
Abstract
Yersinia enterocolitica is a zoonotic agent that causes gastrointestinal disease in humans, as well as reactive arthritis and erythema nodosum. Enteropathogenic Yersinia are the etiological agents for yersiniosis, which can be acquired through the consumption of contaminated foods. As porcine animals are the main carriers of Y. enterocolitica, food safety measures to minimize human infection are of increasing interest to the scientific and medical community. In this review, we examine why it is imperative that information on the reservoirs, prevalence, virulence, and ability of this pathogen to survive in different environments is further investigated to provide rational measures to prevent or decrease associated disease risks.
Introduction
According to Bergey's Manual of Systematic Bacteriology, the genus Yersiniae belongs to the phylum Proteobacteria, class Gammaproteobacteria, order Enterobacteriales, family Enterobacteriaceae (Bottone, 1997). Yersinia are small rod-shaped, Gram-negative coccobacilli often isolated from clinical specimens, including wounds, feces, sputum, and mesenteric lymph nodes. They are not part of the normal human flora. The type species, Y. pestis, is known to be a specialized clone of Y. pseudotuberculosis (Achtman et al., 1999; Morelli et al., 2010). Yersinia species comprise a biochemically heterogeneous group that can grow at refrigeration temperatures of 4°C. Based on their biochemical heterogeneity and DNA relatedness, Y. enterocolitica-like organisms were separated into four species: Y. enterocolitica, Y. intermedia, Y. frederiksenii, and Y. kristensenii (Bercovier et al., 1980; Bottone, 1997). Following additional revisions, the genus Yersinia now includes 17 species (Euzéby, 1997; Murros-Kontiainen et al., 2011a,b) in addition to the three recognized human pathogens Y. pestis, Y. pseudotuberculosis, and Y. enterocolitica: these are Y. aldovae, Y. aleksiciae, Y. bercovieri, Y. enteromophaga, Y. frederiksenii, Y. intermedia, Y. kristensenii, Y. massiliensis, Y. mollaretti, Y. nurmii, Y. pekkanenii, Y. rohdei, Y. ruckeri, and Y. similis. Y. pseudotuberculosis and Y. enterocolitica cause mesenteric adenitis and terminal ileitis in humans. The latter are responsible for the greatest number of cases of zoonotic illness among all Yersinia species, being mainly associated with foodborne illnesses (Bercovier et al., 1984; Tauxe, 2002; Jalava et al., 2004; Martínez et al., 2009; Okwori et al., 2009).
Y. enterocolitica and Y. pseudotuberculosis can be isolated from pigs, birds, beavers, cats, and dogs (Murphy et al., 2010), and Y. enterocolitica has also been detected in environmental sources, including ponds and lakes, along with food sources such as meats, ice cream, and milk (Greenwood et al., 1990; FDA, 2005). Most isolates from these sources do not belong to recognized pathogenic bioserotypes. Y. pseudotuberculosis and Y. enterocolitica are well-established causative agents of foodborne gastroenteritis (Tacket et al., 1984; Tauxe et al., 1987; Nuorti et al., 2004).
Yersinia enterocolitica
Y. enterocolitica is a psychotrophic and facultative anaerobic bacterium that occurs widely in nature (Bottone, 1997; Robins-Browne, 2007). It is classified using biochemical characteristics along with serotyping. There are six major biotypes designated as 1A (the non-pathogenic type), and there are the pathogenic biotypes 1B, 2, 3, 4, and 5 (Table 1) (Wauters et al., 1987). Strains of biotypes 1B through 5 can carry a virulence plasmid, denoted as pYV, and several well-characterized chromosomally encoded virulence determinants (Table 2). However, growing epidemiological, clinical, and experimental evidence now suggests that a minority of biotype 1A strains are virulent and can cause gastrointestinal disease, although no consistent genetic or biochemical marker has been demonstrated to identify these strains (Grant et al., 1998; Tennant et al., 2003; Tennant et al., 2005; Batzilla et al., 2011). Y. enterocolitica is subdivided into Y. enterocolitica subspecies enterocolitica consisting of biotype 1B, which are highly pathogenic and are commonly termed the North American strains (Neubauer et al., 2000; Howard et al., 2006), and Y. enterocolitica subspecies palearctica, which includes biotypes 1A, 2, 3, 4 and 5, all of which have been isolated worldwide. Y. enterocolitica and related species can be separated serologically into groups based on their heat-stable somatic antigens. In 1981, there were 54 serogroups for Y. enterocolitica and related species recognized (Wauters, 1981). This schema was subsequently simplified by Aleksic and colleagues in 1987 to include 18 serogroups within the Y. enterocolitica species (Aleksic et al., 1987). Currently, pathogenic strains belonging to serotypes O:1, 2, 3; O:2,3; O:3; O:8; O:9; O:4,32; O:5,27; O:13a,13b; O:18; O:20; and O:21 have been identified (Table 1). Strains of biotype 1B belong to a small number of pathogenic serotypes, with the most common serogroup being O:8. Biotype 2 only includes two serotypes, O:9 and O:5,27, which are pathogenic to humans. Strains of biotype 4 serotype O:3 constitute the main pathogenic bioserotype associated with human illness (McNally et al., 2004; Nesbakken, 2005; Rosner et al., 2010). Bioserotypes that predominate in human illness include 4/O:3, 1B/O:8, 2/O:9, and 2/O:5,27.
NT, not typable. Serotypes which include strains considered to be primary pathogens are shown in bold (modified from Robins-Browne, 2007 and Wauters et al., 1987).
v, becomes active at this temperature.
Pathogenicity of Y. enterocolitica
Some strains of Yersinia invade mammalian cells such as HeLa cells in tissue culture (Bottone, 1997; Lee et al., 1977). Strains that lose their virulence properties following passaging in vitro maintain their ability to invade HeLa cells, as the invasive phenotype is encoded by chromosomal loci.
Two chromosomal genes in Y. enterocolitica involved in mammalian cell invasion have been identified: inv (Invasin) and ail (Attachment-Invasion Locus) (Miller and Falkow, 1988; Miller et al., 1989). Miller and Falkow (1988) demonstrated that the transfer of these genes into Escherichia coli confers the invasive phenotype on this bacterial host. The inv gene facilitates high-level Yersinia invasion of several tissue culture cell lines (Miller and Falkow, 1988). However, Southern blot analyses showed that inv gene sequences are present in both invasive and non-invasive isolates (Miller et al., 1989; Robins-Browne et al., 1989). Although this suggests that the inv gene alone in Y. enterocolitica may not be directly linked with invasiveness, genetic evidence shows that inv genes are non-functional in the non-invasive isolates (Pierson and Falkow, 1990). Invasin is a non-fimbrial adhesin that binds β1 integrins, is expressed on the apical surfaces of M cells (Isberg and Barnes, 2001), and facilitates invasion of Peyer's patches, in conjunction with other factors.
According to Miller and Falkow in 1988, the ail gene showed greater host specificity with regard to in vitro cell invasion, and it appeared to be present only in pathogenic Yersinia (Table 2). However, recent studies have shown the presence of this gene in Yersinia species thought to be non-pathogenic (Cheyne et al., 2010). Although it is unrelated to inv, it also affords Y. enterocolitica the ability to invade cells in culture. The latter marker works together with a 70-kb virulence plasmid pYV-encoded protein, YadA, to invade host cells promoting resistance to serum by binding host complement regulators (Miller et al., 2001; Biedzka-Sarek et al., 2005; Kirjavainen et al., 2008). Y. enterocolitica isolates have previously been shown to be tissue culture—invasive, and all isolates carried the ail marker (Portnoy et al., 1981; Miller and Falkow, 1988). The latter contributes to serum resistance and invasion in vitro (Miller et al., 1989; Wachtel and Miller, 1995).
All pathogenic strains of Y. enterocolitica biotypes 1B through 5 carry the pYV plasmid. It specifies several thermo-dependent properties, including the secretion of 12 plasmid-encoded proteins termed Yops (
Strains of Y. enterocolitica, belonging to biotype 1B, possess a chromosomal high pathogenicity island (HPI). This encodes an additional siderophore (yersiniabactin) synthesis apparatus and siderophore reuptake mechanism allowing the pathogen to acquire iron from normally inaccessible sources in the host. Yersiniabactin also reduces production of reactive oxygen species (ROS) by innate immune effector cells (Paauw et al., 2009), thereby reducing bacterial killing. Although encoded by an integrative and conjugative element (ICE) (Schubert et al., 2004), which can horizontally spread among Enterobacteriaceae, including E. coli, Klebsiella, and Salmonella, and which is present in Y. pestis and some Y. pseudotuberculosis strains, only Y. enterocolitica biotype 1B strains can produce this factor (Carniel et al., 1996). Biotype 1B strains also contain a chromosomal pathogenicity island known as the Yersinia secretion apparatus (Ysa) encoding a second type III secretion system (TTSS) which encodes at least 11 Yersinia secreted proteins (Ysps). Their role in pathogenesis is not completely understood; however, they are thought to be important virulence determinants (Iwobi et al., 2003; Venecia and Young, 2005).
Human Infection with Y. enterocolitica
The disease caused by Y. enterocolitica and Y. pseudotuberculosis is termed “yersiniosis.” Y. enterocolitica is the most commonly reported species causing human yersiniosis (Bucher et al., 2008; ECDC, 2010; Long et al., 2010). Porcine animals are the major reservoir for human pathogenic strains (Tauxe et al., 1987; Fredriksson-Ahomaa et al., 2006). Most non-porcine isolates, reported in countries with a high incidence, lack the necessary virulent determinants for pathogenesis (Bucher et al., 2008). Bioserotype 4/O:3 is frequently isolated from pigs and is therefore an important source of human infection (Thisted Lambertz and Danielsson-Tham, 2005).
The highest incidence of infection is reported in children under the age of 4 who typically present with self-limiting diarrhea (Prentice et al., 1991; Stolk-Engelaar and Hoogkamp, 1996; Abdel-Haq et al., 2000), which may be bloody. Older children and adults are more likely to present with fever and abdominal pain, along with diarrhea and/or vomiting (Fenwick and McCarthy, 1995; Bottone, 1997; Verhaegen et al., 1998; Zheng et al., 2007). Depending on the age and health status of the host, the clinical manifestation of gastrointestinal disease can vary (Bottone, 1999; Howard et al., 2006). Right iliac fossa pain due to inflammation of the terminal ileum and cecum, with mesenteric lymphadenitis (Matsumoto et al., 1990; Matsumoto et al., 1991), (mimicking appendicitis) and septicemia may occur (Prentice et al., 1991; Stolk-Engelaar and Hoogkamp, 1996; Falcão et al., 2004), along with infections of other extraintestinal sites such as wounds, joints, and the urinary tract. The infectious dose is unknown but is likely to exceed 104 CFU (Robins-Browne, 2007); the onset of illness is usually 24–48 h after ingestion. Some pathogenic Yersinia species then pass through the stomach, multiplying in the small bowel and colonizing the epithelial cell surface of the intestinal mucosa (Hanski et al., 1989; Grutzkau et al., 1990). The organisms can also invade by binding to intestinal brush-border membranes, from where they penetrate M cells and gain access to multiply in Peyer's patches (Hanski et al., 1989; Grutzkau et al., 1990; Bottone, 1997). Bacteria taken up by M cells are usually phagocytosed and killed by macrophages within the Peyer's patches. However, pathogenic Y. enterocolitica factors (including type III secretion effectors and, in some cases, yersiniabactin) enable them to resist phagocytosis. YadA and Ail help these bacteria to escape complement-mediated cell death (Hanski et al., 1989; Grutzkau et al., 1990).
Yersiniosis is diagnosed following isolation of the bacterium from human feces or blood, or on occasion, following the erroneous removal of a healthy appendix. Positive isolations can be further confirmed by biochemical and serological characterization (FDA, 2005). Bacteriological culture for Yersinia species from patients presenting with gastrointestinal symptoms is not a routine protocol used in many hospital laboratories (Sharma et al., 2008), a feature that may lead to underestimates of yersiniosis.
Along with appendicitis, yersiniosis has been misdiagnosed as Crohn's disease (Barry and Lobba, 1977). Post-infectious sequelae include reactive arthritis (Ternhag et al., 2008) particularly in individuals of tissue type HLA-B27 and erythema nodosum. Y. enterocolitica septicemia is more frequent in patients with iron overload (Boelaert et al., 1987; Prentice et al., 1991) and may on occasion be associated with blood transfusions (Prentice, 1992; Kuehnert et al., 2001; Leclercq et al., 2005). This occurs when viable bacteria multiply in the donated unit of blood during refrigerated storage.
Incidence Rates
Y. enterocolitica is a globally distributed gastrointestinal pathogen (Thomson et al., 2006) and is isolated from humans in many countries throughout the world. The bacterium is found in areas of the world with cooler climates such as Canada, the Western Coast of South America, Europe, Australia, New Zealand, and South Africa (Fenwick et al., 1996; Falcão et al., 2004; Nesbakken, 2005).
Yersiniosis is a reportable disease in humans in several countries, including Norway and Denmark, where it is the third most commonly reported cause of acute enteritis after campylobacteriosis and salmonellosis. In the past 10 years, 80–150 cases were recorded annually in Norway (Grahek-Ogden et al., 2007). In 2009, 238 cases were confirmed in Denmark (EFSA, 2011). It was once thought that there was a seasonal variation, as most cases were reported during the cooler autumn and winter months (Verhaegan et al., 1998). However, in 2010, a report by the European Centre for Disease Prevention and Control (ECDC) offered no clear seasonal pattern for yersiniosis infection over a 3-year period (ECDC, 2010).
The number of reported cases of yersiniosis is particularly low in England and Wales. There were 0.1 cases per 100,000 individuals reported in the United Kingdom in 2005, in comparison to 12.2 in Finland and 6.8 in Germany in the same year (Ortiz Martínez et al., 2010). In 2009, Ireland and Italy had the lowest number of confirmed cases with 0.07 and 0.02 confirmed cases per 100,000 individuals, respectively (EFSA, 2011).
The first reported case of the North American strain of Y. enterocolitica, bioserotype 1B/O:8 was identified in Germany in 2003. The strain was isolated from a 4-year-old boy admitted to hospital with the characteristic symptoms of abdominal pain, fever, and diarrhea. Other symptoms that this child presented with included leukocytosis and anemia (Schubert et al., 2003). The incidence of 1B/O:8 has declined in the United States and is rarely reported in Europe; however, an increase has been seen in Poland since 2006 (Rastawicki et al., 2009).
Outbreaks of Yersiniosis
Outbreaks attributed to Y. enterocolitica are rare in Europe and elsewhere. A report by Grahek-Ogden et al. (2007) highlighted that, in one outbreak, 11 individuals were infected with Y. enterocolitica O:9 from a ready-to-eat pork product known as brawn, a traditional food consumed at Christmas. One patient developed reactive arthritis, four patients were hospitalized, and two patients died. In these latter cases, both were elderly individuals with underlying medical conditions. The brawn was prepared by layering precooked head muscles, veal, lard, and spices in a mould and cooking to 74°C. Due to the high fat content, lard can enhance bacterial survival (Grahek-Ogden et al., 2007). Pork products are well-established sources for Y. enterocolitica infection in other European countries, with a high incidence of yersiniosis being reported in Belgium (Tauxe et al., 1987), Germany (Bucher et al., 2008; Rosner et al., 2010), and Sweden (Boqvist et al., 2009).
Y. enterocolitica is rarely associated with outbreaks in the United States. Between 1996 and 2007, 1,903 foodborne cases were reported through the FoodNet surveillance program from seven U.S. states. This represents a crude incidence rate of 0.55 cases/100,000 persons per year. The majority, 1,355 (71%), were attributed to Y. enterocolitica (Long et al., 2010). Some specific foods have been linked with Yersinia-based outbreaks in the United States. These include chocolate milk (Black et al., 1978), powdered milk (Shayegani et al., 1983), pasteurized milk (Tacket et al., 1984), and various foods, including tofu, contaminated with spring water (Tacket et al., 1985).
In 1990 in Georgia, a yersiniosis outbreak affecting 14 infants that occurred between December 1988 and January 1989 associated with household preparation of chitterlings (pork intestines) (Lee et al., 1990) was reported. Similar outbreaks occurred that affected 12 infants in Tennessee between 2001 and 2002 (Jones et al., 2003) and nine infants in Chicago between 2002 and 2003 (CDC, 2003). African-American infants younger than 1 year of age who were exposed to the preparation of chitterlings were most at risk of infection. Although they did not consume the chitterlings, they were exposed during their preparation, which involved substantial handling of large amounts of potentially contaminated product by household food handlers, which led to infections (Jones et al., 2003).
Occurrence of Y. enterocolitica in Pork and Other Foods
Contaminated food or drink is the most common vehicle of infection for Y. enterocolitica, which has been cultured from meats (including pork, beef, and lamb), as well as oysters, fish, raw milk, and cheese (Rea et al., 1992; Davies et al., 2001; Fredriksson-Ahomaa et al., 2004).
In Austria, Hilbert et al. (2003) carried out a study to detect Y. enterocolitica isolates cultured from a variety of foodstuffs by means of biochemical testing. These data showed that, of 120 different foods sampled, (including 45 raw pork samples, 30 chicken samples, 30 beef, and 15 fish samples), 90 isolates were determined to contain urease-positive Y. enterocolitica following biochemical testing. Several studies have been undertaken to determine whether fish were also a source of Yersinia infection. One such study analyzed samples from fresh fish outlets in France, Great Britain, Greece, and Portugal. Only fish studied in Great Britain yielded positive samples, with seven of 30 (23%) isolates found. Four of the isolates were derived from salmon and three from trout. This study did not determine the pathogenicity of the isolates. (Davies et al., 2001).
Biotype 1A Y. enterocolitica (or other virulence plasmid negative species such as Y. frederikensii) have been cultured from raw milk in many countries, including Ireland (Rea et al., 1992), Iran (Soltan-Dallal et al., 2004), the United States (Jayarao et al., 2001), the United Kingdom (Greenwood et al., 1990), and France (Desmasure et al., 1997). Typically, and as described in a report from Turkey that sampled raw milk and cheese for Yersinia in 2002 and 2003, a high percentage (69 of 200 [35%]) were positive. Fifty-five of the Yersinia isolates recovered were found in 100 raw milk samples tested, and 25 (47.3%) were identified as Y. enterocolitica; 14 isolates were cultured from 100 cheese samples, and five (35.7%) were determined to be Y. enterocolitica. None of the isolates were virulent, and all were biotype 1A. Contamination was thought to have occurred during human handling, and was contributed to by poor environmental conditions, low water quality, and the use of previously contaminated raw milk to produce the cheese (Yucel and Ulusoy, 2006). In a similar study on raw and pasteurized milk in Northern Iran, five of 310 (1.6%) raw milk samples contained Y. enterocolitica, whereas none of the 40 pasteurized milk samples were found to be positive. Bioserotyping studies performed on the five samples determined three isolates to be biotype 1A and two isolates to be bioserotype 1B/O:9 (Soltan-Dallal et al., 2004). In Poole, United Kingdom, in 1990, strains of biotype 1A Y. enterocolitica were isolated from pasteurized milk from a local dairy. Similar organisms were found in unpasteurized milk from several farms supplying the dairy. The cause of contamination of the pasteurized milk was faulty processing rather than heat resistance of the strains (Greenwood et al., 1990). Occasional milk contamination with pYV-positive strains of Y. enterocolitica has also been reported in association with some outbreaks.
Fredriksson-Ahomaa et al. (2004) reported on Y. enterocolitica cultured from eight butcher's shops in the Munich area. Of these, six were found to have pathogenic Y. enterocolitica bioserotype 4/O:3. This biotype is frequently isolated from humans in Germany and has also been reported in slaughter pigs in Southern Germany (Fredriksson-Ahomaa et al., 2004). A study in Bavaria compared detection rates between a standard culture method and a molecular method. Of 558 samples (comprising 446 pork samples and 51 game meat samples), 81 (18%) and three (6%), respectively, were confirmed to contain the Y. enterocolitica ail gene using real-time polymerase chain reaction (PCR), whereas the culture method employed identified Y. enterocolitica O:3 in 46 (10%) of the pork samples and none in the game meat samples. None of the 61 raw milk samples yielded any positive results from either method (Messelhäusser et al., 2011).
In Germany, pig carcasses and offal are widely transported to small butcher shops where meat is further processed for consumers. Contaminated pig meat from slaughterhouses may transmit pathogenic bacteria to these shops (Fredriksson-Ahomaa et al., 2004).
Prevalence of Y. enterocolitica in Pigs
Healthy pigs are often carriers of Y. enterocolitica that are pathogenic to humans (Nesbakken et al., 2003). Fattening pigs are frequently colonized with pathogenic Y. enterocolitica in the tonsils, but they may also excrete this pathogen in their feces. However, culturing of the tonsils can provide a more reliable estimate of occurrence compared to screening of the feces (Fredriksson-Ahomaa et al., 2009).
A 2-year study on the prevalence of enteropathogenic Yersinia in England found 278 of 630 (44%) pigs positive for Y. enterocolitica. The study reported that, of 45 farms tested, 31 (69%) were positive for this pathogen. The most common bioserotypes were determined to be 2/O:9 (33%) and 2/O:5 (26%), respectively (Ortiz Martínez et al., 2010).
Fredriksson-Ahomaa et al. (2007) carried out a study to determine the prevalence of pathogenic Y. enterocolitica in a slaughterhouse in Switzerland. They found 148 of 212 (70%) pig tonsils positive with pathogenic Y. enterocolitica using real-time PCR, the most common bioserotype determined to be 4/O:3 (Fredriksson-Ahomaa et al., 2007). A similar study on isolates from Germany and Finland also found 4/O:3 to be the most common bioserotype recovered from the tonsils of finishing pigs (Fredriksson-Ahomaa et al., 2003).
In Finland, a study to detect Y. enterocolitica in fattening pigs at slaughter originating from 46 farms in Belgium, Italy, and Spain found 100% prevalence on farms in Italy and Spain and 80% prevalence on farms in Belgium, with 4/O:3 again identified as the predominant bioserotype. Tonsils from 829 pigs were examined, and 411 (50%) were identified as being positive for Y. enterocolitica (Martínez et al., 2011). Bonardi et al. (2003) investigated the cecal material, carcass swabs, and tonsils of 150 pigs in two slaughterhouses in Northern Italy. Pathogenic Y. enterocolitica were identified in six (4%) of the cecal material samples analyzed, 22 (14.7%) of the tonsils, and none of the carcass swabs. Bioserotype 4/O:3 was the predominant strain found in the samples (Bonardi et al., 2003).
In Belgium, a study analyzing pig tonsils from 139 slaughtered pigs over a 4-month period in 2007 and 2008 recovered 37.4% (52/139) pathogenic Y. enterocolitica. These samples were taken from two slaughterhouses, all of which were identified as bioserotype 4/O:3 (Van Damme et al., 2010). In order to validate a method to detect virulent Y. enterocolitica, Nowak et al. (2006) tested 180 samples from 60 slaughter pigs in Germany. They found 20 (33.3%) to be positive. After validating their method, they applied it to testing samples from pigs from conventional and alternative housing systems. Some 29% (60/210) and 18% (36/200) of the animals were positive from conventional and alternative housing, respectively. The anatomical site from which they recovered most of their positive samples was reported to be the tonsils (Nowak et al., 2006).
In most European Union (EU)–member states, as in the United States, it is not mandatory to report Y. enterocolitica in animals. Thus, zoonotic epidemiological data may be incomplete (EFSA, 2011).
In order to determine the prevalence of pathogenic Y. enterocolitica in pigs in the United States, Bhaduri et al. (2005) conducted a study sampling 2,793 fecal samples in pigs from 77 production sites in the 15 eastern and mid-western pork-producing states over a 27-week period in 2000 and 2001. The mean prevalence was 13.1% (366 of 2,793 fecal samples tested) when both PCR- and culture-positive results were combined. These authors concluded that pigs in the United States are a reservoir for Y. enterocolitica strains potentially pathogenic for humans (Bhaduri et al., 2005).
Antimicrobial Resistance
Dissemination of antimicrobial resistance genes among bacterial strains has become an increasing problem in infectious diseases. This in turn increases the likelihood that human bacterial pathogens that have food animal reservoirs will develop cross-resistance to drugs approved for use in clinical medicine, a feature that has important clinical implications and which may impact on human health (Angulo et al., 2004). In Europe, data on antimicrobial resistance of Yersinia species is not routinely recorded by EFSA, but information from individual studies is available (EFSA, 2010).
Baumgartner et al. (2007) reported that of 386 Y. enterocolitica from human patients, pigs, and retail pork samples studied, only six multidrug-resistant isolates were detected. A more recent study of 379 chicken and beef samples identified 60 (16%) Yersinia species. Five (8%) were multidrug resistant, and they were confirmed to be Y. enterocolitica (Soltan Dallal et al., 2010). In 2004, Mayrhofer et al. (2004) analyzed 304 meat samples (pork, beef, chicken, turkey, and minced meat) collected in butchers and supermarkets in Austria. While both pathogenic and non-pathogenic Y. enterocolitica isolates were found in the meat, they were all determined to be susceptible to the panel of antimicrobial compounds tested (Mayrhofer et al., 2004).
An investigation by researchers in Barcelona, to study antimicrobial resistance trends in enteropathogenic bacteria isolated from fecal and intestinal samples of patients, analyzed 75 Y. enterocolitica strains in the late 1980s and a further 20 strains isolated in the late 1990s (Prats et al., 2000). All were bioserotype 4/O:3. Data published showed that isolates from both time points were resistant to ampicillin and cephalothin, a feature that is to be expected as Y. enterocolitica has a natural resistance to both ampicillin and first generation cephalosporins. However, the resistance rate increased to 90% for streptomycin and sulphonamides, 70% for trimethoprim/sulphamethoxazole, 60% for chloramphenicol, and 5% for nalidixic acid, antimicrobial agents to which these bacteria were previously susceptible (Prats et al., 2000).
In Poland, Rastawicki et al. (2000), using a standardized disk diffusion assay, determined the susceptibility of 114 clinical strains Y. enterocolitica to a panel of antimicrobial compounds. All isolates were bioserotype 4/O:3 and were resistant to ampicillin and cephazolin, but they were susceptible to all of the other drugs tested. These authors reported that the most effective antimicrobial agents included the fluoroquinolone ciprofloxacin, as well as third-generation cephalosporins such as cephotaxime and cephtriaxone (Rastawicki et al., 2000). Food is but one route by which antimicrobial-resistant bacteria can be transmited to humans, and consequently the development and spread of antimicrobial resistance through food must be prevented (EFSA, 2008).
Remedial Strategies, Prevention, and Control
There is as yet no vaccine available for Y. enterocolitica (De Berardis et al., 2004). Therefore, prevention measures are required in order to avoid contamination of foodstuffs, which may lead to subsequent human infection. Considering the reservoir of pathogenic Y. enterocolitica, avoidance of the consumption of contaminated raw meat (especially pork), unpasteurized milk, and unchlorinated water is important; in addition, routine hand-washing for persons exposed to contagious material and proper drainage systems for human and animal fecal material are important.
A number of biosecurity measures can be implemented at the primary stage of production so as to minimize the risk of contamination/infection (Fig. 1). Such measures include providing footbaths containing an appropriate disinfectant for use upon entering the farm or slaughterhouse environment, along with the provision of a change of footwear. Ensuring that any areas where pigs may be housed are free from rodents and birds by netting of vents, and keeping the surroundings of the farm or slaughterhouse free from debris are crucial. Allowing only essential vehicles and visitors such as animal feed providers to access the site will help to reduce the risk. Finally, use of equipment that is properly cleaned and disinfected (FAO, 2010) should be encouraged.
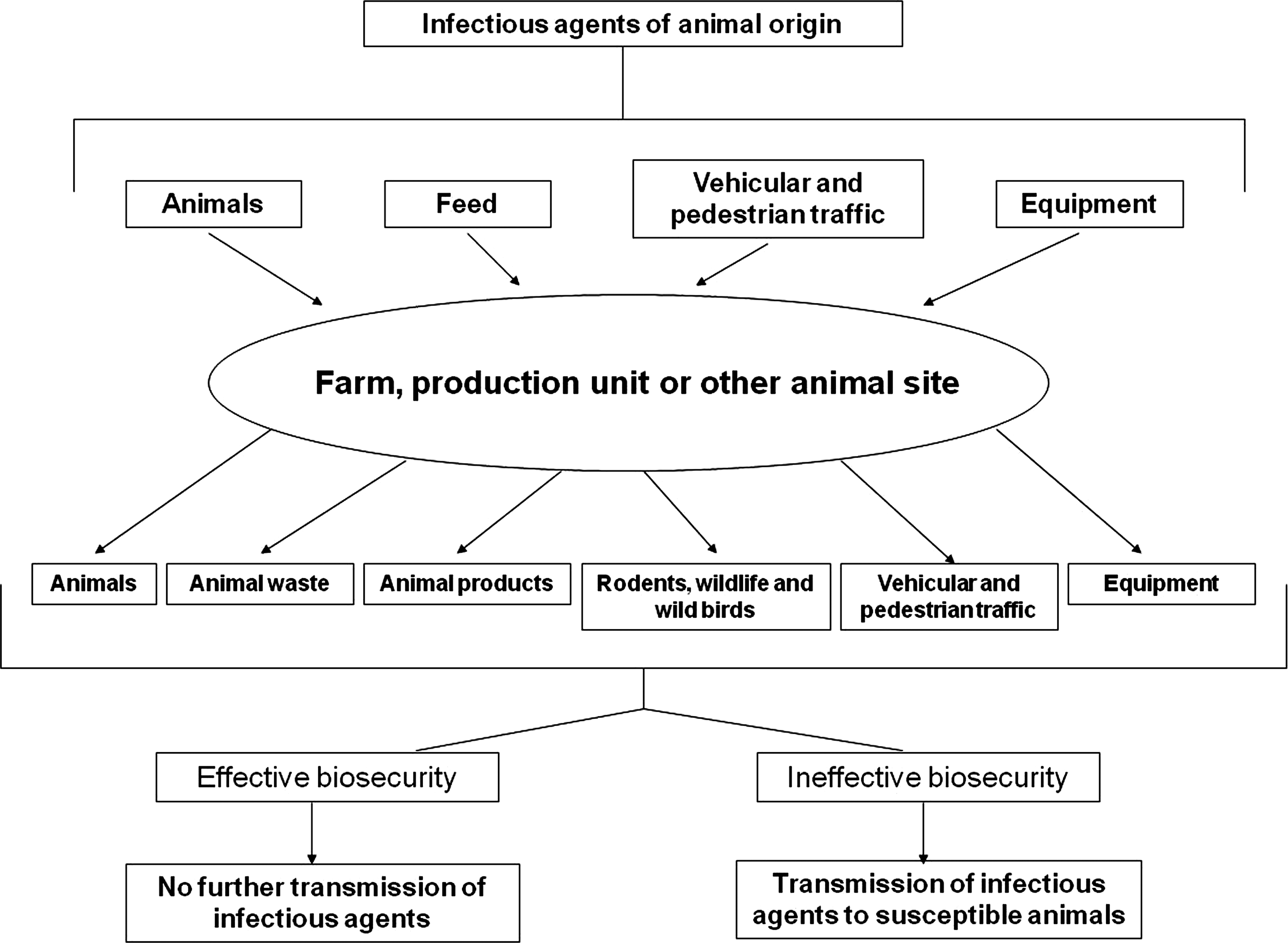
A schematic illustration showing the key steps in a biosecurity program aimed at reducing the transmission of a biological hazard.
Skerjve et al. (1998) outlined steps that can be taken at the farm and slaughterhouse level so as to reduce or eliminate herd carriage with Y. enterocolitica. These measures include farrow-to-finish production, where there is no contact between herds, and an under-pressure ventilation system to allow constant aeration of the housing units. The use of hygiene barriers for humans on herd entry would help lower the prevalence rate. Clean straw bedding should be used, as it was found that bedding can favor Y. enterocolitica survival (Skjerve et al., 1998). In a surveillance study to determine how different conditions can affect the prevalence of Y. enterocolitica in pigs at farm level, researchers found that the use of an organic production type causes less stress on the animal and thus reduces the prevalence rate. Other factors that can reduce prevalence include a limited use of antimicrobial agents, having a lower animal density, the use of a municipal water supply, and the limitation of snout contact between animals (which can cause fecal shedding) through the use of solid walls separating the pens (Virtanen et al., 2011).
Nesbakken et al. (2007) reported a strategy for producing Y. enterocolitica–-free pigs using a specific pathogen-free (SPF) breeding pyramid. Most Norwegian pig production is organized in a closed breeding system in which primary nucleus-herd farms sell breeding animals to secondary multiplying-herd farms. These multiplying-herd farms sell breeding animals to conventional-herd farms. In turn, animals from young pig-production farms are sold to fattening-herd farms. By founding a breeding pyramid with animals obtained by hysterectomy, herds free from well-defined animal diseases can be obtained. Through these measures, Y. enterocolitica has been eliminated from test herds, suggesting that the impact of these pathogens on human health can be reduced (Nesbakken et al., 2007).
At the slaughterhouse level, serological testing can also be used to control Y. enterocolitica. Ideally, only herds that are free of the organism should be in contact during transport and when in the lairage, and they should be slaughtered at the same time. Herds that are carriers of Y. enterocolitica should be kept separate from non-carriers and slaughtered separately (Nesbakken et al., 2003). An enzyme-linked immunoassay (ELISA) was applied to test for three serotypes of Y. enterocolitica in pigs at the farm and slaughterhouse level (Thibodeau et al., 2001). These authors found no cross-reaction against a large number of Enterobacteriaceae. Although the ELISA has a low level of detection in individuals, pooling of samples at farm level can be applied to detect infected herds.
The primary contamination source in pig slaughterhouses seems to be Yersinia-positive tonsils, which can subsequently contaminate the carcass, offal, and slaughterhouse environment during the slaughtering process (Nesbakken, 1988; Kapperud, 1991). Possible points of contamination during the slaughtering process include evisceration, removal of the pluck sets (heart, lungs, esophagus, trachea, diaphragm, liver, kidneys, tongue, and tonsils), and meat inspection (Nesbakken et al., 1994; Fredriksson-Ahomaa et al., 2000; Laukkanen et al., 2010b). Implementing strict hygiene procedures and educating workers in the slaughterhouse may lower the incidence of cross-contamination. Studies have also found that enclosing the rectum bag during slaughter reduces carcass contamination in the abattoir (Nesbakken et al., 1994; Laukkanen et al., 2010a). Nesbakken et al. (2008) determined that blast chilling had no effect on reducing pathogenic Y. enterocolitica when present on pig carcasses during slaughter.
Conclusion
Y. enterocolitica is a zoonotic pathogen found in a range of hosts and environments. Several of the bioserotypes are known to cause illness and disease in humans. Pathogenic Y. enterocolitica possess virulence factors, including inv and ail chromosomal genes and plasmid-mediated markers, which allow this organism to survive and proliferate within the human host. Outbreaks of yersiniosis are rare, with reported cases being sporadic. Scandinavian countries report the highest number of confirmed cases annually.
Pigs are the primary source of pathogenic strains such as bioserotype 4/O:3. These bacteria colonize the tonsils of healthy animals. There is a growing body of information describing the emergence of multidrug-resistant strains. Thus, implementation of scientifically based control measures at all levels of production and retail are required to reduce the risk of contamination and decrease the impact on public health.
Footnotes
Acknowledgments
We wish to thank the Irish government's Department of Agriculture, Fisheries and Food (DAFF) for their financial contribution to this project under the Food Institute Research Measure (FIRM) initiative (grant 06/R&D/D419).
Disclosure Statement
No competing financial interests exist.