
Abstract
Listeria monocytogenes is a foodborne pathogen causative of opportunistic infections. Listeriosis is associated with severe infections in pregnant women causing abortion or neonatal listeriosis. An alternative to antibiotics are safe novel bacteriocins peptides such as enterocin CRL35 with strong antilisterial activity produced by Enterococcus mundtii CRL35. In the present paper, our goal is to study the effectiveness of this peptide and the producer strain in a murine model of pregnancy-associated listeriosis. A single dose of 5×109 colony-forming unit of L. monocytogenes FBUNT (Faculty of Biochemistry—University of Tucumán) resulted in translocation of pathogen to liver and spleen of BALB/c pregnant mice. The maximum level of Listeria was observed on day 3 postinfection. Interestingly, the intragastric administration of enterocin CRL35 significantly reduced the translocation of the pathogen to vital organs. On the other hand, the preadministration of E. mundtii CRL35 slightly inhibited this translocation. Listeria infection caused a significant increase in polymorphonuclear leukocytes at day 3 postinfection compared to the noninfected group. This value was reduced after the administration of enterocin CRL35. No significant changes were observed in either white blood cells or lymphocytes counts. Based on the data presented in the present work enterocin CRL35 would be a promising alternative for the prevention of Listeria infections.
Introduction
L
After the ingestion of contaminated food, Listeria cause gastroenteritis in humans and can traverse the intestinal barrier to cause systemic disease. The groups that are susceptible to severe infections are pregnant women, the elderly, and patients with immunocompromising conditions (Schlech, 2000; Chen et al., 2007). Women infected with the pathogen during pregnancy may develop life threatening infections such as septicemia, meningitis, encephalitis, etc. Furthermore, infection of the fetus can result in abortion or neonatal listeriosis (Chen et al., 2007; Van de Velde et al., 2009; Orsi et al., 2011). In humans, approximately 33% of clinically documented cases of listeriosis are among pregnant women (Bakardjiev et al., 2004; Orndorff et al., 2006).
Long treatments with antibiotics are required for Listeria infections that in turn results in the emergence of multidrug-resistant Listeria strains (Charpentier and Courvalin, 1999). High doses of amoxicillin in combination with an aminoglycoside (Rihakova et al., 2010) or ampicillin, either alone or in combination with gentamicin, penicillin G (Van de Velde et al., 2009), or trimethoprim-sulfamethoxazole are mainly used. On the other hand, the discovery of conventional antibiotics is slow and this generates the need for alternative strategies to treat or prevent listeriosis (Charpentier and Courvalin, 1999; Palumbo et al., 2010). Novel and safe molecules such as bacteriocins are now a major interest to prevent the emergence of resistant strains (Rihakova et al., 2010). Bacteriocins are antimicrobial peptides produced by bacteria and are used as “natural” preservatives by food industry as a strategy to prevent the survival and growth of foodborne pathogens (Galvez et al., 2007; Settanni and Corsetti, 2008). In this regard, subclass IIa bacteriocins are the most promising agents for an effective prevention or treatment of listeriosis due to their strong antilisterial activity (Drider et al., 2006).
Enterocin CRL35 is a subclass IIa bacteriocin produced by Enterococcus mundtii CRL35 that is active against Listeria. It was biochemically and genetically characterized (Saavedra et al., 2004b; Salvucci et al., 2010). In addition, previous studies demonstrated that combinations of enterocin CRL35 with certain antibiotics strongly increase their potency leading to a marked reduction of their dose (Minahk et al., 2004; Salvucci et al., 2010). Considering the promising use of enterocin CRL35, for the prevention or treatment of listeriosis, the aim of the present work was to evaluate the effectiveness of this bacteriocin or its producing bacteria (E. mundtii CRL35) in a murine model of pregnancy associated L. monocytogenes infection.
Materials and Methods
Strains and growth conditions
E. mundtii CRL35 (Centro de Referencia para Lactobacilos (CERELA) Culture Collection) and L. monocytogenes FBUNT (serotype 4b) (Vignolo et al., 2000) were grown for 16 h at 37°C in LAPTg (1% yeast extract, 1.5% peptone, 1% tryptone, 0.1% Tween 80, 1% glucose) and brain heart infusion (BHI) broth, respectively.
For the administration of Listeria in animals, L. monocytogenes FBUNT was grown in BHI medium at 37°C for 16 h, cells were collected by centrifugation and washed with 0.01 M phosphate-buffered saline (PBS) (pH 7.3). The pellet was resuspended in the same buffer to obtain a final cell density of 5×109 colony-forming units (CFU)/0.2 mL. E. mundtii CRL35 was grown at 37°C for 16 h, cells were collected by centrifugation, washed, and resuspended to a final cell density of 2×109 CFU/0.2 mL in 0.01 M PBS (pH 7.3).
Doses of 0.2 mL of L. monocytogenes FBUNT and/or E. mundtii CRL35, prepared as mentioned before, were orally administrated to each pregnant mouse by using a stainless steel feeding needle and a 1 mL syringe following the designed protocol.
Bacteriocin purification
Enterocin CRL35 was purified from a 16 h culture supernatant by our standard laboratory procedure of ammonium precipitation and passage through a C-18 solid phase extraction cartridge (Saavedra et al., 2004a, Vera Pingitore et al., 2007). The active fraction was collected, concentrated, and lyophilized obtaining the enterocin extract (Ent CRL35). Ent CRL35 was rehydrated in 0.01 M PBS (pH 7.3) and doses of 65 μg corresponding to 256 AU were intragastrically administrated to each animal according to the experimental protocol.
Animals
Pregnant female BALB/c mice at gestational day 14 (37–39 g) were obtained from the inbred closed colony maintained at CERELA. They were provided a temperature-controlled environment (20°C±2°C) with a 12 h light/dark cycle. Mice were fed with a balanced rodent diet and fresh water ad libitum. All animal protocols were approved by the Animal Protection Committee of CERELA and handled in conformity with the latest recommendations of the Federation of European Laboratory Animal Science Associations. All experiments comply with the current laws of Argentina.
Experimental design
A murine model of pregnancy associated L. monocytogenes infection was used (Abram et al., 2003). On gestational day 14, mice were randomly divided into groups (at least three animals per group in each experiment were used) and L. monocytogenes and/or enterocin CRL 35 was orally administrated (therapeutic treatment) according to the following scheme: Group L: L. monocytogenes FBUNT (5×109 CFU in 200 μL PBS); Group L+Ent CRL35: L. monocytogenes FBUNT (5×109 CFU in 200 μL PBS), 6 h later and every 12 h for three days: 65 μg in 200 μL PBS of Ent CRL35 per animal; Group Ent CRL35 65 μg in 200 μL PBS of Ent CRL35 every 12 h for three days. Control mice received only PBS (200 μL).
Enterocin CRL35 has shown a minimal inhibition concentration of 1.6 ng/mL against L. monocytogenes FBUNT (Salvucci et al., 2010). The dose used in the present study (65 μg) was selected to ensure that enough concentration of active molecules could reach to the intestinal tract of treated animals.
To evaluate the efficacy of the natural bacteriocin producing strain, a preventive administration scheme was designed as follows, Group E5h+L: E. mundtii CRL35 (2×109 CFU in 200 μL PBS) and 5 h later animals were challenged with L. monocytogenes FBUNT (5×109 CFU in 200 μL PBS); Group E3d+L: E. mundtii CRL35 (2×109 CFU in 200 μL PBS) for three consecutive days and then animals were challenged with L. monocytogenes FBUNT (5×109 CFU in 200 μL PBS); Group E3d: E. mundtii CRL35 (2×109 CFU in 200 μL PBS) for three consecutive days; Group L: L. monocytogenes FBUNT (5×109 CFU in 200 μL PBS). Control mice received only PBS (200 μL).
Mice were surveyed for 72 h post challenge to observe signs of diarrhea, respiratory distress, and depression. The translocation of L. monocytogenes FBUNT to liver and spleen was monitored on day 1, 3, and 7 postinfection. Mice were anesthetized with ketamine (100 mg/kg of animal) and xylazine (5 mg/kg of animal) administered via intraperitoneal and humanely euthanized. Liver, spleen, and fetus were aseptically removed, weighted, homogenized, and plated directly on Listeria selective agar (Oxoid) and BHI. Plates were incubated at 37°C for 48 h. Viable counts were expressed CFU per gram of organ.
Blood collection and determination of total and differential number of leukocytes
Blood was collected by cardiac puncture from anesthetized mice. A 21-gauge needle and 1 mL syringe were used to obtain blood from the left ventricle. The blood was then immediately transferred to (1) EDTA (1.5 mg/mL) tubes for total leukocytes counting determined with a hemocytometer; (2) blood smears stained with MayGrünwald-Giemsa for differential cell counts; (3) tubes containing hemoculture media (Hemoculture media–Britania Laboratories). Bacteremia was reported as negative or positive hemocultures after incubation for 24 h at 37°C. Positive control culture was obtained by L. monocytogenes FBUNT grown in the same media.
Feed and fluid intake and body weight measurements
Food and water consumption was measured in each group along the days of the experiments. Feed intake was determined by subtracting the amount of left uneaten food from the total amount provided to the animals. Animal body weight was monitored daily and expressed as the median increment of weight for each group.
Statistical analysis
Microbiological data were expressed as mean log CFU±standard error of the mean. Statistical analysis was performed with the software package Minitab14 software (Minitab, Inc.) using one-way analysis of variance followed by a Tukey's post hoc test, and p≤0.05 was considered significant.
Results
Enterocin CRL35 inhibits the multiplication of L. monocytogenes FBUNT in pregnant mice
The single intragastric dose of 5×109 CFU of L. monocytogenes FBUNT allowed the translocation of the intestinal barrier and the localization of the pathogen in liver and spleen. The administration of one dose of 1×108 or 1×107 CFU did not result in the translocation of the pathogen to the evaluated organs (data not shown).
Microbiological counts of the pathogen expressed as CFU per gram of infected animal organs are shown in Figure 1. The maximum level of L. monocytogenes FBUNT was observed on day 3 postinfection with count of 2.6×104 CFU/g in the spleen and 3.9×103 CFU/g in the liver (Group L). The administration of Ent CRL35 every 12 h postinfection decreased the counts of the pathogen to 6×102 CFU/g in the spleen and to 7×102 CFU/g in the liver (Fig. 1B). These results represent a significant decrease of 1.6 log and 1 log of L. monocytogenes FBUNT in spleen and liver, respectively (p=0.008). On day 7 postinfection, L. monocytogenes FBUNT was not recovered from the evaluated organs of L and L+Ent CRL35 animal groups.

Therapeutic administration of enterocin CRL35 in Listeria infections. Liver and spleen colonization levels of Listeria monocytogenes FBUNT at one
L. monocytogenes was undetectable in the different organs of control animals (PBS) and in those receiving only the bacteriocin (Ent CRL35) on day 1, 3, or 7 postinfection (Fig. 1A, B).
No deaths or clinical signs during the complete scheme of treatment were evident in any of the animal groups (L; L+Ent CRL35; Ent CRL35 and PBS). Moreover, no bacteria were recovered from the bloodstream of mothers or fetuses evaluated at any point of time.
Feed and fluid intake assessment in pregnant mice receiving enterocin CRL35
The pattern of food and fluid intake of mice under the therapeutic treatment with enterocin CRL35 as signs of discomfort or distress was evaluated (Fig. 2).

Feed and fluid intake assessment during therapeutic administration of enterocin CRL35
Mice receiving only Ent CRL35 along three days showed a significant reduction in the feed intake (3.86 g per animal per day) compared with the control group (PBS). Although there was a decreased in food consumption among animals from the rest of the groups, no significant differences were obtained (Fig. 2A).
Regarding the water intake, L. monocytogenes FBUNT infected animals treated with Ent CRL35 (L+Ent CRL 35) and the ones receiving only Ent CRL35 during three days displayed a significant decrease in water intake (6.12 mL per animal per day) compared with the control group (PBS) (Fig. 2A). There were no significant changes of these variables on one or seven days postinfection (data not shown).
The body weight of control mice increased at a median of 1.1 g after three days post infection (Fig. 2B). L. monocytogenes FBUNT infected animals evidenced a body weight media increase of 0.95 g showing no significant differences with respect to control group.
The administration of a single dose of Ent CRL35 did not cause a decrease in the body weight. However, the continued administration of the bacteriocin during three days post infection caused a significant loss of weight in mice with a reduction of about 0.3 g (Fig. 2B). On the other hand, mice receiving L. monocytogenes FBUNT plus Ent CRL35 (L+Ent CRL35) showed an increase in body weight of 0.75 g with no significant differences with respect to control group (Fig. 2B).
Effect of the administration of enterocin CRL35 on total and differential number of leukocytes
Hematologic parameters are shown in Table 1. L. monocytogenes infection did not show an increase in total leukocytes (white blood cells [WBC]) (7.26±0.76×109 cells/L) and polymorphonuclear leukocytes (PMN) (1.38±0.14×109 cells/L) one day postinfection compared with the control group (PBS group). PMN only showed a significant increase three days postinfection in L group coincidently with the invasion of the pathogen to vital organs. On the other hand, counts for lymphocytes in pregnant mice with listeriosis (L group) were significantly higher than those observed in the control group only one day postinfection (6.04±0.1×109 cells/L).
The values are given as mean±SD.
WBC, white blood cells; PMN, polymorphonuclear leukocytes; PBS, phosphate-buffered saline.
The administration of enterocin CRL35 to L. monocytogenes FBUNT infected animals (L+Ent CRL35) showed a significant decrease in PMN on day 1 and 3 postinfection (0.94±0.2×109 cells/L and 1.4±0.4×109 cells/L, respectively) compared with the nontreated group (L group). No significant changes were observed in either WBC or lymphocytes counts.
Pregnant mice that received only the bacteriocin showed significant changes in WBC (4.46±0.5×109 cells/L) and PMN (0.54±0.2×109 cells/L) on day 1 postadministration with lower values compared with the control group (PBS group). Lymphocytes considerably increased on day 1 or 3 postadministration (3.60±0.4×109 cells/L and 3.9±0.5×109 cells/L, respectively) compared with the control group (PBS group) but significantly lower than values found for L. monocytogenes FBUNT infected mice.
Evaluation of the bacteriocin producing strain, E. mundtii CRL35, in preventing L. monocytogenes infection
Two experimental schemes were used to evaluate the preventive administration of the bacteriocinogenic strain E. mundtii CRL35 in L. monocytogenes FBUNT infection (Fig. 3). First, E. mundtii CRL35 was administered for three consecutives days to pregnant BALB/c mice before the L. monocytogenes FBUNT challenge (E3d+L group). No changes were observed in food and fluid intake in these groups of mice except for a significant decrease in water consumption compared with control mice (Fig. 3A). In E3d+L group, microbiological counts showed that there was a slight decrease in L. monocytogenes FBUNT counts, 1.6×104 CFU/g in spleen and 2.6×103 CFU/g in liver compared with L group (Fig. 3B). Second, a single dose of E. mundtii CRL35 (2×109 CFU) was administered 5 h before the L. monocytogenes challenge (E5h+L group). This resulted more effective in decreasing the number of the pathogen in the tested organs reaching 2.5×103 CFU/g in spleen and 7.5×101 CFU/g in liver (Fig. 3B).
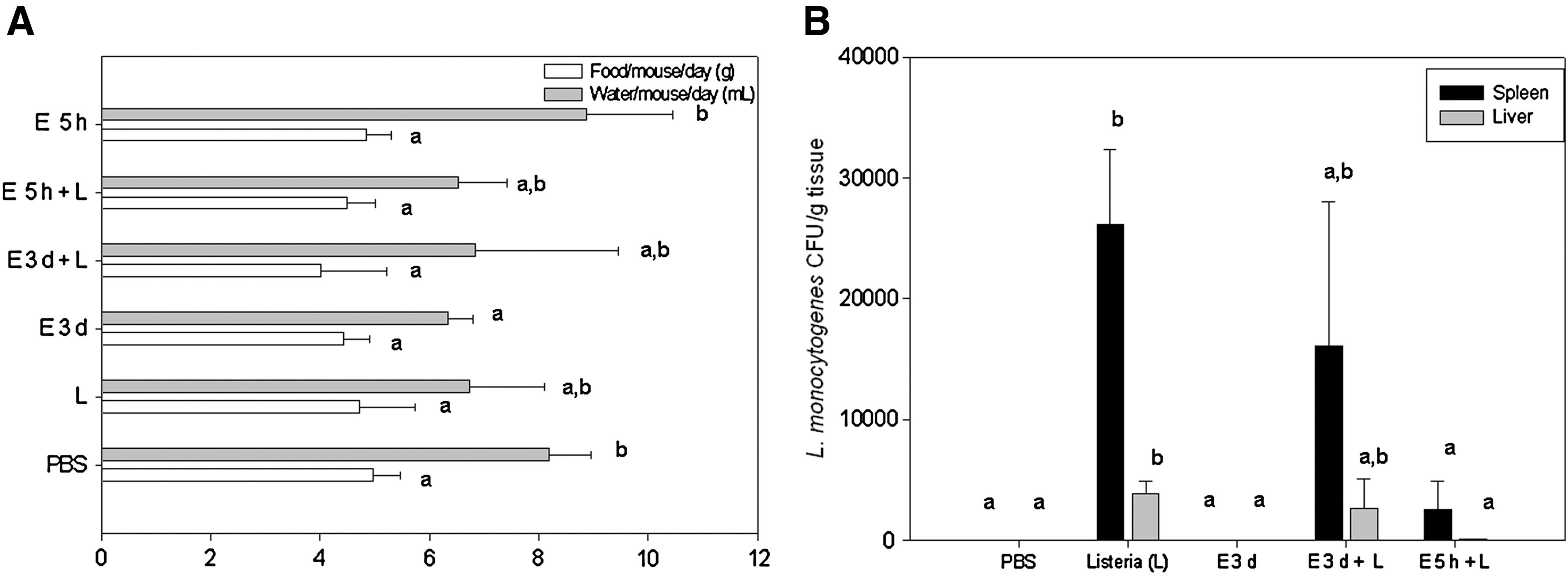
Administration of Enterococcus mundtii CRL35 in the prevention of Listeria infections.
The administration of E. mundtii CRL35 at a dose of 2×109 CFU during three consecutive days (E3d group) resulted in an increase in WBC (7.1±1.4×109 cells/L), lymphocytes (5.6±1.2×109 cells/L), and 15% of Gumpecht cells. More studies are now being carried out to fully analyze this issue.
Discussion
The effectiveness of enterocin CRL 35 and its natural producer strain was evaluated in a pregnant mice model of Listeria infection where this pathogen was administered orally. This procedure more closely imitates natural infection in humans and address behavior and translocation in the gut that are the main factors in the human infection (Kim et al., 2004).
The single intragastric dose of 5×109 CFU of L. monocytogenes FBUNT resulted in the translocation of pathogen to liver and spleen of BALB/c pregnant mice. After three days of infection, the counts of L. monocytogenes FBUNT were 2.6×104 CFU/g in the spleen and 3.9×103 CFU/g in the liver, which is in agreement with the results obtained by Czuprynski et al. (2002). L. monocytogenes FBUNT translocated to the liver and spleen but it was not found in the fetus or bloodstream during the seven days of the experiments.
No mortality of mother could be attributed to the infection at late gestational days. A recent report described that susceptibility to lethal infection occurs very early in gestation and peaked in animals inoculated at 3.5 and 6.5 gestational day when embryonic development is occurring rapidly (Suyemoto et al., 2010).
Bacteriocins are considered an alternative for prophylactic and therapeutic treatments. The in vivo effectiveness of the antilisterial enterocin CRL35 as plausible therapeutic alternative for listeriosis was evaluated.
The administration of 65 μg enterocin CRL35 provided a good protective effect against L. monocytogenes FBUNT. Lower doses (30 μg) of the bacteriocin resulted ineffective for lowering L. monocytogenes FBUNT counts in liver and spleen. Considering the proteinaceous nature of this peptide and the background of the use of bacteriocins as additive in foods, this dose may be considered safe (Foulquié Moreno et al., 2006; Garcia et al., 2010).
L+Ent CRL35 group showed L. monocytogenes FBUNT counts of 1.2 log less than the untreated mice in liver and spleen at 24 h postinfection. Interestingly, although enterocin CRL35 is sensitive to gut proteolytic enzymes in vitro (Farias et al., 1996), it retains at least part of its activity in the gut after oral administration. Dabour et al. (2009) have reported a successfully intragastrical administration of pediocin PA-1, a subclass IIa bacteriocin. The specific inhibition of Listeria by this class of bacteriocins ensures that it cannot dramatically affect intestinal microbial equilibrium (Dabour et al., 2009). This becomes another advantage against antibiotics. The mice treated with enterocin CRL35, however, have shown a slight decrease in weight along the three days of treatment but no symptoms like diarrhea have been observed. It seems possible that an improved purification of enterocin CRL35 or the use of a synthetic peptide might reduce this “collateral effect.”
E. mundtii CRL35 was evaluated in the prevention of L. monocytogenes infection. Enterococci constitute normal flora of the human intestine and they have been used as starter cultures for the preparation of several fermented dairy products all over the world (Giraffa, 2003; Foulquié Moreno et al., 2006; Khan et al., 2010). E. mundtii CRL35 lacks the genes related to virulence (Saavedra et al., 2003); it is sensitive to vancomycin and does not have hemolytic activity (Vildoza et al.; unpublished data).
The administration of 2×109 CFU of E. mundtii CRL35 5 h previous to the administration of L. monocytogenes FBUNT resulted in an effective reduction of L. monocytogenes FBUNT counts in liver and spleen. It seems possible that enterococci could inhibit or delay the translocation of Listeria, but not prevent it completely. This effect did not improve when the same dose of enterococci was given for three consecutive days prior to L. monocytogenes FBUNT challenge. Other promising strains failed in an assay of their capacity to prevent an infection. Bernbom et al. (2006) reports a higher translocation of Listeria when Lactobacillus plantarum DDEN 11007 was used in gnotobiotic rats. More recently, Dabour et al. (2009) found that Pediococcus acidilactici UL-5 increased the invasive ability of Listeria. In the latter case the authors suggest that a possible explanation for this behavior could be that the LAB decreases intestinal pH enough to induce Listeria virulence genes.
In conclusion, enterocin CRL35 has shown to be more effective in the inhibition of translocation of L. monocytogenes FBUNT than the use of the bacteriocinogenic strain and it might be considered in the prevention of Listeria infections.
Footnotes
Acknowledgments
This work was partially supported by a grant from the Agencia Nacional de Investigaciones Científicas y Técnicas (PICT06 0813). We would like to thank Dr. Laciar (University of San Luis) and Bacteriology Department from Faculty of Biochemistry, Chemistry and Pharmacy (University of Tucumán) for kindly providing L. monocytogenes FBUNT strain and Drs. V. Molina and M. Medici for their valuable contribution on animal procedures.
Disclosure Statement
No competing financial interests exist.