
Abstract
The purpose of this study was to analyze and compare antimicrobial resistance in Campylobacter spp. isolated from pork and poultry carcasses, and pork and poultry meat (at slaughterhouse level, during meat cutting, and at retail) in Belgium, using available surveillance data over the period 2004–2009. The susceptibilities of 1724 Campylobacter isolates for ampicillin, ciprofloxacin, nalidixic acid, tetracycline, erythromycin, and gentamicin were tested by E-test. Gentamicin resistance was low (near 0%) until 2007, with an increase to over 20% by 2009 for all species-matrix combinations. Resistance to tetracycline fluctuated around the same level during the entire study period and was significantly higher (p-value of <0.05) in C. coli than in C. jejuni. Erythromycin resistance was low and showed a slight decrease between 2004 and 2007, but increased from 2007 until 2009. Fluoroquinolone and ampicillin resistance was significantly higher in isolates derived from poultry, compared to pork-related isolates. This correlates with the higher use of these antimicrobials in poultry husbandry. A total of 25% of C. coli isolates from poultry showed the most apparent multiresistance (resistance to four or more antimicrobials). Approximately 1% of the poultry-derived isolates (both C. coli and C. jejuni) showed resistance to all tested antimicrobials, while none was found in pork products.
Introduction
C
Due to the zoonotic nature of campylobacteriosis, the intensive use of antimicrobial agents for therapy, prophylaxis, or growth promoters in food animal production (although the latter has been prohibited in Europe since 2006) can contribute considerably to increased resistance in clinical Campylobacter isolates (EFSA, 2009, 2010; Aerestrup and Wegener, 1999). Several European countries have monitoring programs to assess and mitigate the risk related to antimicrobial resistance (Ammon and Makela, 2010). The main mode of transmission is the consumption of undercooked meat or contamination by the food handler of fresh poultry and pork meat (Corry and Atabay, 2001; Altekruse and Tollefson, 2003; Cools et al., 2005). Since 2000, the contamination of pork and poultry carcasses and meat with Campylobacter spp. has been monitored in Belgium. The proportion of positive poultry samples shows strong annual fluctuations but overall remains high (Table 1) (Trends and Sources, 2004 –2009). Since 2004, these strains have been tested for antimicrobial resistance. Belgium does not yet have a central detailed surveillance system in place on each level of antimicrobial consumption or antimicrobial resistance, in contrast to some other European countries such as Denmark and Sweden. These countries annually publish detailed reports on the general use of antimicrobials and occurrence of antimicrobial resistance in food animals, foods, and humans. Recent efforts have been made by the European Medicines Agency (EMA, 2009) to develop a central database on the use of antimicrobial agents in animals (i.e., European Surveillance of Veterinary Antimicrobial Consumption [ESVAC]). Belgium is also working on a program for surveillance of antimicrobial use in agriculture; however, linking and explaining different observed trends in antimicrobial use and resistance development remains difficult.
Sample size of 1 g.
Sample size of 0.01 g.
Sample size of 600 cm2.
The aim of the present study is to analyze and compare antimicrobial resistance proportions in Campylobacter spp. isolated from food samples (i.e., pork and poultry carcasses and meat) from Belgium through surveillance data over the period 2004–2009. Six antimicrobials were tested, including those that are recommended for therapy. Focus was placed on detection of reduced susceptibility using European Committee on Antimicrobial Susceptibility Testing (EUCAST) epidemiological cut-off values.
Methods
Sampling and isolation of Campylobacter isolates
Samples of pork and poultry carcasses, meat, and meat products were taken by inspectors of the Belgian Federal Agency for the Safety of the Food Chain (FASFC) as part of continuous surveillance plans, as laid down in Belgian or European Legislation. Sampling was done at random at the level of production (i.e., at the slaughterhouse directly after slaughter, during meat cutting, and at retail). Samples were kept at 4°C until analysis. Food stuffs sampled were pork carcasses, broilers, spent hens, turkey carcasses, poultry parts, poultry meat preparations, and minced poultry meat.
Campylobacter isolates from food were obtained as described by ISO 10272-1 and 10272-2 (ISO, 2006). After isolation, the strains were sent to the national reference laboratory of Campylobacter at the Institute of Public Health (Brussels, Belgium).
Identification of species
DNA was extracted through heat lysis (Englen and Kelley, 2000). One colony was suspended in 100 μL of MQ water, boiled for 17 min, and chilled immediately. Three different polymerase chain reaction (PCR) protocols were utilized to confirm all isolates as belonging to the Campylobacter genus (Denis et al., 2001) and to identify the isolates as C. jejuni or C. coli by molecular weight (Debruyne et al., 2008; Denis et al., 1999; Linton et al., 1997). Primers are listed in Table 2.
Alonso et al. (2005).
Minimal inhibitory concentration (MIC) determination
Since the E-test has been found to compare favorably with agar dilution methods (Luber et al., 2003; Oncul et al., 2003), this test was chosen for its ease of use in the routine testing of single isolates. Confirmed Campylobacter isolates were subcultured on Colombia agar with 5% sheep blood (Oxoid Ltd., Hampshire, UK) for 24 h at 42°C under micro-aerophilic conditions, using sealed boxes with gas-generating packets (Biomérieux, Lyon, France). MIC were determined using the E®-test strips (Biomérieux), as described by the manufacturer. In short, inoculates were adjusted to the turbidity of a 0.5 MacFarland standard and plated on Mueller-Hinton agar supplemented with 5% sheep blood (BioRad, Hercules, CA). Metric antimicrobial strips were applied, two per plate, in opposite direction. Tested antimicrobials were ampicillin (Amp), tetracycline (Tet), nalidixic acid (Nal), erythromycin (Ery), gentamycin (Gen) (all 0.016–256 μg/mL), and ciprofloxacin (Cip; 0.002–32 μg/mL). Plates were incubated for 48 h at 37°C. C. jejuni American Type Culture Collection (ATCC) 33560 was used as control strain, and was included in each batch during E-test.
The MIC was defined as the lowest concentration of an antimicrobial agent that completely inhibited visible growth and was read at the point where the elliptical zone of inhibition intersected the MIC scale on the strip. Different measures were used to study resistance. EUCAST epidemiological cut-off values were used as the most sensitive measure of resistance development; different cut-off values are defined for C. coli (Amp >8, Tet >2, Nal >32, Ery >16, Gen >2, Cip >1) and C. jejuni (Amp >8, Tet >2, Nal >16, Ery >4, Gen >1, Cip >1). Multi-resistance was defined as simultaneous resistance to at least four antimicrobials tested.
Mutation analysis
The gyrA C257T mutation and 23S rRNA A2074C and A2075G mutations were examined by Mismatch Amplification Mutation Assay (MAMA) as described by Zirnstein et al. (1999, 2000) and Alonso et al. (2005), respectively. Primers are listed in Table 2. Resistant strains showing no mutations by MAMA were verified by sequence analysis, using Big Dye Terminator mix (Applied Biosystems, Carlsbad, CA) and analyzed on an ABI 3130 genetic analyzer.
Statistical analysis
Significance of differences between proportions of resistant isolates were evaluated using a χ2-test with Yates continuity correction for large sampling groups (N>20), whereas the two-tailed Fisher exact test was used for small sampling groups (N<20). The 5% significance level was used (p-value<0.05) for all statistical analyses.
Results and Discussion
A 6-year surveillance of randomly sampled Belgian pork and poultry meat products at the level of slaughterhouse, processing, and retail resulted in 1724 Campylobacter isolates. This isolate collection encompassed C. jejuni (n=1096) and C. coli (n=377) from poultry carcasses and meat, and C. coli (n=251) from pork carcasses. Trends per species, per food matrix, per antimicrobial, and per year were analyzed. It should be mentioned that it is difficult to compare results with data from other countries due to the unavailability of comparable data. With the exception of Denmark, other European countries only started reporting Campylobacter antimicrobial resistance data from meat since 2006–2007 (EFSA, 2010). Furthermore there is a 2-year time lag of the European data analysis.
Resistance trends per antimicrobial
Yearly trends in resistance per antibiotic are shown in Figure 1. Ampicillin resistance fluctuated around the same resistance levels for all three species-matrix combinations, with an average resistance proportion of 13.1% and 30.8% for C. coli from pork and poultry meat, respectively, and 37.4% for C. jejuni from poultry meat. Tetracycline resistance showed yearly fluctuations around the same level for all three species-matrix combinations, with a significantly higher average resistance (p-value<0.05) for C. coli (76.9% and 84.1% for isolates from poultry and pork meat, respectively) compared to C. jejuni (average resistance proportion of 40.8%).
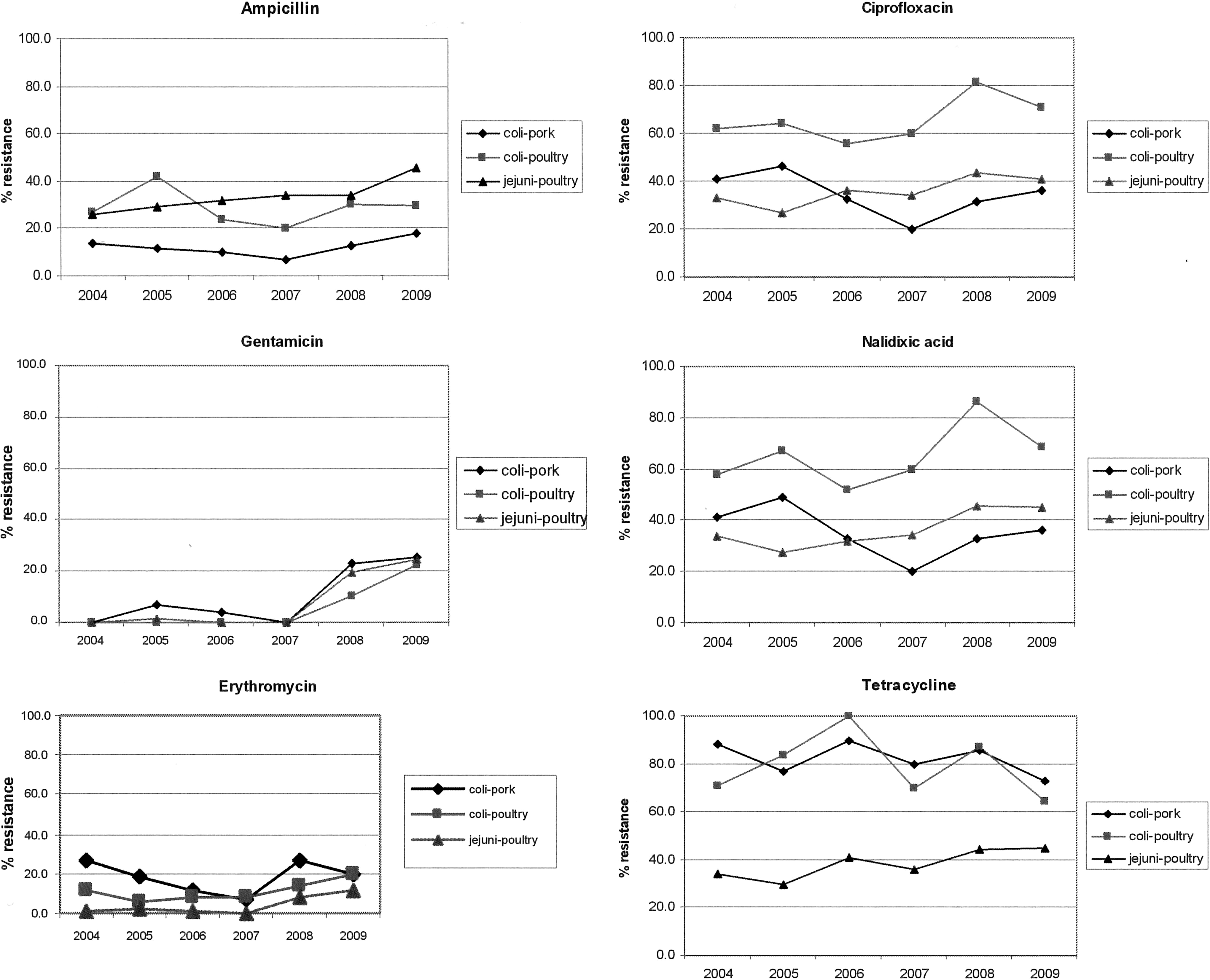
Yearly trends of resistance to ampicillin, ciprofloxacin, nalidixic acid, tetracycline, gentamicin, and erythromycin for Campylobacter coli isolated from pork, and C. coli and C. jejuni from poultry. The epidemiological cutoff values as stated in the text were used.
Gentamicin resistance showed a significant increase between 2007 and 2009 for all three species-matrix combinations, from 0% in 2007, to over 20% in 2009. The most recent surveillance data from the European summary report does not show this high gentamicin resistance among C. jejuni from broiler meat (EFSA, 2010). None of the member states reported any gentamicin resistant isolates from broiler meat (EFSA, 2010). The observed increase of resistance against aminoglycoside antibiotics (like gentamicin) in all species-matrix combinations highlights the potential risk on further spread of the resistance among human pathogens. Since the introduction of the aminoglycoside apramycin for agricultural use (early 1980s), resistance to apramycin has emerged among Escherichia coli isolates found in cattle and pigs in France and the United Kingdom (Chaslus-Dancla and Lafont, 1985; Wray et al., 1986 ). The aminoglycoside 3-N-acetyltransferase type IV resistance gene (AAC(3)IV) and 1-N-acetyltransferase (AAC(1)) confer resistance to several aminoglycoside antimicrobials (apramycin, tobramycin, gentamicin, kanamycin, and neomycin) and were first observed in animal isolates after the introduction of apramycin usage (Hedges and Shannon, 1986). Although apramycin has never been used for the treatment of infections in humans, these resistance determinants were subsequently found in human clinical isolates of E. coli, Salmonella enterica, Klebsiella pneumoniae, and Campylobacter spp. (Johnson et al., 1994; Gomez-Lus et al., 1999). This indicates that the usage of apramycin in agriculture primarily selected for the emergence of the aminoglycoside resistance genes among food animals, which then spread horizontally to human pathogens, where gentamicin is used for treatment.
Trends in resistance to erythromycin of all three species-matrix combinations were stable or decreased from 2004 to 2007, but increased in 2008 and 2009 to a resistance proportion of near 20.0% for C. coli and 12.1% for C. jejuni. Looking at the (fluoro)quinolone resistance proportions, the yearly trends for both Campylobacter species and both matrices were similar for both ciprofloxacin and nalidixic acid. Resistance in C. jejuni from poultry meat and C. coli from pork fluctuated around 40% for the whole study period. C. coli from poultry meat showed a consistently higher resistance for both antimicrobials, fluctuating around 60% for 2004–2007, peaking in 2008 (81.4% Cip and 86.0% Nal), and slightly decreasing in 2009. Belgian C. jejuni fluoroquinolone resistance proportions were significantly higher (p-value<0.05) than reported in poultry meat in Denmark (19% fluoroquinolone resistance), yet similar to poultry meat imported in Denmark (53% fluoroquinolone resistance), yet lower than observed in poultry meat from Austria and Latvia (65% and 100% fluoroquinolone resistance, respectively) (DANMAP, 2008; EFSA, 2010). The observed trends of resistance proportions for all antimicrobials, specifically the distinct peaks for 2008, could result from changes in therapeutic use of these antimicrobials in animal food production during 2008–2009. Unfortunately, this hypothesis cannot be corroborated since detailed surveillance data of antimicrobial use in Belgian agriculture are not yet available.
Although antimicrobial resistance was monitored as acquired reduced susceptibility (using EUCAST epidemiological cut-off values), which cannot be used to deduce clinical implications, these observed high resistance proportions could have important human health consequences as erythromycin and ciprofloxacin are the preferred treatment for diagnosed cases of campylobacteriosis (Belanger and Shryock, 2007). In addition, most diarrheic patients that seek medical attention—and are not diagnosed with campylobacteriosis—are empirically treated with a broad-spectrum antimicrobial (Belanger and Shryock, 2007), most frequently with fluoroquinolones (Engberg et al., 2004), for which especially C. coli from poultry meat shows a high proportion of resistance.
Resistance trends per species and matrix
Average resistance per species and matrices were calculated for the whole study period based on MIC distribution (Table 3). In general, resistance proportions of C. coli and C. jejuni to the tested antimicrobials from poultry meat are in agreement with the average European results per year, which were based on the results of only a few member states (five for C. jejuni and three for C. coli) and to which Belgium contributed antimicrobial resistance results of 2006-2009. In Belgium during the study period 2004-2009 comparison of the two monitored meat matrices revealed significantly higher average resistance proportion (p-value<0.05) to tetracycline (84.1% for pork and 76.9% for poultry) and erythromycin (17.9% for pork and 13.0% for poultry) for C. coli than C. jejuni (40.8% and 6.0% respectively). Significant differences in resistance proportion between matrices was not observed, except for ciprofloxacin and nalidixic acid, for which C. coli isolates from poultry showed higher resistance proportion (69.5% and 69.8% respectively) compared to C. coli from pork (both 35.5%). Furthermore, significantly higher resistance to ampicillin was observed in poultry isolates (30.8% for C. coli and 37.4% for C. jejuni), compared to pork isolates (13.1% for C. coli). Although detailed surveillance data on antimicrobial use in agriculture in Belgium are not available, some reports show trends which could explain the observed difference of fluoroquinolone resistance between pork and poultry food matrices. A study conducted by Timmerman et al. (2006) on the antimicrobial drug consumption at 821 Belgian pig herds revealed that nearly no fluoroquinolones are used in pig husbandry (TIUDD<0.1, with TIUDD being the treatment incidence based on the Used Daily Dose or the number of pigs treated/1000 pigs at risk/day). The antimicrobial use in Belgian broiler production shows significant use of fluoroquinolones (enrofloxacin, TIUDD=6.1; flumequine, TIUDD=1.39) (Persoons and Dewulf, 2010). These data are consistent with results from monitoring programs on antimicrobial use of Denmark and The Netherlands (DANMAP, 2008; MARAN, 2007). Similarly, the use of β-lactam antimicrobials is higher in poultry- than in pig-husbandry (TIUDD 37.9 and 28.9, respectively) (Persoons and Dewulf, 2010; Timmerman et al., 2006).
Results from 2004–2009 were summated.
European Committee on Antimicrobial Susceptibility Testing epidemiological cut-off values were used to assign resistance.
Average %R.
Resistance (%) per year.
Multiresistance
Multiresistance (defined as resistance to four or more of the monitored antimicrobials) of the tested Campylobacter isolates is shown in Figure 2. Multiresistance of C. coli from poultry was on average high (27.6%), but showed strong fluctuations during the monitoring period. C. coli from pork and C. jejuni from poultry showed significantly less multiresistance, with less variation (11.2% and 15.8%, respectively). For both C. coli and C. jejuni from poultry, isolates resistant to all tested antimicrobials were observed (0.8% and 1%, respectively), whereas none were found in pork. Pan-susceptibility was significantly lower in C. coli (8% and 5% in pork and poultry, respectively), compared to C. jejuni (29% in poultry).
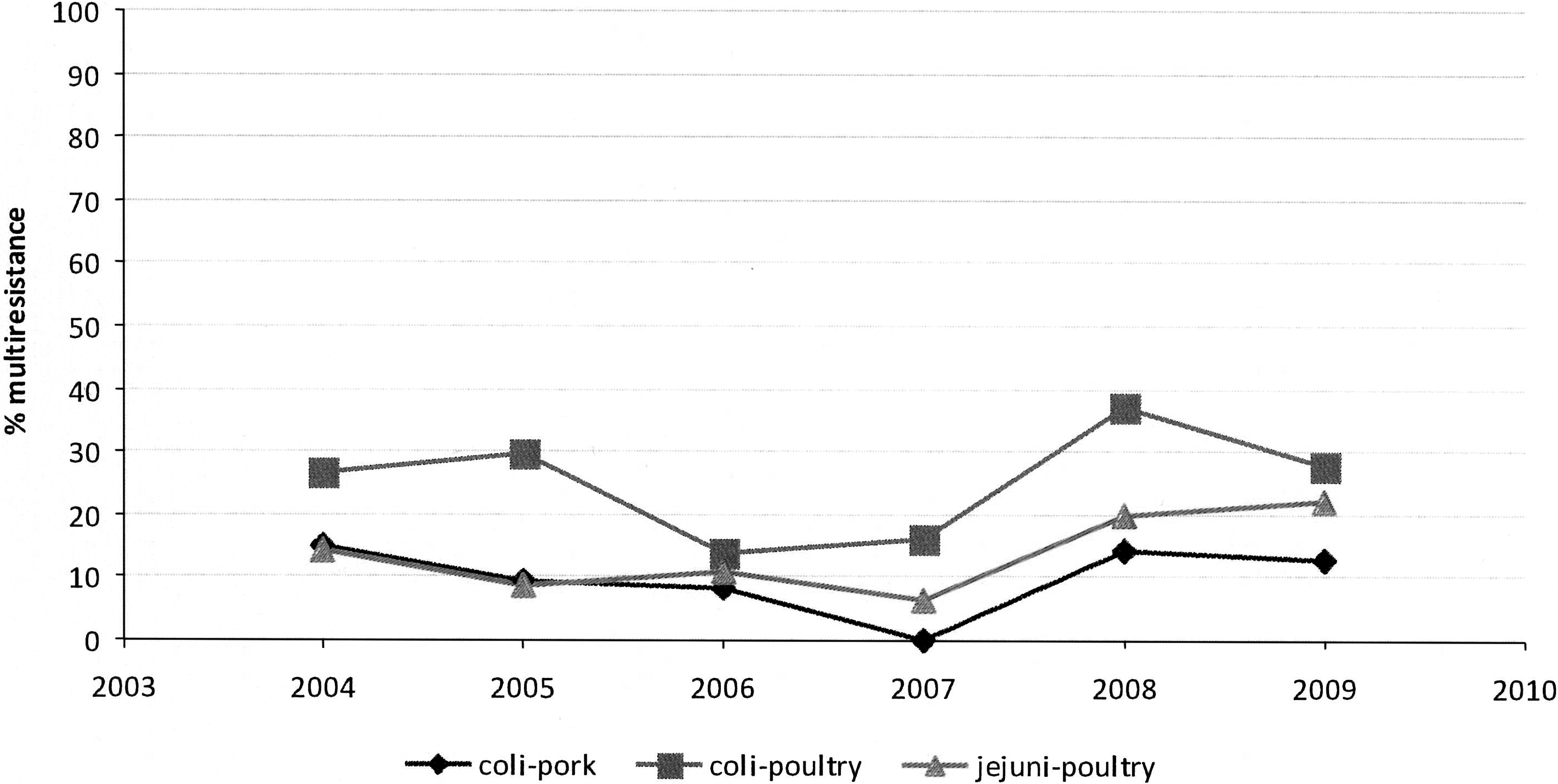
Yearly trends in multiresistance per species-matrix combination. Multiresistance is defined as resistance to four or more of the tested antibiotics.
Resistance to (fluoro)quinolones (like ciprofloxacin and nalidixic acid) in Campylobacter spp. is mainly caused by mutations in the DNA gyrase A gene (Payot et al., 2006). Unlike in other Gram-negative bacteria, where high level resistance is acquired by stepwise accumulation of point mutations, a single mutation in the quinolone-resistance-determining region (QRDR, C257T mutation) suffices to confer resistance (Luo et al., 2003). For 97.5% of isolates, out of a selection of 89 ciprofloxacin-resistant strains, the C257T mutation was confirmed by mismatch amplification mutation assay–PCR (MAMA-PCR). Only two ciprofloxacin-resistant strains showed the wild-type gyrA gene with a cytosine at position 257.
The most common mechanism for macrolide resistance (like erythromycin) is target modification through point mutations in the 23S rRNA genes (A2074C and/or A2075G), of which there are three copies. MAMA-PCR showed the presence of the A2075G mutation in approximately 60% of the selected erythromycin-resistant strains, while the A2074C mutation was absent in all isolates. The sequence of the 23S rRNA gene of the remaining 40% of erythromycin-resistant strains was determined. All strains showed the wild-type adenine at position 2074 and 2075, indicating that another mechanism is responsible for the observed erythromycin resistance. Both laboratory studies and surveillance programs showed that fluoroquinolone resistance-conferring mutations in the gyrA gene of Campylobacter spp. result in a fitness advantage (Luangtongkum et al., 2009). For instance, Luo et al. (2005) observed that quinolone-resistant C. jejuni isolates repeatedly outcompeted susceptible strains in the gut of a chicken animal model, demonstrating the positive effect of resistance on the fitness of this strain. On the other hand, resistance to macrolides are reported to cause a fitness burden, resulting in macrolide-resistant strains that will be outcompeted by natural strains once antibiotic pressure disappears (Caldwell et al., 2008). Regardless of the potential biological relationship between virulence and antimicrobial resistance in Campylobacter spp., the clinical evidence suggests a bigger health burden due to infections caused by resistant strains (Aarestrup et al., 2008).
Furthermore, Campylobacter spp. develop resistance to β-lactam antimicrobials by expression of β-lactamases (chromosomal OXA-type) and to tetracyclines through expression of the ribosome conformational modifying protein Tet(O) (Avrain et al., 2004). However, apart from these individual resistance mechanisms, the expression of the energy-dependent multidrug efflux system CmeABC is known to be widespread among Campylobacter spp. (Olah et al., 2006). Studies have shown that CmeABC expression confers resistance to fluoroquinolones, β-lactams, erythromycin, and tetracycline (with an increase of MIC by 8-, 32-, 4-, and 8-fold, respectively) (Lin et al., 2002). The effect of CmeABC on aminoglycoside resistance (like gentamicin) is less apparent (Lin et al., 2002). The most common antimicrobial multiresistance profiles found in our monitoring program were Amp-Cip-Nal-Tet and Cip-Nal-Ery-Tet, which are in agreement with the reported effect of the CmeABC multidrug efflux system.
Conclusion
Our study demonstrated an overall high antimicrobial resistance in Campylobacter isolates from meat. A remarkable increase of gentamicin resistance has been observed since 2007, which has thus far been unreported in poultry meat. Significantly more fluoroquinolone resistance is observed in meat from poultry compared to meat from pig, which is in agreement with the more frequent use of these antimicrobials in poultry husbandry. The C257T mutation of the gyrA gene was confirmed for nearly all fluoroquinolone-resistant isolates, whereas 40% of the erythromycin-resistant isolates in both C. jejuni and C. coli have the wild-type 23S rRNA gene. The high and increasing resistance proportions observed in this study, especially in poultry, combined with the high prevalence of Campylobacter in poultry carcasses at retail (25%, Table 1) are worrisome.
Footnotes
Acknowledgments
We thank Veronique Real for the technical assistance with determining the MIC values of the Campylobacter isolates.
Disclosure Statement
No competing financial interests exist.