
Abstract
Many foods originate on the farm where cross-contamination with pathogens can occur, with implications for human health. This study characterized a bank of 51 Listeria monocytogenes isolates originating from 12 farms located in Ireland by pulsed-field gel electrophoresis (PFGE) to establish the molecular diversity of the isolate collection, and examine transmission patterns of L. monocytogenes across the farm environment, and also determined resistances against five different antibiotics (ampicillin, ciprofloxacin, erythromycin, penicillin G, and tetracycline). Analysis using a combination of AscI and ApaI digestion showed the 51 isolates comprised a total of 40 individual PFGE types, compared to individual restriction enzyme analysis, which was less discriminatory (36 types with ApaI analysis and 38 types with AscI analysis). Four of the PFGE types were common to multiple farms, and five farms had isolates with indistinguishable PFGE types in multiple locations on the farm. Indistinguishable PFGE types were common to multiple farms in different geographical locations up to ∼200 km apart, and were found in a variety of different sample types, indicating multiple niches for the organism in the dairy farm environment. The presence of L. monocytogenes in samples related to animals other than cattle indicated that there are multiple possible vectors of contamination. The farm environment harbors a diverse collection of L. monocytogenes isolates that must be considered as possible agents of food contamination.
Introduction
Ruminant farms have been identified as reservoirs for L. monocytogenes strains linked with human disease, particularly cattle farms (Borucki et al., 2004; Nightingale et al., 2004). To this end, studies have been carried out relating to occurrence, epidemiological characterization, and persistence of L. monocytogenes in the dairy chain, to try to improve control strategies and prevent cross-contamination of food that could lead to human infection (Borucki et al., 2005; Latorre et al., 2009; Mohammed et al., 2009). Studies have shown that L. monocytogenes can be found in a variety of sources in the farm environment (Borucki et al., 2004, 2005; Gundmundsdottir et al., 2004; Fox et al., 2009; Mohammed et al., 2009). Few studies including close examination of the molecular diversity of farm isolates, however, have been published (Borucki et al., 2004, 2005; Gundmundsdottir et al., 2004). Such studies not only allow a greater understanding of population diversity, but also enable more accurate identification of possible food contamination routes, and thus facilitate the establishment of control measures that are likely to address the problem of cross-contamination of food. Strain typing methods such as pulsed-field gel electrophoresis (PFGE) are effective for this type of analysis involving L. monocytogenes populations.
The objective of this work was to investigate the molecular diversity of 51 L. monocytogenes isolates from 12 Irish dairy farms using PFGE, to investigate the transmission patterns of L. monocytogenes on these farms. In addition, their resistances to five antibiotics (ampicillin, ciprofloxacin, erythromycin, penicillin G, and tetracycline) were determined. Isolates originating from a variety of sample locations (e.g., feces, water, grass, soil, silage, dust, bedding, and milk filter) were tested, which allowed comprehensive representation of on-farm L. monocytogenes strains.
Materials and Methods
L. monocytogenes isolates
L. monocytogenes isolates in this study were previously isolated from 12 different farms (Fox et al., 2009). One isolate from each positive sample was analyzed. Among these farms, milk produced at farms 1–3 and 5 was supplied to artisan cheese producers. Farm 4 produced cheese using milk from their own cattle herd. Farms 6–9 supplied milk, which was subsequently processed commercially for human consumption. Farms 10–12 were farms that had a cattle herd.
Serotyping of isolates
Isolates were serotyped using a combination of O-antigen antisera testing (Denka Seiken Co., Ltd.), and a multiplex serotyping PCR (Doumith et al., 2004a), as described by Fox et al. (2009). The O-antigen antisera cannot distinguish between serotypes 1/2a, 1/2b, and 1/2c, or between 3a, 3b, and 3c. The multiplex PCR was used to differentiate these serotypes.
Antibiotic testing
Antibiotic minimum inhibitory concentration (MIC) testing was carried out using M.I.C.Evaluators™ (Oxoid), as per manufacturers' instruction. Ampicillin (0.015–256 mg/L), ciprofloxacin (0.002–32 mg/L), erythromycin (0.015–256 mg/L), penicillin G (0.002–32 mg/L), and tetracycline (0.015–256 mg/L) sensitivity were tested (concentration range tested is in parenthesis). Antibiotic resistances (“sensitive,” “intermediate,”, or “resistant”) were assessed according to the Clinical and Laboratory Standards Institute guidelines.
PFGE typing and analysis
PFGE analysis was carried out using a method based on the International Standard PulseNet protocol (PulseNet USA, 2009). Isolates were initially cultured overnight on tryptic soy agar supplemented with 0.6% yeast extract (TSAYE) plates, and a 2 mL suspension in 10 mM Tris:1 mM EDTA buffer, pH 8.0 (TE), was prepared with an OD610 of 1. To 400 μL cell suspension, 20 mg/mL lysozyme was added. This was incubated for 20 min at 55°C, after which 20 μL of a 20 mg/mL Proteinase K stock was added, and 400 μL of plug agarose mix was added (1% SeaKem Gold agarose +1% sodium dodecyl sulfate in TE buffer). Plugs were prepared by dispensing the solution into plug molds, and solid plugs were then lysed overnight in 5 mL Cell Lysis Buffer (50 mM Tris:50 mM EDTA, pH 8.0 + 1% Sarcosyl), containing 25 μL of a 20 mg/mL Proteinase K solution. Plugs were then washed sequentially, twice in ultrapure water, then four times in TE buffer (solutions were prewarmed to 55°C). DNA was restricted using either AscI FastDigest (1 μL enzyme at 37°C for 2 h) (Fermentas) or 50U ApaI (30°C for 18 h) (Roche) restriction endonuclease. DNA fragments were separated on a 1% SeaKem Gold agarose gel for 21 h as described in the PulseNet protocol, on a CHEF-DR III (Bio-Rad). Gels were observed after staining in ethidium bromide solution (1 μg/mL) for 30 min. Cluster analysis of restriction profiles was performed using BioNumerics v5.10 software (Applied Maths).
Results and Discussion
PFGE typing of the 51 isolates (Lm01– Lm51), which originated on 12 different farms, was performed using ApaI and AscI digestion. Similarities were analyzed using BioNumerics software (V5.10; Applied Maths). ApaI digestion alone yielded 36 PFGE types, whereas AscI analysis alone yielded 38 PFGE types. When both restriction analysis methods were combined, 40 distinct PFGE profiles were identified (Fig. 1).
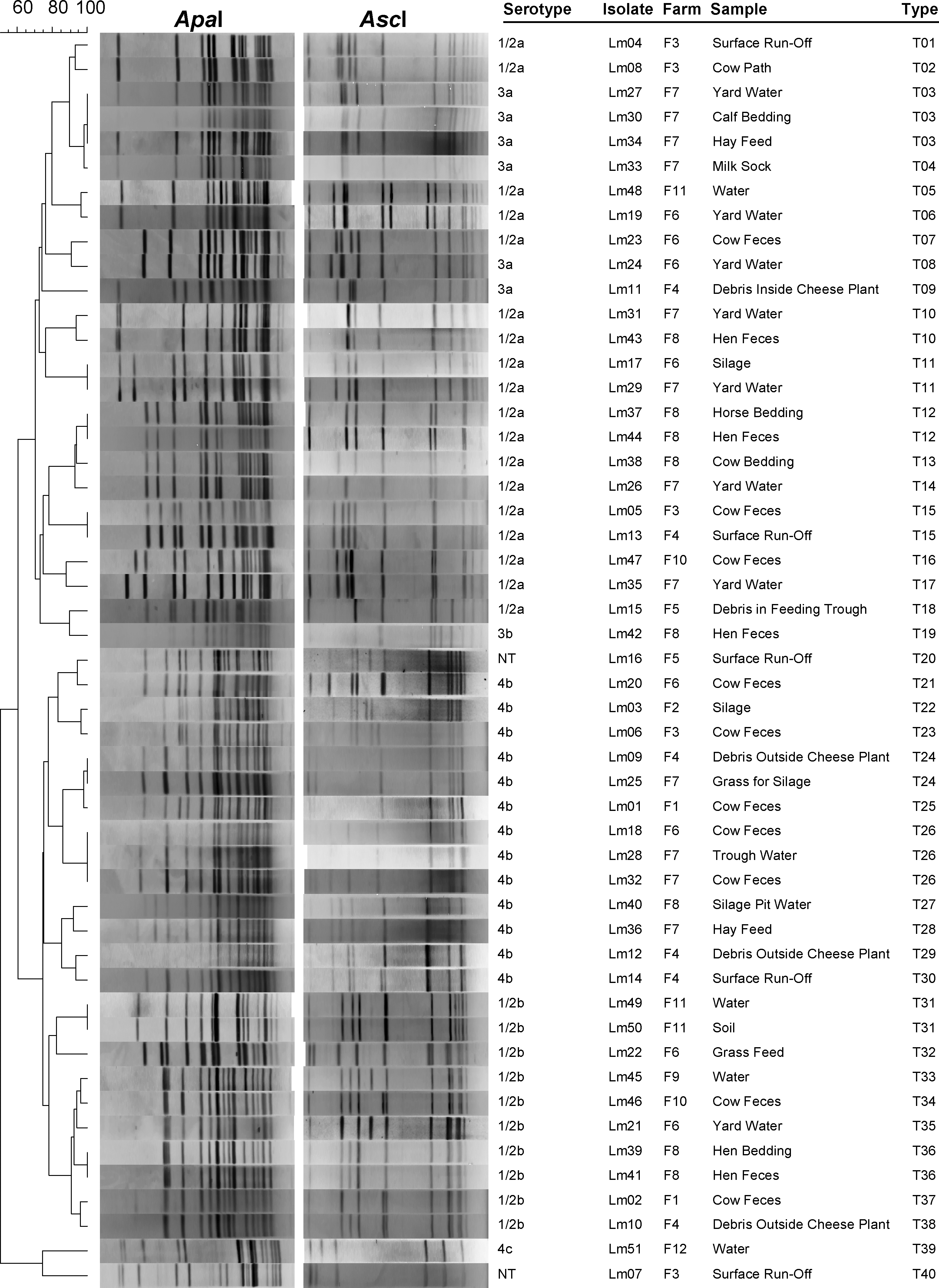
Dendogram showing pulsed-field gel electrophoresis profiles of isolates in this study, using combined ApaI and AscI restriction.
The PFGE profiling in this study showed isolates clustered largely into two main groups, corresponding to lineage I (serotype 1/2b and 4b isolates) and lineage II (serotype 1/2a and 3a isolates). Lm42 was an exception to this, as it was serotype 3b, but clustered in among the lineage II isolates. Lm07 and Lm51 clustered separately from these two main clusters. Lm07 was serotyped as 4a or 4c, and Lm51 was serotyped as 4c, which would place them among lineage III. Results indicate a majority of lineage I and lineage II isolates (94%), and few lineage III isolates. This is consistent with other studies of L. monocytogenes strain banks, such as the study by Roberts et al. (2006) on a collection of L. monocytogenes strains isolated from various different sources, although lineage III has been shown to include some 4b isolates (Ward et al., 2004). All three lineages contain isolates that have been linked with human illness, and although lineage III has been previously thought to contain predominantly strains virulent in animals (Wiedmann et al., 1997; Doumith et al., 2004b), this may in fact be due to very low lineage III food-related isolates (Roberts et al., 2006).
The percentage similarity of isolates originating from a number of different sources on Farm 7 is depicted in Figure 2. Three of the isolates isolated from different samples had the same PFGE type, T03. These isolates originated in yard water, bedding, and hay for feeding. It is possible that the cattle themselves acted as a vector for spread of this strain across the farm, as they had direct contact with all these sample sites. Five different PFGE types were isolated from yard water samples, indicating that there is potential for yard water to be contaminated with strains from many different sources. This water is accessible to all domestic animals and wildlife on the farm, and could also be contaminated by farm personnel. The presence of T04 on this farm, which appears to be a subtype of T03 (or vice versa), indicates that native strains may also give rise to mutant subtypes (milk from this farm was subsequently processed into milk for human consumption). Such genetic events may lead to increased virulence potential, and a high-prevalence strain may increase this possibility, as such a strain, found in a variety of niches in the environment, would have a greater chance of exposure to horizontal gene transfer, or acquisition of foreign DNA. L. monocytogenes possesses different types of internalin genes, and horizontal gene transfer is thought to be responsible for this gene diversity (Kelly et al., 2009). Indeed, a study into the aquisition of tet(M) in tetracycline-resistant Listeria strains by Bertrand et al. (2005) concluded that successive transfers between other Gram-positive strains may be responsible.

Listeria monocytogenes ecology on Farm 7, based on pulsed-field gel electrophoresis analysis.
Four of the PFGE types (i.e., T11, T15, T24, and T26) were found on more than one farm in the study (Fig. 1). In the case of T24 there was a distance of roughly 200 km between the two farms on which this strain was isolated. No direct connection between the farms is known (staff, suppliers, etc.). All four PFGE types were found in different sample matrices, indicating that strains can occur, and possibly survive and multiply, in a multitude of niches on the farm. This is also supported by results relating to types T03, T12, T26, T31, and T36, which were found in different samples on the same farm. Thus, the findings of this study are consistent with the fact that L. monocytogenes is known to be ubiquitous in the environment. The bacterium is also capable of persisting in the farm environment, and Borucki et al. (2005) found a high prevalence of isolates of the same PFGE type in multiple samples taken 9 months apart. This is very important in terms of risk to human health, as farms supplying products to the food industry risk introducing potentially virulent strains into the human food chain. Indeed, in a separate study by Borucki et al. (2004), dairy farms were shown to be reservoirs for strains implicated in sporadic and epidemic cases of human listeriosis. In the case of T24, there was 12 months between isolation of Lm09 and Lm25. Although neither T24 isolate originated from the same farm, it is possible that some of the isolates characterized in this study may in fact persist at the locations in which they were found. T03, comprising samples Lm27, Lm30, and Lm34, all from Farm 7, were isolated each month over 3 months of sampling. It is unclear whether such persistence is a result of adaptation of the strain to survival in its niche farm environment, is due to a series of recontamination events, or is the result of a life-cycle involving, for example, re-infection and shedding by cattle. Oliver et al. (2005) propose a cycle by which foodborne and veterinary pathogens may persist in a dairy farm environment, and this may well apply to L. monocytogenes.
The main source of L. monocytogenes infection on the farm is generally thought to be poor quality silage feed (Borucki et al., 2005). T24 was found in cut grass, which was subsequently to be processed into silage. The same PFGE type was found in debris outside the cheese production facility at another farm. Other silage, or silage-related samples were also found to be positive: Lm03, Lm17, Lm31, and Lm40. As there is potential for grass to be contaminated with L. monocytogenes from a variety of sources, it is important that silage stores on farms are maintained under conditions that limit the potential growth of L. monocytogenes, including low pH and high lactic acid concentration (Pauly and Tham, 2003).
In the case of Farm 8, a feces sample taken from the hen flock and the straw bedding in the hen coop yielded L. monocytogenes isolates with the same PFGE profile (T36). T12 was found in both hen feces and a horse bedding sample. T19 was also found in hen feces. These results suggest that other animals on the farm, apart from the cattle, may contribute to the spread of L. monocytogenes in the farm environment, in particular by fecal shedding. In the case of Farm 8, the hens had freedom of movement around the entire farm (including the dairy area), which represents a risk for spread of L. monocytogenes strains throughout the farm. The family living here consumed raw milk from their herd, indicating an increased risk of exposure to L. monocytogenes.
Antibiotic testing was carried out on all the isolates using 5 different antibiotics: ampicillin, ciprofloxacin, erythromycin, penicillin G, and tetracycline. β-lactams are generally the antibiotic of choice for treatment of human listeriosis cases, with ampicillin and penicillin G often prescribed (Hof et al., 1997). In the case of allergy to these, a macrolide such as erythromycin is an alternative used (Hof, 1991). Fluoroquinolones, such as ciprofloxacin, are generally used to treat Gram-negative bacteria, and as such are not generally associated with treatment of L. monocytogenes infection (Hof et al., 1997). However, it was included to examine the effectiveness of a nontypical L. monocytogenes treatment. As with erythromycin, tetracycline was initially one of the treatments of choice for treating listeriosis, in the case of patient allergy to ampicillin or penicillin G (Hof, 1991). Spontaneous or acquired resistance to tetracycline, however, has been found in L. monocytogenes isolates (Schmitt-Slomska et al., 1982; Poyart-Salmeron et al., 1992).
Susceptibility to the antibiotics tested in this study were assessed according to the Clinical and Laboratory Standards Institute guidelines, and was defined as having an MIC below the following: ampicillin MIC ≤2 mg/L; ciprofloxacin MIC ≤1 mg/L; erythromycin MIC ≤0.5 mg/L; penicillin G MIC ≤2 mg/L; tetracycline MIC ≤4 mg/L. MICs for all 51 isolates are listed in Table 1. None of the isolates in this study were resistant to any of the five antibiotics tested. There is immense variation in results of antibiotic resistance studies of L. monocytogenes. As was the case with this study, there are reports where no antibiotic resistance among banks of isolates tested was found. A screen by Mayrhofer et al. (2004) included erythromycin, penicillin, and tetracycline, and no resistance was found among all 60 L. monocytogenes isolates tested. Chen et al. (2010) also found no resistance to ampicillin, erythromycin, and tetracycline among the 86 L. monocytogenes isolates they tested. A study by Walsh et al. (2001) assessed resistance of isolates from retail foods in Ireland, which included ampicillin, erythromycin, penicillin G, and tetracycline screening. No resistance was found among the 351 isolates tested to ampicillin, erythromycin, or penicillin G. Two of the isolates, however, showed resistance to tetracycline. In contrast to these studies, Ayaz and Erol (2010) found high levels of resistance to antibiotics among the 78 isolates they tested. No isolates were resistant to tetracycline; however, 41% of the isolates showed intermediate resistance to erythromycin, and 69% and 81% of the isolates were resistant to ampicillin and penicillin G, respectively. Although pathogenic potential of these isolates in humans is unknown, these results are positive from an Irish public health perspective. Overuse of antimicrobials in the agriculture sector in recent years has been a major cause of concern, as development of antimicrobial resistance among dairy-related pathogens has implications for their treatment in cases of human infection (Oliver et al., 2011). Absence of antimicrobial resistance among isolates in this study may indicate correct and responsible use of antimicrobials on farms included in this study; however, no data were taken on herd exposure to antibiotic/antimicrobial treatments, and without further isolate screening and genome analysis the reason for this susceptibility is difficult to postulate.
Antibiotics tested were (concentration range tested is in parenthesis) ampicillin (0.015–256 mg/L), ciprofloxacin (0.002–32 mg/L), erythromycin (0.015–256 mg/L), penicillin G (0.002–32 mg/L), and tetracycline (0.015–256 mg/L).
Conclusions
The results of this study show that indistinguishable L. monocytogenes PFGE types are present on multiple farm locations, spread throughout the South-West of Ireland. Although AscI typing differentiated isolates more than ApaI typing (38 PFGE types as opposed to 36, respectively), the combination of both AscI and ApaI PFGE typing gave the greatest isolate differentiation, with 40 different types. Strategies to prevent cross-contamination of food with farm-associated strains of L. monocytogenes are an important component of limiting the spread of this pathogen in the food chain. None of the isolates tested in this study were resistant to any of the five antibiotics tested, indicating a positive potential prognosis for any associated human listeriosis that may occur.
Footnotes
Acknowledgments
This work was supported by the EU 6th Framework Program through the project BIOTRACER (project no. 036272) and the Teagasc Walsh Fellowship Scheme.
Disclosure Statement
No competing financial interests exist.