
Abstract
The objective of the present study was to examine whether the expression of qnrA may contribute to a high level of resistance among parent and induced strains of Kluyvera spp. Two clinical isolates of ciprofloxacin-resistant Kluyvera spp. were obtained from livers of diseased chickens, and upon induction with ciprofloxacin, six strains with increased resistance were produced. Point mutations in qnrA, aac(6')-Ib-cr, gyrA, gyrB, parC, and parE were investigated by polymerase chain reaction (PCR) amplification and DNA sequencing, and expression levels of acrAB and qnrA in all strains were investigated by quantitative real-time PCR (qRT-PCR). The induced strains contained the same mutations in quinolone resistance-determining region as those of the parent strains. qRT-PCR showed that the expression of the acrA gene was not detected in any strain and acrB gene expression was unchanged between induced and parental strains. However, difference in expression of qnrA was observed, which correlated well with the level of quinolone resistance in the parent and induced strains. The induced high resistance was not affected by mutations in qnrA and aac(6')-Ib-cr, by new mutations in the quinolone resistance-determining region of gyrA, gyrB, parC, and parE, or by the expression level of acrAB. These data suggest that the expression of qnrA may be a factor contributing to the high level of resistance among parent and induced strains of Kluyvera spp.
Introduction
Fluoroquinolones are widely used in clinical medicine; however, bacterial resistance has become an increasing problem, limiting the number of agents available for treatment of infections (Lu et al., 2011). Mechanisms of bacterial resistance to fluoroquinolones include (1) accumulation of mutations in bacterial enzymes targeted by fluoroquinolones (Robicsek et al., 2006a), (2) changes in expression of efflux pumps, which causes accumulation of these agents in the bacterial cell to decrease (Poole, 2000), and (3) plasmid-mediated resistance, mediated by the qnrA gene (Martínez-Martínez et al., 1998) and other related genes.
Mutations in the target enzymes of the quinolone, namely DNA gyrase and topoisomerase IV, are important mechanisms of quinolone resistance. The majority of mutations described to date have been found within a region called the quinolone resistance-determining region (QRDR). Mutations in this active region may alter binding of quinolone to the site and lead to reduced susceptibility or resistance to quinolone (Willmott and Maxwell, 1993). On the other hand, MarA-induced upregulation of acrAB, encoding a stress-induced efflux pump (Ma et al., 1993, 1995), leads to decreased intracellular drug accumulation. Studies to date suggest that the acrAB efflux pump plays a major role in the resistance phenotype mediated by the mar locus. AcrA was overexpressed in 9 of 10 high-level fluoroquinolone-resistant (minimal inhibitory concentrations [MICs] ≥32 μg/mL) clinical isolates of Escherichia coli (Mazzariol et al., 2000), and acrAB deletion mutants were hypersusceptible to all antimicrobials tested.
PMQR has been reported recently. The gene responsible for PMQR has been identified as qnr (Martínez-Martínez et al., 1998). It belongs to the pentapeptide repeat family that protects DNA gyrase and topoisomerase IV activities in vitro by reducing the number of holoenzyme–DNA targets for quinolone inhibition (Tran and Jacoby, 2002; Wang et al., 2003; Tran et al., 2005). PMQR could be horizontally spread, the spectrum of resistance included fluoroquinolone as well as nalidixic acid, and resistance could be transferred serially to successive recipients in all strains of E. coli, Klebsiella pneumoniae, Citrobacter freundii, Salmonella typhimurium, and Pseudomonas aeruginosa (Martínez-Martínez et al., 1998). It was thought that the clinical importance of qnrA resides in facilitating selection of higher resistance, raising the level at which quinolone exposure would be lethal to the cell. But, beyond this, little is known about the clinical importance of the qnrA gene. Recently, other PMQR genes, namely qnrB, qnrS, qnrC, qnrD, and qepA, have been also described on transferable plasmids (Hata et al., 2005; Jacoby et al., 2006; Yamane et al., 2007; Cavaco et al., 2009; Wang et al., 2009). Another type of PMQR determinant has been discovered in qnrA-positive E. coli, named aac(6')-Ib-cr gene; this variant possesses two substitutions at codons Trp102Arg and Asp179Tyr, compared with the wild-type aac(6')-Ib enzyme (LinksPark et al., 2006; Robicsek et al., 2006b). Consequently, the variant enzyme acetylates ciprofloxacin and norfloxacin, conferring slightly higher MICs (a two- to fourfold increase) (Barco et al., 2010).
In this study, two clonally unrelated clinical isolates of Kluyvera spp. carrying qnrA and their derived higher-level ciprofloxacin-resistant induced strains were investigated. Target genes (gyrA, gyrB, parC, and parE), qnrA, and aac (6')-Ib-cr were examined, and the transcription levels of acrAB and qnrA were studied by real-time fluorescent quantitative polymerase chain reaction (qRT-PCR) to determine the association of qnrA expression with quinolone resistance.
Materials and Methods
Bacterial strains, identification, conjugation, and MIC determination
Two Gram-negative bacteria were isolated from livers of chickens with the symptom of hepatomegalia from a farm in Sichuan Province, China. All bacteria were routinely grown at 37°C in Luria-Bertani (LB) broth or on Mueller–Hinton agar (MHA) plates (Xia et al., 2010). Conjugation experiments were carried out, as previously described (Wang et al., 2004), in LB broth with E. coli J53AzR as the recipient to determine whether quinolone resistance was transferable in strains containing qnrA-bearing plasmids. Transconjugants were selected on trypticase soy agar plates containing sodium azide (100 μg/mL) for counter selection and chloramphenicol (50 μg/mL) to select for plasmid-encoded resistance. Ciprofloxacin MICs of isolates, induced strains, and transconjugants were determined using agar dilution in accordance with CLSI guidelines (CLSI, 2004, 2005; Huang et al., 2009).
Selection of high-level ciprofloxacin-resistant induced strains in vitro
Two ciprofloxacin-resistant isolates (MIC=32 μg/mL) that carried qnrA were used as parent strains for induction of high level of ciprofloxacin resistance. A single colony of each strain was isolated from Mueller–Hinton agar plates and incubated in LB broth at 37°C for 12 h. The LB broth cultures (105 colony forming units/mL) contained ciprofloxacin at 16, 32, and 64 μg/mL and were incubated in 96-well cell-cluster plates (Corning Incorporated) for 20 h. The well that showed growth at the highest ciprofloxacin concentration was used as the MIC of the following generation. The procedure was then repeated with elevated ciprofloxacin concentrations (levels equal to 0.5×, 1×, and 2× MIC) until high-level ciprofloxacin-resistant strains (MIC=512 μg/mL) were generated.
Sequencing of qnrA, aac(6′)-Ib-cr, and QRDR of gyrA, gyrB, parC, and parE
PCR amplifications were carried out with the TaKaRa LA PCR kit, and incubation conditions were those suggested by the manufacturer (Xu et al., 2007). Primers (Table 1) were used to amplify qnrA, gyrA, gyrB, parC, parE, and aac(6')-Ib-cr. Purified PCR products were sequenced on both strands, and DNA sequences of qnrA and aac(6')-Ib-cr and QRDR of gyrA, gyrB, parC, and parE were compared with those available in GenBank (AY070235, EU543272, and NC000913, respectively).
Primers used for amplification and sequencing.
Primers used to carry out real-time polymerase chain reaction.
RNA extraction and cDNA synthesis
Total RNA was prepared using Trizol agent (Invitrogen) and treated with DNase I (RNase-free; Takara) according to the manufacturer's instructions. RNA purity and content were determined using a UV spectrophotometer (Thermo Spectronic) at 260 and 280 nm. Avian myeloblastosis virus reverse transcriptases (Takara) were used to synthesize cDNA from RNA samples. A total of 1.0 μg RNA was added in a 50 μL reverse transcription (RT) reaction mixture, and incubation conditions used were according to the manufacturer: 25°C for 10 min, 42°C for 60 min, and 83°C for 1 min.
Generation of standard curves for qRT-PCR
Fragments of acrA, acrB, qnrA, and 16S rRNA genes were 175, 183, 182, and 186 bp (primers listed in Table 1), respectively, and were amplified from a standard strain by PCR. The PCR products were purified with the gel purification kit (Qiagen) according to manufacturer's instructions and ligated into the pGEM-T Easy vector (Promega). Escherichia coli DH5α competent cells were transformed with the plasmids and positive clones were selected by PCR. Plasmids containing DNA of interest were extracted with the Mini Plasmid Kit (Qiagen). Plasmids at 10-fold serial dilutions were amplified and used to construct standard curves.
Real-time fluorescent qRT-PCR
Real-time PCR was performed using SYBR green PCR master mix (Applied Biosystems) and carried out using the DNA Engine® Chromo 4™ real-time PCR Detector (Peltier Thermal Cycler; Bio-Rad). cDNA reaction products (2 μL) were used per 25 μL in real-time PCR employing SYBR green detection. All reactions were performed in tetraplicate. Amplification of an endogenous control, the 16S rRNA gene, was performed to standardize the amount of sample RNA or DNA added to a reaction. Primer sequences used for each target gene are shown in Table 1. PCR cycling conditions were 1 cycle at 95°C for 5 min, followed by 30 cycles at 95°C for 30 sec, 57°C for 30 sec, and 72°C for 1 min, and 1 sec plate reading at 80°C; melting curve was read from 65°C to 95°C on every 0.2°C and the final extension was for 10 min at 72°C. Results were analyzed with the MJ Opticon Monitor analysis software. Relative quantification was determined by the 2–ΔΔCT or ΔΔC T method. Expression of the endogenous control gene, 16S rRNA, was used to normalize the data.
Results
Identification of the qnrA gene and aac(6′)-Ib-cr
Two Gram-negative bacteria were collected from a farm in Sichuan Province, China, and they contained the aac(6')-Ib-cr and qnrA gene (detected by PCR assays and confirmed by sequencing). The API 20E system (bioMerieux; Marcy l'Etoile) used for identification revealed that both of them were Kluyvera spp. Strains, named L10 and L19, respectively. The qnrA genes were cloned and sequenced. DNA sequencing showed that Kluyvera strains L10 and L19 had 95.6% (628/657) similarity in nucleotides and 98.6% (215/218) similarity in amino acids with those of the originally reported qnrA (known as qnrA1) (Martínez-Martínez et al., 1998). Quinolone resistance in L10 and L19 could be transferred by conjugation, and the ciprofloxacin MICs of the transconjugants were 0.5 μg/mL.
Sequencing of qnrA and QRDR of gyrA, gyrB, parC, and parE
The qnrA genes in induced strains with different levels of ciprofloxacin resistance were amplified and sequenced to detect mutations. No mutation in qnrA was observed in induced strains with different levels of ciprofloxacin resistance (Table 2). The gyrA and parC in the isolates (L10 and L19) and the induced strains with different levels of ciprofloxacin resistance were cloned and sequenced to detect mutations. Three mutations (S83I, I112V, and S171A) in gyrA emerged in L10, three mutations (S83I, I112V, and K154R) in gyrA emerged in L19, and a single mutation (S80I) in parC was detected in L10 and L19. These were the same mutations detected in the L10 or L19 induced strains with different levels of ciprofloxacin resistance (Table 2). No mutations were detected in gyrB and parE in either induced strains or parents.
L10-2, L10-3, and L10-4: different levels of ciprofloxacin-resistant induced strains that were derived from L10.
L19-2, L19-3, and L19-4: different levels of ciprofloxacin-resistant induced strains that were derived from L19.
MIC, minimum inhibitory concentration; CIP, ciprofloxacin.
Detection of expression levels of acrAB and qnrA
We hypothesized that the transcription level of acrAB and qnrA may be related to the level of ciprofloxacin resistance. The study was performed by incubating strains L10 and L19 in LB broth containing 32 μg of ciprofloxacin per mL and incubating strains L10-2 and L19-2, L10-3 and L19-3, and L10-4 and L19-4 in LB broth containing 128, 256, and 512 μg of ciprofloxacin per mL, and the expression level of acrAB and qnrA was tested using the total RNA harvested from the cells during midlogarithmic growth phase. Expression of acrA was not detected in any of the strains. There was also no significant difference in the expression levels of acrB in the induced strains compared with their parent strains (Table 2). Conversely, we found that expression levels of qnrA in strains with the same level of resistance were similar, and the number of qnrA transcripts in both strains (L10 and L19) increased along with increasing levels of ciprofloxacin resistance (Fig. 1, A and B). The greater abundance of the qnrA transcripts was observed in induced strains with higher level ciprofloxacin resistance (p<0.01). This demonstrates that the qnrA transcript is a factor relating to ciprofloxacin resistance.
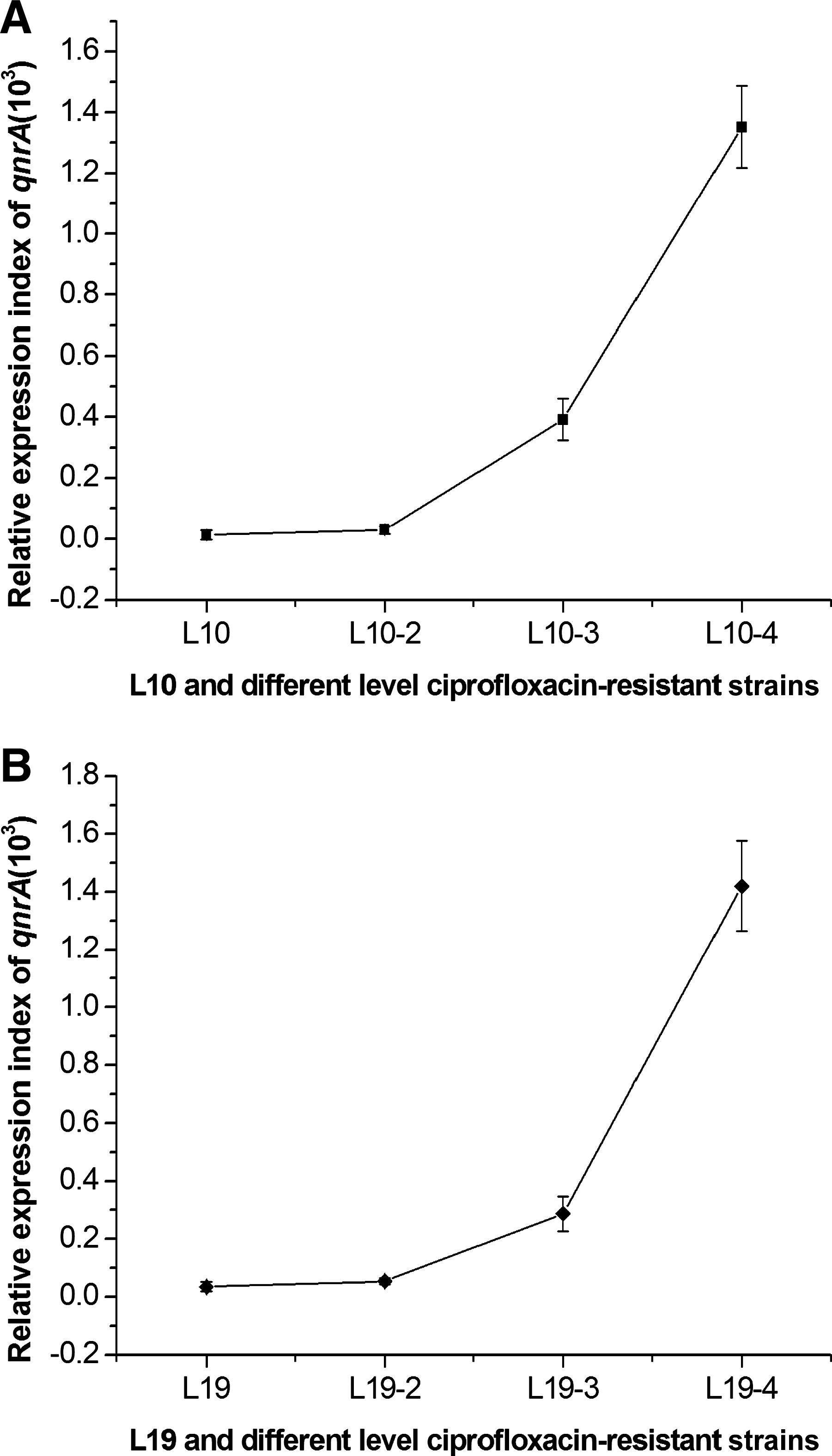
Expression levels of the qnrA gene in different strains:
Discussion
Kluyvera spp. have a more pathogenic nature than previously believed and have been isolated from many sources, including sputum, urine, stool, throat, and blood specimens, sewage, soil, kitchen food, water, hospital sinks, and shell eggs (Farmer et al., 1981; Musgrove et al., 2008; Toprak et al., 2008). In pediatric patients, Kluyvera spp. has emerged as a cause of diseases ranging from soft tissue infections to sepsis with multiorgan failure (Carter et al., 2008). These findings suggested that Kluyvera spp. is wildly distributed and becoming more important.
In this study, we detected two isolates of Kluyvera in diseased chickens, with PMQR genes. As a foodborne pathogen, Kluyvera could also transfer resistant genes in vivo between Enterobacteriaceae (Lascols et al., 2008). Therefore, clinicians should be aware of their virulence and potential resistance to many antimicrobials such as fluoroquinolones.
The causes for antimicrobial resistance are multifactorial. It has been well documented that resistance in general is caused by overuse of antibacterial agents (Li et al., 2010). To improve our understanding of fluoroquinolone resistance, we investigated the mechanisms of fluoroquinolone resistance in induced strains with a high level of ciprofloxacin resistance selected from Kluyvera spp. isolated from chickens, which contained the qnrA gene.
Quinolone can inhibit DNA gyrase by stabilizing the complex between DNA and DNA gyrase, thus blocking the progression of polymerase. Tran and Jacoby (2000) reported that QnrA encoded by qnrA, the novel quinolone resistance plasmid gene, can specifically bind to DNA gyrase holoenzyme and protect E. coli DNA gyrase from inhibition by ciprofloxacin. We examined the qnrA gene of strains with increased levels of ciprofloxacin resistance to determine whether qnrA contained new mutations that could cause an increase or decrease in the activity of QnrA. Our results, however, revealed no changes in the qnrA gene when compared with the parent strains with lower resistance to ciprofloxacin, indicating the gene is stable. Cattoir et al. (2007) also reported that random and site-directed mutagenesis failed to identify any single amino acid substitution in qnrA and qnrS that could be responsible for higher level of resistance to quinolone or fluoroquinolone. Quinolone resistance in Gram-negative bacteria is usually caused by mutations in chromosomal genes coding for type II topoisomerases (Hopkins et al., 2005). Our results revealed that both strains L10 and L19 contained three mutations in gyrA and one mutation in parC (Table 2). No new mutation was detected in induced strains with a high level of ciprofloxacin resistance, suggesting that the high level of ciprofloxacin resistance in the induced strains was not caused by new mutation in the QRDR of gyrA and parC.
aac(6')-Ib-cr, a variant of aac(6')-Ib, is another PMQR gene that encodes an aminoglycoside acetyltransferase that confers reduced susceptibilty to ciprofloxacin by N-acetylation of its piperazinyl amine. aac(6')-Ib-cr has two amino acid changes, Trp102Arg and Asp179Tyr, which together are necessary and sufficient for the enzyme's ability to acetylate ciprofloxacin (Hopkins et al., 2005). However, the aac(6')-Ib-cr gene already existed in the original strain prior to our mutation study. Therefore, the developed high level of ciprofloxacin resistance in these induced strains could not be attributed to the aac(6')-Ib-cr gene.
The presence of efflux pumps might explain different MICs for the clinical isolates and their induced strains. There are seven resistance-nodulation cell-division transporters, five of which, namely AcrAB, AcrEF, AcrD, YhiUV, and MdtABC, have been functionally characterized and confirmed to participate in drug efflux. The AcrAB has been identified as the predominant drug efflux pump (Sulavik et al., 2001), and it is the best characterized resistance-nodulation cell-division pump to date. Knockout experiments with acrEF, yhiUV, and mdtABCD did not change drug susceptibilities of wild-type cells, suggesting that these pumps did not play a significant role in the antimicrobial resistance. In this study, acrA was absent in all strains and no difference in the expression of acrB was observed in any of the strains with increased ciprofloxacin resistance (Table 2). Therefore, elevated MICs in induced strains were probably not affected by the expression level of these efflux pumps.
To determine whether elevated MICs observed for the induced strains were caused by differential transcription of the qnrA gene, the number of qnrA-specific transcripts was compared by qRT-PCR. It was surprising that the expression level correlated very well with the MICs of the isolates and their induced strains. The relation between ciprofloxacin resistance and the expression level of qnrA gene was assessed by calculating Spearman's correlation coefficient and the corresponding P value (r s=1, p<0.01). The higher basal expression of qnrA in induced strains may explain, at least in part, the higher MICs of quinolone for these induced strains. Rodríguez-Martínez et al. (2006) also reported that the expression of the qnrA gene correlated well with the level of quinolone resistance in E. coli transconjugants, and the main factor determining the resistance level in transconjugants was the different levels of qnrA expression. Similarly, we hypothesized that a high level of expression of the qnrA gene could be one of the factors responsible for the high level of ciprofloxacin resistance. In this study, we chose a standard strain that did not contain the qnrA gene as a control, we obtained induced strains with a high level of fluoroquinolone resistance from the L10 and L19 parents, but the same strain of standard strain was difficult to obtain (data not shown). However, an increase in qnrA in the presence of high concentrations of ciprofloxacin makes sense for bacterial survival. A higher number of qnrA transcripts could provide the cell with a rapid response by protecting DNA gyrase and the topoisomerases from the damaging effects of antimicrobial agents that inhibit DNA replication. We found that strains containing the qnrA gene could have easily survived under the pressure of ciprofloxacin, compared with those without the qnrA gene. Similar to the high resistance levels observed for nalidixic acid, overuse of fluoroquinolone may increase the current fluoroquinolone breakpoints.
In conclusion, the level of qnrA transcription correlated well with MICs of ciprofloxacin, but the expression level of acrB did not change in these Kluyvera spp. induced strains. Therefore, it can be proposed that the main factor influencing resistance in these induced strains was the expression of qnrA, especially as no further mutation was detected in qnrA, gyrA, gyrB, parC, parE, and aac(6')-Ib-cr and the acrA gene was absent. As increased expression was not associated with any mutations within the open reading frame, future studies will focus on characterizing the promoter regions of the different isolates to determine the reason and/or whether other plasmid factors affect the expression of qnrA. Our results suggest that the expression of the qnrA gene is a factor relating to high fluoroquinolone resistance. In this study, only two isolates were included, and in the future, much more research is needed to determine whether these findings are at all common.
Footnotes
Acknowledgments
The authors thank George A. Jacoby, David C. Hooper, and Minggui Wang for kindly providing the E. coli strains J53AzR, V517, R1, R27, and plac. This study was supported by a grant from the Program for Cheung Kong Scholars and the Innovative Research Team at the University of China (No. IRT0866) and Exclusive Research Fund for Public Welfare from Ministry of Agriculture of the People's of China (No. 200903055).
Disclosure Statement
No competing financial interests exist.