that contained inoculated Salmonella Typhimurium was reduced by phage treatment, but a significant (p < 0.05) reduction was only observed in the rectum. We conclude that phages can be a viable tool to reduce Salmonella in swine. Further research needs to be performed to determine the most efficacious dosing regimens and the most effective combinations of phages targeting the diverse Salmonella population found in swine before they can enter the food supply.
Introduction
Bacteriophages are one pathogen-reduction strategy that has received increased attention over the last 10 years because these naturally occurring viruses infect bacteria and reproduce within them, killing the host bacterium through cellular lysis accompanied by the release of daughter phages (Barrow and Soothill, 1997; Kutter and Sulakvelidze, 2005; Johnson et al., 2008). Phages are considered safe having been used as antimicrobials in human medicine in Eastern Europe instead of antibiotics for over 80 years and described as the “infectious cure for infectious disease” (Barrow, 2001). The high degree of specificity phages exhibit for their bacterial hosts offers the potential to serve as a targeted treatment or “designer antimicrobial” (Greer, 2005) in which specific pathogens (such as Escherichia coli O157:H7, Salmonella, and Campylobacter) are removed from the gastrointestinal microflora of food animals before slaughter (Loc Carrillo et al., 2005; Jamalludeen et al., 2009; Wall et al., 2010). In a previous study our research group isolated phages from commercial swine feces and found these phages killed Salmonella Typhimurium (Callaway et al., 2010). Therefore, the present study was designed to determine if these phages could reduce artificially inoculated gastrointestinal populations of the foodborne pathogen Salmonella Typhimurium in a growing-swine model.
Materials and Methods
Bacterial cultures
Salmonella enterica serotype Typhimurium (ATCC BAA-186) from the Food and Feed Safety Research Unit culture collection was repeatedly grown (4 passages) by 10% (vol/vol) transfer in anoxic (85% N2, 10% CO2, 5% H2 atmosphere) Tryptic soy broth medium at 37°C to adapt the culture for growth in the anaerobic intestinal tract. This strain was made resistant to novobiocin and nalidixic acid (20 and 25 μg/mL, respectively) by repeated transfer and selection in the presence of sub-lethal concentrations of each antibiotic. This resistant phenotype was stable through multiple unselected transfers in batch culture and through repeated culture vessel turnovers in continuous culture (data not shown). Overnight cultures (1 L) contained populations of Salmonella Typhimurium that were determined to be 4 × 109 colony forming units (CFU)/mL by serial dilution and plating as described below.
Bacteriophage
Bacteriophages that lysed Salmonella Typhimurium were collected and isolated previously from growing swine feces (Callaway et al., 2010). These phages were tested in vitro during our previous study (Callaway et al., 2010) and only displayed lytic characteristics, with no apparent temperate traits. Phage stocks were prepared by adding phages (multiplicity of infection [MOI] = 0.1–0.001) to a Salmonella Typhimurium culture that was in early exponential growth phase (OD600 < 0.3). After growth was completed (overnight) chloroform was added to cultures followed by vigorous shaking to lyse bacterial cells releasing progeny phage and cultures were centrifuged at low speed (5000 g, 10 min) to remove cellular debris. Phage supernatants were serially diluted and spot tested against lawns of Salmonella Typhimurium to determine phage titer. Phages (n = 2 isolates at individual concentrations of ∼108 plaque forming units [PFU]/mL) were grown individually and pooled for use as a cocktail (total phage cocktail titer of ∼108 PFU/mL).
Swine, rations, and experimental design
All procedures in this study were approved by the Institutional Animal Care and Use Committee (IACUC protocol 08-002). Yorkshire/Duroc crossbred swine (n = 48; average 10 kg BW) were purchased and transported to the FFSRU laboratory. This breed of pigs was chosen because it is a commonly used crossbreed in commercial pork-producing facilities. Pigs were fed a commercial ration composed of (Dry Matter basis) ground corn 63.7%, soybean meal 25.2%, spray-dried porcine plasma 2%, select fish meal 4%, soybean oil 3%, dicalcium phosphate 1.4%, vitamin/trace mineral mix 0.4%, lysine-HCl 0.2%, and DL-methionine 0.1%. The diet was formulated according to National Research Council (NRC) recommendations and pigs were allowed ad libitum access to water. Pigs were housed in groups of n = 12 in environmentally controlled facilities and feces from each pig was sampled on arrival and each subsequent day (n = 5 days) during the dietary/facility adaptation period to verify that no organisms capable of growth on novobiocin (20 μg/mL) and nalidixic acid (25 μg/mL) supplemented Brilliant Green agar (BGANN) were present in the pigs. During this period no colonies grew on any of the BGANN plates. During this period, fecal samples were also analyzed by enrichment for the presence of bacteriophage that could lyse Salmonella Typhimurium (Callaway et al., 2010), the bacterial strain used in the present inoculation study.
Forty eight (n = 48) swine were randomly assigned to one of two treatment groups (n = 24 control and n = 24 phage-treated). Each pig was inoculated with Salmonella Typhimurium (2 × 1010 CFU/pig) via oral gavage (10 mL total volume per pig) at 0 h (Fig. 1). This high dose of Salmonella Typhimurium was chosen to ensure passage through the acid stomach and to ensure relatively high concentrations in the gastrointestinal tract for the phage to have an opportunity to replicate in. Swine did not demonstrate disease symptoms or fever during this short-term infection study. Fecal samples were collected via rectal grab at 24 h intervals after inoculation and populations of inoculated Salmonella Typhimurium were enumerated via serial dilution and plating. Swine were inoculated with the phage cocktail via oral gavage (3 × 109 PFU; phage concentrations determined as described below) or a phosphate-buffered saline sham gavage at 24 and 48 h. The estimated final phage concentration in the intestinal tract at the conclusion of the study was ∼104 PFU/mL intestinal contents.
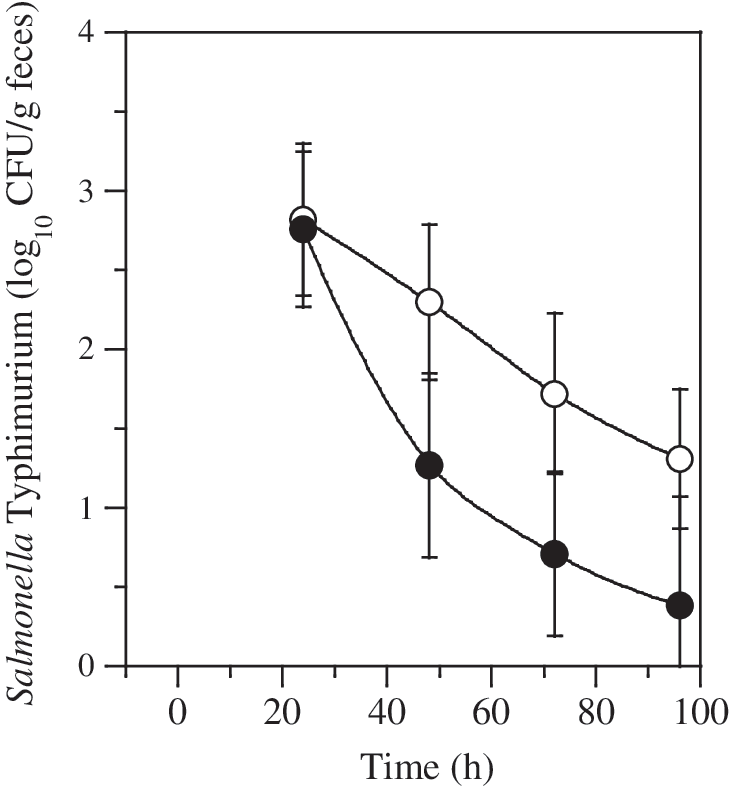
Fecal populations of experimentally inoculated Salmonella Typhimurium (CFU log10/g feces) over time. Open symbols represent controls; closed symbols indicate swine that were treated with bacteriophage at 24 and 48 h. Pigs were inoculated with Salmonella at time 0. Error bars indicate standard deviations. CFU, colony forming units.
Gastrointestinal sample collection
Pigs in both groups were humanely euthanized and exsanguinated at 96 h (Fig. 1). Digesta and epithelial tissues from the distal cecum and terminal rectum were aseptically collected upon necropsy. Ileocecal lymph nodes were collected and qualitatively enriched after maceration (see below). Digesta samples were diluted as described below for quantitative enumeration of intestinal Salmonella Typhimurium populations. Sample aliquots and epithelial tissues were added to tetrathionate broth for primary and secondary qualitative enrichment for Salmonella Typhimurium and re-isolation of treatment phages as described below.
Bacterial enumeration
Cecal and rectal contents and feces were serially diluted (10-fold increments) in phosphate-buffered saline (pH 6.8). Dilutions were plated on BGANN and incubated overnight at 37°C. Colonies that grew on BGANN plates and demonstrated typical Salmonella morphology after 24 h incubation were directly counted (quantitative enumeration). To qualitatively confirm the presence of inoculated Salmonella Typhimurium in samples that were negative for quantitative analysis, intestinal contents and epithelial tissue samples as well as feces were incubated overnight in tetrathionate broth at 39°C and a 200 μL aliquot was transferred to Rappaport-Vassiliadis R10 Broth, which was incubated at 42°C for 24 h. After this secondary enrichment, samples were streaked on BGANN plates. Plates that exhibited colonies after 24 h incubation were classified as positive for experimentally introduced Salmonella Typhimurium. Unless otherwise noted, all media and agar were from Difco Laboratories (Sparks, MD). Reagents and antibiotics were obtained from Sigma Chemical Co. (St. Louis, MO).
Bacteriophage enumeration
Phage populations in pure isolates and in swine intestinal contents were estimated by treating an (2 mL) aliquot of each phage isolate or intestinal contents dilution tube (above) with chloroform to lyse bacterial cells. The supernatant was then spotted (5 μL) on a bacterial lawn of Salmonella Typhimurium. The presence of phage in each diluted sample was determined by the presence or absence of plaques (clearing zones) in the lawn.
Statistics
Salmonella Typhimurium CFU/g were log10 transformed. Treatment groups were compared at each time point by the Mixed procedure of SAS (SAS Institute Inc., Cary, NC). The experimental unit was the individual pig. Time × treatment interactions were discounted due to the natural decay of Salmonella Typhimurium populations in this artificially inoculated model; therefore, only pointwise comparisons were performed. Significance was determined at p < 0.05. Swine positive for Salmonella were analyzed using Pearson Exact Chi Square analysis of SAS.
Results
Swine (n = 48) in both the control and phage-treatment experimental groups shed 2.8 × 103 ± 4.9 × 102 CFU Salmonella Typhimurium/g feces 24 h after inoculation (time of initial bacteriophage treatment; Fig. 1), and there was no difference in fecal shedding between groups. Swine were orally dosed with a placebo treatment or ∼109 PFU of phage/pig at 24 and 48 h, after which populations of Salmonella Typhimurium/g feces declined in both control and phage-treated groups as is typical in experimental inoculation studies (Fig. 1). Fecal populations of Salmonella in phage-treated pigs were lower at all time points after the initial phage application (24 h) but the difference was not significant (p > 0.05).
Phage treatment reduced intestinal populations of inoculated Salmonella Typhimurium in pigs compared to controls when animals were sacrificed at 96 h (Fig. 2). Cecal populations were reduced (p = 0.07) >1.4 log10 CFU/g digesta, and rectal populations were reduced 1 log10 CFU/g feces (p = 0.09). The number of pigs that were positive for inoculated Salmonella Typhimurium upon enrichment was also reduced by phage treatment (23 vs. 13 pigs; Fig. 3). The number of pigs with Salmonella-positive ileocecal lymph nodes was the least reduced by phage treatment (20 vs. 16 pigs), but this result was not unexpected because of the invasive nature of Salmonella but not the phage. Swine that were positive for inoculated Salmonella Typhimurium were reduced by this particular regime of phage treatment, but this decrease was only statistically significant (p < 0.05) for rectal samples. All phage-treated pigs tested positive for introduced phage in all intestinal compartments; in contrast, none of the control pigs' intestinal compartments contained phages that lysed Salmonella Typhimurium (data not shown).
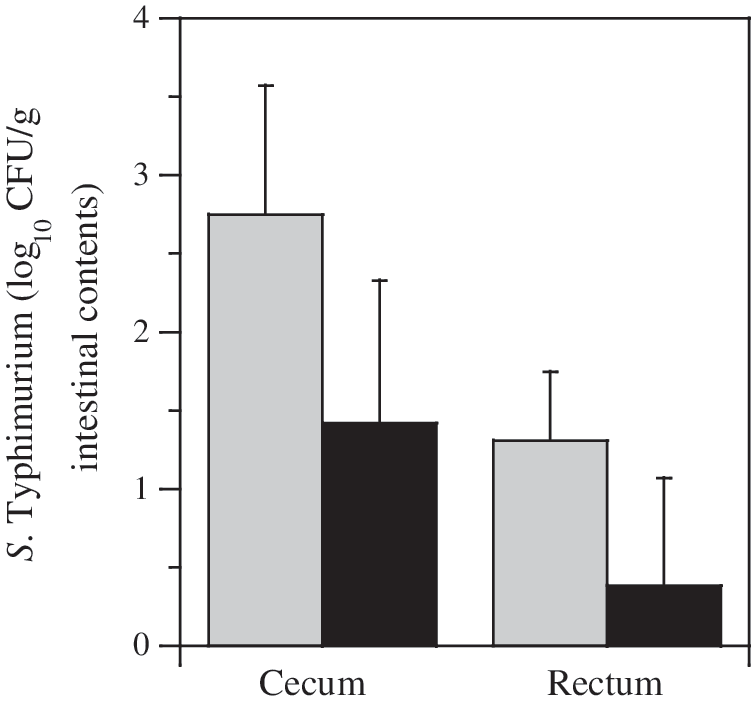
Intestinal populations of experimentally inoculated Salmonella Typhimurium (CFU log10/g digesta) in samples collected from the cecum and rectum. Samples were collected 96 h after Salmonella inoculation and 48 h after final phage inoculation. Gray bars indicate control swine; black bars depict pigs treated with phage. Error bars indicate standard deviations.
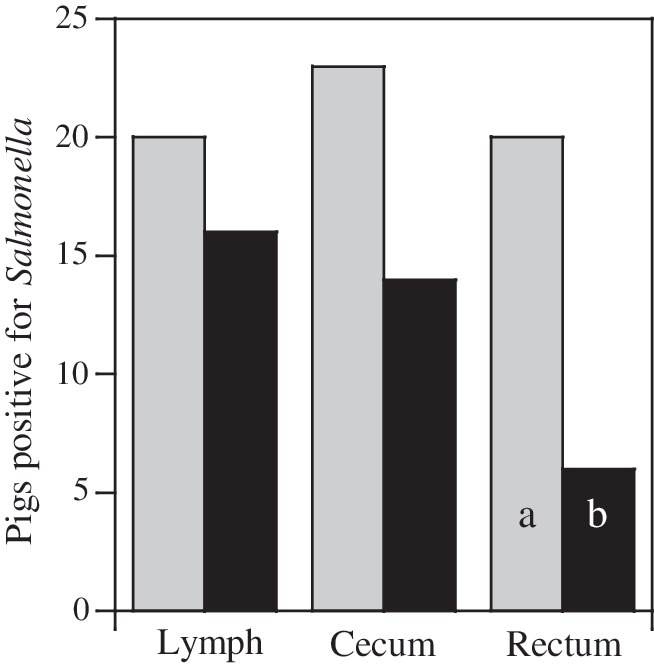
Number of swine positive for experimentally inoculated Salmonella Typhimurium in samples collected from the ileocecal lymph nodes, cecum, and rectum. Lymph node samples represent tissue samples only; cecal and rectal samples indicate Salmonella-positive samples from tissues or contents. Samples were collected 96 h after Salmonella inoculation and 48 h after final phage inoculation. Gray bars indicate control swine; black bars depict pigs treated with phage. Bars with different letters indicate difference of p < 0.05.
Discussion
Salmonella serotypes are one of the most common agents of foodborne human infections around the world and represent a significant drain on the Gross National Product of the United States (Scharff, 2010). In recent years, there has been an increasing focus on reducing pathogenic bacteria in all food animal species before slaughter (Sargeant et al., 2007; Oliver et al., 2008). Although the introduction of new strategies to reduce pathogens in processing plants have been largely successful (Koohmaraie et al., 2005), too many foodborne illnesses still occur. Reducing the pathogen burden before it enters the processing plant has been suggested as a method to enhance the effectiveness of current and future in-plant intervention strategies, and thereby most efficiently reduce human illnesses (Hynes and Wachsmuth, 2000). Because Salmonella is spread easily by direct fecal contact, direct contact and environmental exposure on farms, during transit, and in lairage (Davies et al., 1997; Fedorka-Cray et al., 1997; Rostagno et al., 2003; Rodriguez et al., 2006), any attempt to reduce Salmonella entry into the food chain must be economically feasible enough to be applied to a large segment of the swine population at several control points during the production continuum.
Bacteriophages are viruses that exist in a predator–prey relationship with bacteria and naturally occurring in the gut of swine (McLaughlin and King, 2008; Callaway et al., 2010). These viruses specifically infect bacteria, often only affecting a few target bacterial species, resulting in phages being dubbed a “designer antimicrobial” (Greer, 2005). As a result, researchers have again turned to phages as a method to reduce pathogenic bacteria in many environments, including foodborne pathogenic bacteria in the gastrointestinal tracts of food animals (Greer, 2005; Johnson et al., 2008). Phages have been used in in vivo research studies to reduce the incidence and populations of E. coli that cause disease in weaned pigs, sheep, and poultry (Smith and Huggins, 1983, 1987; Huff et al., 2002). When administered to calves, bacteriophage treatment reduced enterotoxigenic E. coli (ETEC)–induced diarrhea and splenic ETEC colonization (Smith and Huggins, 1987). Air saculitis, a production disease caused by E. coli infection in poultry broilers, was reduced by phage treatment (Huff et al., 2002). More recently, researchers have found that bacteriophage treatment in weaned pigs reduced ETEC populations in the gut and reduced resultant postweaning diarrhea (Jamalludeen et al., 2009).
Research has also been performed examining the use of phages to reduce foodborne pathogenic bacteria in live animals. Phages have been used against E. coli O157:H7 with mixed results (Kudva et al., 1999; Bach et al., 2003). More recently, phages isolated from the feces of feedlot cattle that were active against E. coli O157:H7 reduced E. coli O157:H7 populations in experimentally infected sheep (Callaway et al., 2008). Researchers found that phages that lysed Salmonella Typhimurium isolated from municipal wastewater sources reduced experimentally inoculated Salmonella populations in finished swine (Wall et al., 2010). These researchers found that their phage cocktail reduced phage populations in the ileum and cecum in a production-type environment (Wall et al., 2010).
In implementing phages as a preharvest intervention strategy to kill Salmonella resident in the gut of swine the phage should originate from the gastrointestinal tract of swine, because the fitness of most organisms is typically greatest in their natural environment. In the present study, the two individual phages in the cocktail were isolated from commercial swine feces and selected for their efficacy against this inoculated strain of Salmonella Typhimurium (Callaway et al., 2010). However, Wall et al. (2010) demonstrated that phages isolated from municipal wastewater streams could effectively reduce Salmonella populations in swine. In spite of differing sources of phage isolates, phage cocktails in both studies reduced inoculated Salmonella Typhimurium populations in the intestinal tract of experimentally infected swine. In the present study, the maximum efficacy of Salmonella reduction occurred in the cecum although rectal populations were also reduced. In this study and that of Wall et al. (2010), the number of pigs that were positive for Salmonella was reduced by phage treatment, but the pathogen was not eliminated from phage-treated swine.
Because bacteriophages recognize specific receptors on bacteria, the bacteria can mutate, giving rise to phage-resistant mutants; therefore, in our study we used a cocktail of two phages to prevent the emergence of phage-resistant inoculated Salmonella. In our studies, no phage-resistant Salmonella were detected. The use of phages with narrow spectra of activity must be balanced by the use of more diverse cocktails of phages, or the phages selected for use in swine must have a broad range of serotypes that they are active against to both prevent phage resistance and to enhance the effectiveness of phage as a food safety intervention. Any real-world phage cocktail should also contain many different phages that are active against a variety of other common human illness-causing Salmonella serotypes and cell receptors, such as has been previously reported (Wall et al., 2010).
Further, one factor that must be addressed when selecting phages for use as an antibacterial treatment is the nature of the phages: are they lytic or temperate (lysogenic)? Lysogenic phages have the capacity to transduce bacteria by integrating into the genome and carrying DNA from one bacteria to another during excision/infection process (Brabban et al., 2005); in fact, phages were the vehicle for the introduction of the Stx genes in EHEC from Shigella (Brabban et al., 2005). Therefore, care must be taken when selecting phages to ensure that they do not display any temperate or genomically integrative characteristics, so that we do not inadvertently create an enhanced pathogen or equip them with new antibiotic resistance or virulence genes (Yoichi et al., 2004; Johnson et al., 2008). In the current study, our phages were preselected for only demonstrating the characteristics of strictly lytic phages; however, for phages to be utilized in the real world, all members of a phage cocktail must be screened under many physiological conditions for this trait.
One obvious limitation of the present study is that it utilized an artificially inoculated model to demonstrate the efficacy of a short-term phage cocktail treatment. Further, this study examined Salmonella populations in the gut after only 48–72 h of phage treatment, modeling phage use as a short-term intervention strategy for use immediately pretransport, transport, lairage, and slaughter. We used young growing swine as a model of a swine gastrointestinal tract, whereas the point of Salmonella colonization of greatest import to food safety is immediately slaughter; however, this study was designed as a proof of concept for a short-term, immediately preharvest intervention strategy. This report does not address questions about longer usage in persistently Salmonella-colonized swine.
Based upon our data, we conclude that phages show promise as a potential viable strategy to reduce Salmonella in swine before harvest. If we are to implement this technology in the human food chain, a great deal of further research needs to be performed to determine the most efficacious dosing strategies (dose and number of applications) and the most effective combinations of phages targeting the key most significant serotypes that affect humans of the diverse Salmonella population that is comprised of >2500 distinct serotypes. Phage treatment is not a panacea to control all foodborne illness; however, phage can be utilized in an integrated, multi-hurdle system aimed at reducing the passage of Salmonella from farm to fork (Callaway et al., 2007).
Footnotes
Acknowledgments
Portions of this research were supported by America's swine producers and were produced for the National Pork Board. This project was graciously funded in part by the U.S. pork producers through the National Pork Board.
Disclaimer
Proprietary or brand names are necessary to report factually on available data; however, the U.S. Department of Agriculture (USDA) neither guarantees nor warrants the standard of the product, and the use of the name by the USDA implies no approval of the product, or exclusion of others that may be suitable.
Disclosure Statement
No competing financial interests exist.