
Abstract
This study aimed to evaluate the antibacterial activities of the essential oils from Origanum vulgare L. (OV) and Rosmarinus officinalis L. (RO), both singly and in combination at sub-inhibitory concentrations (¼ MIC + ¼ MIC), against Aeromonas hydrophila and to investigate the possible mechanisms underlying these activities. Used singly (OV: 2.5 μL/mL; RO: 20 μL/mL) or in a mixture (OV: 0.625 μL/mL + RO: 5 μL/L), these essential oils led to a significant decrease (p<0.01) in bacterial viability after 24 h of exposure. A decrease in glucose consumption by A. hydrophila and release of cellular material were observed immediately after the addition of the essential oils, both singly and as a mixture, and continued for up to 6 h. Electron microscopy of cells exposed to the essential oils revealed severe changes in the plasma membrane, cytoplasmic appearance, and cell shape during the 6-h exposure period. OV and RO essential oils combined at sub-inhibitory concentrations could be rationally applied to inhibit the growth of A. hydrophila in food products, particularly minimally processed vegetables.
Introduction
A. hydrophila is capable of growing at low temperatures, such as those used to preserve minimally processed vegetables (MPVs) (Szabo et al., 2000). The MPV industry has expanded in the past few years in response to a growing demand for the consumption of convenient, fresh, and ready-to-eat foods. However, the emergence of pathogenic microorganisms, including A. hydrophila, that have not been previously linked with raw or processed food products has enhanced the potential for outbreaks of foodborne diseases associated with the consumption of MPVs (Monge et al., 1998; McMahon and Wilson 2001; Uyttendaele et al., 2004).
To improve the safety and shelf life of MPVs, researchers in the food industry have applied several different sanitizers for controlling the initial amount of spoilage and pathogenic microorganisms in these foods (Gomez-López et al., 2005). Hypochlorite is often applied to MPVs for disinfection, although this procedure has presented some disadvantages, such as the formation of carcinogenic chlorinated compounds and poor antimicrobial efficacy against some microorganisms (Singh et al., 2002; Allende et al., 2008), including A. hydrophila (Uyttendaele et al., 2004).
In this context, plant essential oils have received particular interest for their potential as antimicrobial compounds during the treatment of MPVs (Guttierrez et al., 2008). Early studies have shown that the essential oils from Origanum vulgare L. (OV) and Rosmarinus officinalis L. (RO) possess antimicrobial activities that combat spoilage and pathogenic food-related microorganisms (Sacchetti et al., 2005; Souza et al., 2007; Muñoz et al., 2009; Oliveira et al., 2010). However, researchers have shown that these essential oils display antimicrobial properties in synthetic media, but the amounts required to substantially inhibit bacterial growth in food matrices are often higher than would be organoleptically acceptable (Naveena et al., 2006; Souza et al., 2009). Because these high concentrations are likely to impart a certain flavor to foods, the addition of sub-inhibitory levels of essential oils as mixtures should be adjusted to find a balance between sensory acceptability and antimicrobial efficacy (Dimirtiević et al., 2007).
In a previous study, Azerêdo et al. (2011) found MIC values of 2.5 and 20 μL/mL for OV and RO, respectively, against A. hydrophila INCQS 7966. In the same study, analysis of the fractional inhibitory concentration index for the combined application of these essential oils suggested a synergistic interaction against this bacterium and great inhibition in vegetables. The sensory evaluation of vegetables sanitized with a mixture of these essential oils at sub-inhibitory concentrations (¼ MIC+¼ MIC) indicated increased acceptability of most of the sensory attributes assessed after refrigerated storage when compared to vegetables sanitized with each essential oil independently (at the MIC). In this manuscript, we report further investigations of the effects of OV and RO essential oils, used singly and in combination at sub-inhibitory concentrations, on the cell viability, glucose consumption, membrane permeability and morphology of A. hydrophila. To our knowledge, there is a lack of studies emphasizing the antimicrobial mechanism of the application of a mixture of essential oils at sub-inhibitory concentrations against pathogenic microorganisms that are associated with foods.
Methods
Essential oils
Essential oils from OV (batch OREORG01; density at 20°C: 0.90; refractive index at 20°C: 1.47) and RO (batch ROSTUN04; density at 20°C: 0.94; refractive index at 20°C: 1.51) were obtained from Aromalândia Ind. Com. Ltda. (Minas Gerais, Brazil), and the quality parameters of the oils were described in an accompanying technical report. This supplier extracts the essential oils on an industrial scale by steam distillation. Essential oil solutions were prepared in a nutrient broth (Difco, Juiz de Fora, Brazil) using bacteriological agar (0.15 g/100 mL) as a stabilizing agent (Mann and Markam, 1998; Bennis et al., 2004).
Bacterial strain
A. hydrophila INCQS 7966 was obtained from the Brazilian Institute of Quality Control in Health (Rio de Janeiro, Brazil) and used as the test microorganism. A stock culture was kept on nutrient agar (NA) slants under refrigeration (7±1°C). Inocula used in the antimicrobial assays were obtained from overnight cultures grown on NA slants at 28°C. Unless stated otherwise, the inocula used in the assays were obtained from suspensions of the strain grown to stationary phase and were prepared by inoculating 400 mL of Brain Heart Infusion (BHI) broth (Difco) with two colonies of each strain from an overnight culture on BHI agar (Difco). The newly inoculated culture was then incubated at 30°C for 18 h. After incubation, the bacteria were separated from the growth medium by centrifugation at 10,000 x g for 12 min at 4°C, washed twice with phosphate-buffered saline (PBS; pH 7.4), and resuspended in PBS. Suspensions were adjusted so that the optical density at 620 nm (OD620) of a 1:100 dilution was approximately 0.3, corresponding to approximately 3×1010 colony forming units per milliliter (CFU/mL) (Carson et al., 2002). The suspension was serially diluted in PBS (10−1 to 10−3) to provide a viable cell count of approximately 107 CFU/mL.
Time-kill assays
The effects of the essential oils used singly (OV: 2.5 μL/mL; RO: 20 μL/mL) or in combination (OV: 0.625 μL/mL; RO: 5 μL/mL—OV/RO) on the cell viability of A. hydrophila were evaluated by the viable cell count procedure. For this assay, 4 mL of nutrient broth was inoculated with 1 mL of the bacterial inoculum, and then 5 mL of the essential oil solution was added to the mixture. The culture was gently shaken for 30 s and then incubated at 30°C. At several time intervals (0, 1, 2, 4, 8, 12, and 24 h), 1 mL of the suspension was serially diluted (10−1 to 10−5) in PBS and used to inoculate sterile nutrient agar Petri dishes, which were subsequently incubated for 24 h at 30°C (Barros et al., 2009). Control flasks without essential oils were tested under the same conditions. The results are expressed as the log of the CFU/mL value.
Glucose consumption
Aliquots of 100 μL of the bacterial inocula were exposed to the essential oils singly (OV: 2.5 μL/mL; RO: 20 μL/mL) or in combination (OV: 0.625 μL/mL; RO: 5 μL/mL—OV/RO) in PBS supplemented with glucose (0.2 g/L) at 30°C. At several time intervals (0, 30, 60, 90, 120, 240, and 360 min), an aliquot (1 mL) of the mixture was removed, and the extracellular glucose consumption (mg/100 mL) was estimated using the photometric Glucose PAP Liquiform kit (Labtest Diagnóstica S.A., Minas Gerais, Brazil). Control flasks without essential oils were also tested (Tassou et al., 2000).
Release of cellular material
Measurement of the release of material absorbing at 260 nm from A. hydrophila was conducted using 2-mL aliquots of the bacterial inocula in PBS after the addition of essential oils, either singly (OV: 2.5 μL/mL; RO: 20 μL/mL) or in combination (OV: 0.625 μL/mL; RO: 5 μL/mL—OV/RO), at 30°C. At several time intervals (0, 30, 60, 90, 120, 240, and 360 min), the bacterial cells were separated from the growth medium by centrifugation at 10,000 x g for 12 min at 4°C, and the supernatant was examined for leakage of cellular material by measuring the absorbance at 260 nm with a Biochrom Libra S32/S32 spectrophotometer, as described by Carson et al. (2002). Control flasks without essential oils were also tested.
Ultrastructural analysis
Cells exposed to either individual essential oils (OV: 2.5 μL/mL; RO: 20 μL/mL) or a mixture of the oils (OV: 0.625 μL/mL; RO: 5 μL/mL—OV/RO) for 3 or 6 h were harvested by centrifugation at 10,000 x g for 12 min at 4°C, washed in PBS, and fixed for 24 h at 4°C in PBS with glutaraldehyde (0.25 g/L). After being washed in the same buffer, the cells were post-fixed for 30 min with osmium tetroxide (0.01 g/L) in 0.1 M cacodylate buffer (pH 7.2) and allowed to adhere to poly-lysine-coated cover slips. The samples were dehydrated with ethanol followed by critical point drying with CO2, coated with a 20-nm-thick gold layer, and observed with an FEI Quanta 200 FEG scanning electron microscope (Battistelli et al., 2005). For transmission electron microscopy, cells exposed to the essential oils were fixed and post-fixed as described above, dehydrated in a graded acetone series, and embedded for 72 h at 60°C in Poly/Bed 812 resin (PolySciences, Warrington, PA). Ultrathin sections were stained with uranyl acetate and lead citrate and were observed with a Morgagni 268D transmission electron microscope (Olivotto et al., 2008). Bacterial cells that were not exposed to the essential oils were fixed and observed similarly as a control.
Statistical analysis
All assays were performed in triplicate on two separate occasions, and the results are expressed as an average of the assays. Statistical analysis was performed to determine significant differences (p<0.01) by analysis of variance followed by Tukey's test. For statistical analysis, the SigmaStat 3.1 computer program was used.
Results
Time-kill assays
Individual addition of the essential oil of OV (2.5 μL/mL) or RO (20 μL/mL) caused a sharp drop in the bacterial count over the evaluated periods of time (Fig. 1). Values lower than two log cycles were found for A. hydrophila in broth containing only the essential oil of OV after 4 h, and no recovery in viable cell count was noted in the remainder of the evaluated intervals.

Survivor curves for Aeromonas hydrophila INCQS 9610 in nutrient broth at 28°C added with Origanum vulgare (OV) and Rosmarinus officinalis (RO) singly and in mixture: (+) OV: 2.5 μL/mL; (○) RO: 20 μL/mL; (▵) OV: 0.62 μL/mL+RO: 5 μL/mL; (▪) control: 0 μL/mL. Detection limit of the assay: 2.0 Log of CFU/mL.
The combination of both essential oils at sub-inhibitory concentrations (OV: 0.625 μL/mL and RO: 5 μL/mL) reduced the initial inocula of A. hydrophila by four log cycles after 12 h of exposure, and no recovery in viable bacteria was noted after 24 h of exposure. A smaller decrease in the viability of A. hydrophila was found in the broth containing only the RO essential oil. Application of the essential oils both singly or in combination caused a significant decrease (p<0.01) in bacterial counts over the evaluated intervals in comparison to the control assay.
Effects of essential oils on glucose consumption
The amount of glucose in the broth without essential oils progressively decreased, indicating rapid consumption of glucose by the A. hydrophila cells over time (Fig. 2). On the other hand, similar amounts of glucose (p>0.01) were found in broth containing essential oils (both singly and in combination) over the assessed time period. These results suggest that the glucose consumption of A. hydrophila was affected immediately upon the addition of the essential oils, followed by a steady inhibitory effect. The addition of the essential oils, both singly and in combination, caused a significant decrease (p>0.01) in the consumption of glucose by A. hydrophila when compared to the control assay.
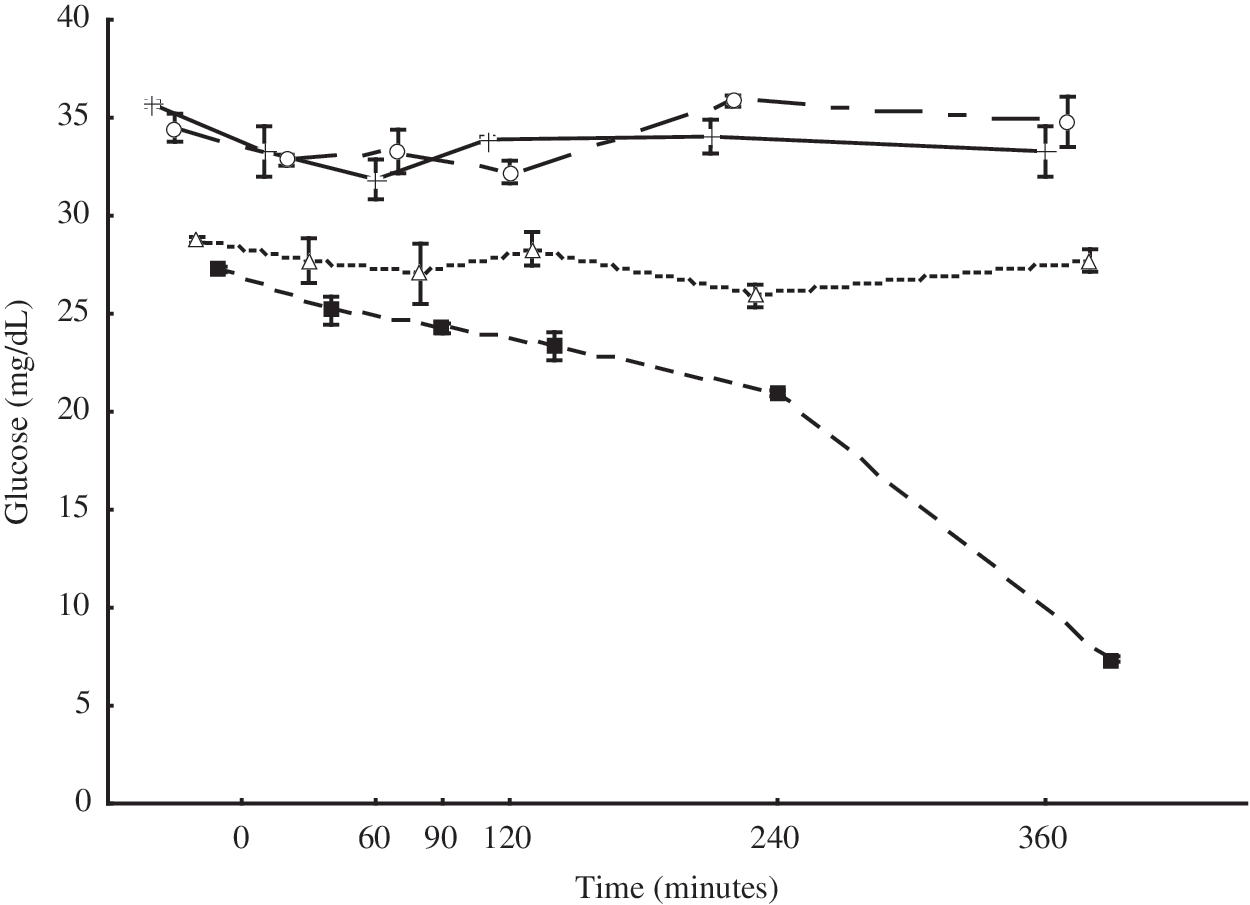
Inhibitory effect on glucose consumption of Aeromonas hydrophila cells by exposure to essential oils of Origanum vulgare (OV) and Rosmarinus officinalis (RO) singly and in mixture. (+) OV: 2.5 μL/mL; (○) RO: 20 μL/mL; (▵) OV: 0.62 μL/mL+RO: 5 μL/mL; (▪) control: 0 μL/mL.
Release of cellular material
Measuring the optical density at 260 nm of the suspension of cells that were exposed to the essential oils, both singly and in combination, revealed an increase in the release of material absorbing at 260 nm throughout the period of exposure (p<0.01; Fig. 3). These results suggest that increased membrane permeability at least partially underlies the antimicrobial property of the tested essential oils against A. hydrophila.

Rate of 260-nm-absorbing material release from Aeromonas hydrophila exposed to essential oils of Origanum vulgare (OV) and Rosmarinus officinalis (RO) singly and in mixture. (+) OV: 2.5 μL/mL; (○) RO: 20 μL/mL; (▵) OV: 0.62 μL/mL+RO: 5 μL/mL; (▪): control: 0 μL/mL.
Ultrastructural analysis
Scanning electron microscopy of A. hydrophila cells revealed morphological damage due to the exposure to the essential oils of OV and RO, both singly and in combination for 3 or 6 h (Fig. 4A–F). Relevant morphological changes occurred in the presence of OV and RO essential oils (both singly and in combination) after only 3 h of exposure. Normal shaped cells disappeared, and elongated and distorted forms appeared after only 3 h of incubation. Microscopy of the cells exposed to the essential oils revealed membrane damage with the appearance of projecting cellular material. Bacteria grown in control flasks (no essential oils) presented normal bacillary morphology after both 3 and 6 h of incubation (Fig. 4G–H).
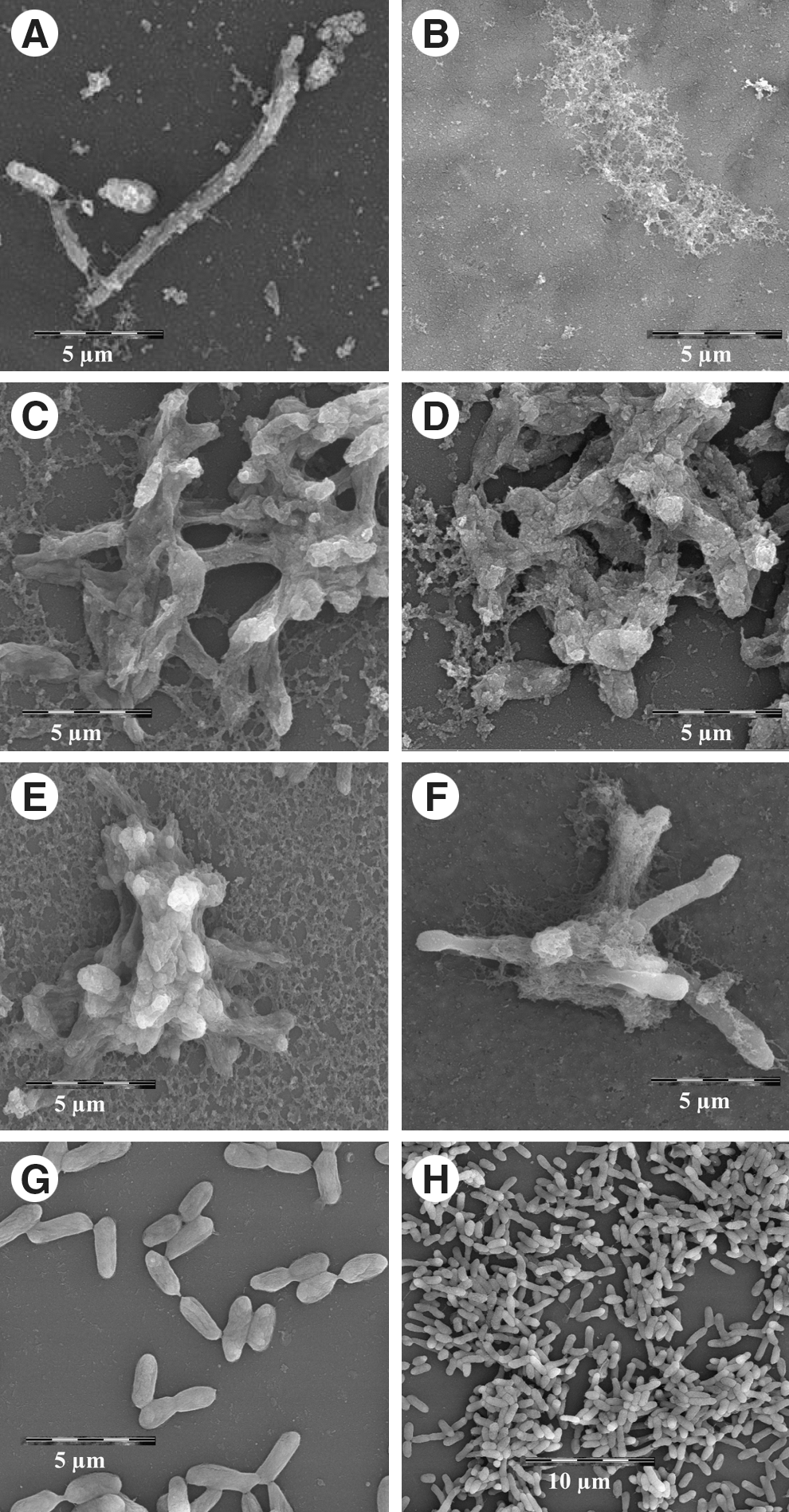
Scanning electron microscopy of Aeromonas hydrophila cells incubated with essential oils of Origanum vulgare (OV) and Rosmarinus officinalis (RO) alone and in mixture.
A. hydrophila cells, when exposed to the essential oils both singly and in combination, demonstrated changes to the outer membrane, cytoplasmic appearance, and cell shape (Fig. 5A–F). Exposure to the essential oil of OV alone caused cell deformation, condensation and loss of cytoplasmic material after 3 h of exposure (Fig. 5A), and after 6 h, the essential oil caused total collapse of the structural integrity of the cells (Fig. 5B). Similar changes were found when the cells were exposed to the essential oils in combination (Fig. 5E,F). Cells exposed to the RO essential oil displayed detachment of the cell wall from the cytoplasmic membrane, damage to the outer membrane and loss of cytoplasmic material after only 3 h of incubation (Fig. 5C). Cells grown in control flasks (no essential oils) appeared normal in shape, with intact outer membranes and evenly distributed cytoplasm (Fig. 5G,H).
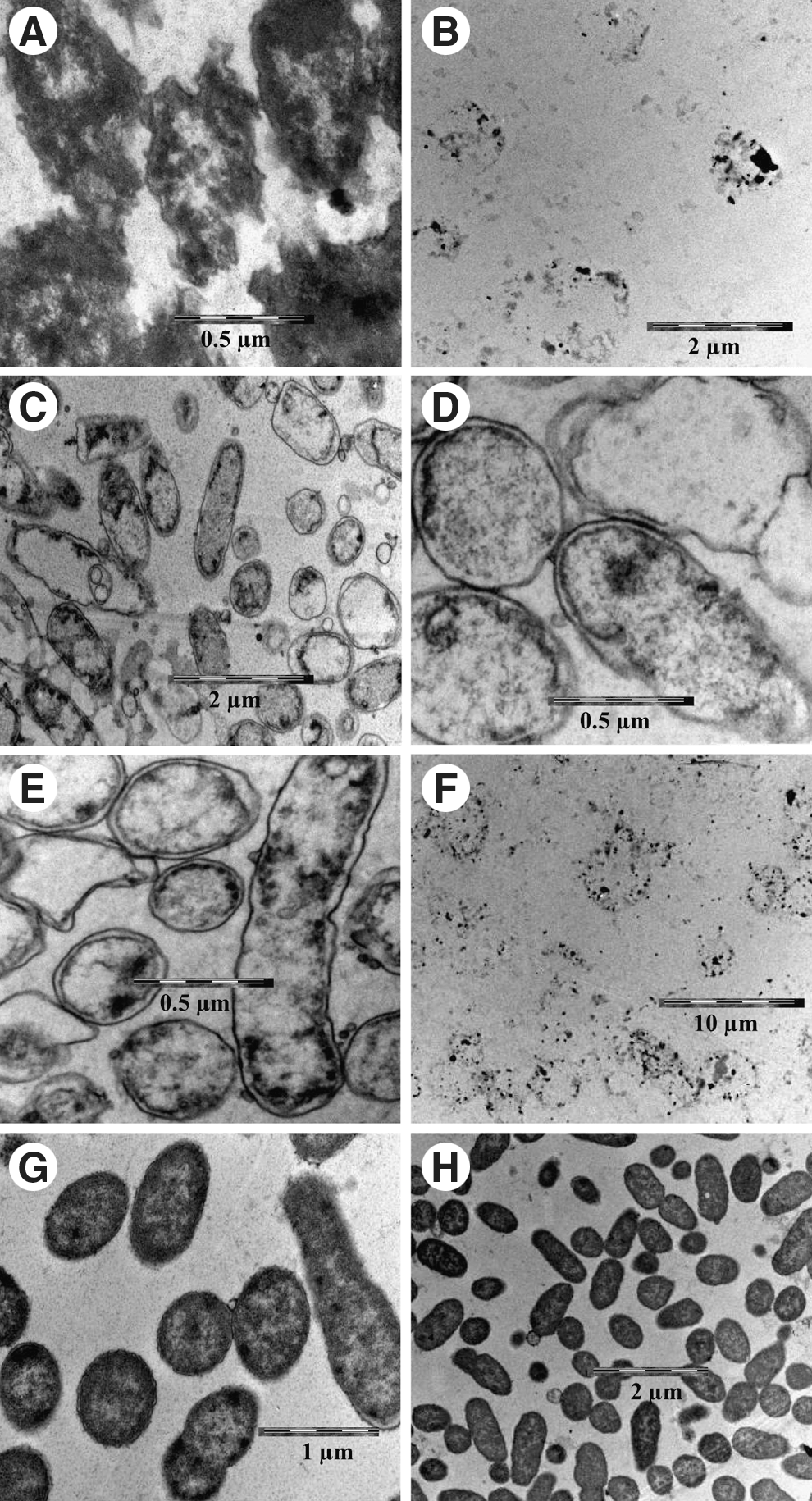
Transmission electron microscopy of Aeromonas hydrophila exposed to essential oils of Origanum vulgare (OV) and Rosmarinus officinalis (RO) alone or in combination.
Discussion
OV and RO essential oils, used singly and in combination at sub-inhibitory concentrations, severely inhibited the cell viability of A. hydrophila INCQS 7966 after 24 h of exposure. Few studies have previously investigated the efficacy of essential oils in inhibiting the cell viability of A. hydrophila. Uyttendaele et al. (2004) reported a decrease in the counts of A. hydrophila in fresh vegetables when exposed to thyme essential oil at 5 and 10 μL/mL.
A. hydrophila cells showed a decrease in glucose consumption after 360 min of exposure to the essential oils of OV and RO, both singly and in combination. According to Tassou et al. (2000), these findings may indicate inhibition of either glucose uptake or the glucose metabolic pathway.
Exposure of A. hydrophila cells to essential oils caused a rapid release of material absorbing at 260 nm. Demonstrable leakage of cytoplasmic material is indicative of gross and irreversible damage to the cytoplasmic membrane (Cox et al., 1998). In particular, such damage may occur as a result of the disruption of peptidoglycan (Ruiz-Barba et al., 1990) and/or damage to the cell membrane (Hugo, 1991).
Scanning electron microscopy of A. hydrophila cells grown in the presence of the essential oils revealed drastic changes in cell morphology with damage to the cytoplasmatic membrane accompanied by obvious surface alterations. Cells exposed to the essential oils, both singly and in combination, had elongated shapes after 3 h of exposure. Elongated cells are regarded as viable because they are capable of dividing and forming new, normal rod cells after exposure to stress-inducing conditions (McMahon et al., 2007). Researchers have proposed that elongation is a possible strategy for coping with unfavorable conditions (Piuri et al. 2005; Pianetti et al., 2008; Pianetti et al., 2009). No bacteria were observed after 6 h of cultivation in the presence of OV essential oil alone or a mixture of OV and RO essential oils, indicating that these treatments effectively killed A. hydrophila.
Observations from transmission electron microscopy suggested that the assayed essential oils also caused changes in the cell wall and outer membrane of A. hydrophila. Components of the essential oils could easily enter into the cells through these lesions, facilitating the leakage of cellular components and inducing cell lysis. Visualization of the damage to the cell structure confirms the sensitivity of the outer membrane of A. hydrophila to the tested essential oils. These oils may penetrate the cytoplasmic membrane, influencing membrane permeability and resulting in the leakage of cytosolic material over a short period of exposure. These results are in agreement with observations of other gram-negative bacteria, in which stress conditions lead to similar morphological changes (Pianetti et al., 2009; Lacombe et al., 2010).
We have previously found that carvacrol (66.9 g/100 g) and 1,8-cineole (32.2 g/100 g) are two major compounds (57.71%) of the essential oils from OV and RO that were assayed here (Azerêdo et al., 2011). It is difficult to understand the exact mechanism underlying the enhanced antimicrobial effect of the combined application of OV and RO essential oils, although the increased antimicrobial activity caused by the mixture of these essential oils may partially be explained by considering the different compounds found in each individual essential oil. Essential oils with the strongest antibacterial activities, such as the essential oil of OV, generally present high amounts of carvacrol and/or thymol. Hydroxyl groups enhance the antibacterial properties of essential oils, and 1,8-cineole, which was the most prevalent compound in the essential oil of RO, may have also contributed to the rapid and steady antimicrobial effect achieved with the combination of the two oils (Elgayyar et al., 2000).
Cineole, one of the main components of the RO essential oil (Azêredo et al., 2011), possesses oxygenated monoterpenes in its structure, and these structures are known to increase the antimicrobial properties of terpenoids (Naigre et al., 1996). Hydrocarbons (such as α-pinene, camphene, myrcene, α-terpinene, and p-cymene), which have very weak antibacterial activity, are found in the RO essential oil in lower amounts (Azerêdo et al., 2011) and appear to swell bacterial cells to a greater extent than carvacrol. Therefore, these compounds enable carvacrol to be more easily transported into the cell (Guttierrez et al., 2008).
According to our results, the essential oils from OV and RO, used singly and in combination at sub-inhibitory concentrations, inhibited the cell viability of A. hydrophila. The observation of the release of cytoplasmic material, decreased glucose consumption, and altered morphology of the cells suggests that the cell wall and cytoplasmic membrane of A. hydrophila are compromised by exposure to these essential oils. Overall, these results reveal that the combination of OV and RO essential oils at sub-inhibitory concentrations could be rationally applied to inhibit the growth of A. hydrophila in food products, particularly in vegetables, to balance the microbial safety and organoleptic acceptability of food products.
Footnotes
Disclosure Statement
No competing financial interests exist.