
Abstract
The objective of this study was to determine persistence of clonal strains from farm to retail by assessing the clonal relatedness of Campylobacter coli isolated on farm, peri-harvest, and at processing from 11 individually identified pigs. Phenotypic (antimicrobial susceptibility) and genotypic (pulsed field gel electrophoresis [PFGE] and multi-locus sequence typing [MLST]) characterization of isolates was conducted. There was high genetic diversity of Campylobacter isolates from on-farm fecal samples. Campylobacter isolates from farm, post-evisceration, hide, and carcass samples showed similar phenotypes and belonged to the same genotypic clusters based on PFGE and sequence types (STs) based on MLST. Five STs that have not been previously reported were identified (ST-4083, ST-4084, ST-4085, ST-4086, ST-4087). Despite high genotypic diversity of C. coli on farm, retail meat products were consistently contaminated with isolates of the same STs, particularly ST854 and ST1056, as isolates collected from previous stages confirming persistence of strains from pre- to post-harvest.
Introduction
Genotypic approaches are routinely implemented to characterize Campylobacter. Pulsed-field gel electrophoresis (PFGE) is the most common fingerprinting method for Campylobacter and other foodborne pathogens. Multi-locus sequence typing (MLST) has increasingly been used in Campylobacter, as it is suitable and discriminatory to characterize bacterial organisms with hypervariable genomes, including Campylobacter (Dingle et al., 2002). To date, only few reports compared clonal relatedness of Campylobacter in swine from the same animal at different stages along the food processing chain. This is especially important in defining the role of pre- harvest factors for contamination of meat. The objective of this study was to determine the phenotypic and genotypic relatedness of C. coli isolates from the same pigs collected from the farm through processing.
Methods
Study design
The isolates originated from a previous study (Abley et al., 2012). One hundred pigs from one farm were individually identified and followed within seven cohorts (10–16/cohort) through the harvest process at one harvest facility. Samples collected from each pig included a fecal sample on farm (farm), rectal contents post-evisceration (post-evis), a hide swab in lairage (hide), carcass swabs immediately after wash and pre-chilling (carcass), and a meat sample (ribs). Inclusion criteria for this study were for pigs from which a C. coli was isolated from each of these sample types. Eleven pigs (55 isolates) met this criterion sampled on five different dates from October 2006 to March 2007 (Table 1). Of these 55 isolates, 43 were recoverable from cryopreservation and included in this study.
Ar, azithromycin; Ca, clindamycin; CIP, ciprofloxacin; Er, erythromycin; Ff, florfenicol; Gm, gentamycin; Nl, naladixic acid; Tt, telithromycin; Te, tetracycline; Pan S., pan-susceptible.
NG, no growth (some of the isolates originally identified for this study based on previous polymerase chain reaction confirmation were not successfully recovered from crypreservation for the initial testing of multi-locus sequence typing and then later a few more were not recovered for pulsed-field gel electrophoresis [PFGE]); OC, outside of PFGE cluster.
Founder Sequence Type (ST) is derived from the Based Upon Related Sequences (BURST) analysis.
Campylobacter culture
Fecal and rectal samples were cultured without pre-enrichment. One gram of feces was diluted in 9 mL of buffered peptone water (BPW; Becton Dickenson, Sparks, MD), and 100 μL was plated in duplicate on Campy-Cefex (Oyarzabal et al., 2007; LaGier et al., 2004) plates. The remaining swab and meat samples were enriched in Bolton broth followed by plating. The dilution for the samples were as follows: hide swab in 30 mL, carcass swabs (five pooled from same pig) in 90 mL, and 1 pound (454 g) of rib meat in 500 mL. All broth enrichments were incubated under microaerophilic conditions for 48 h at 42°C. One hundred microliters from each enrichment broth sample was plated onto a Campy-Cefex plate and incubated under microaerophilic conditions for 48 h at 42°C. Biochemical confirmation was done using catalase (Becton Dickenson) and oxidase (Becton Dickenson) tests. DNA was extracted using the DNeasy Tissue kit (Qiagen). Polymerase chain reaction (PCR) was performed on the extracted DNA targeting the hipO gene for Campylobacter jejuni and the glyA gene for Campylobacter coli (LaGier et al., 2004).
Antimicrobial susceptibility
Antimicrobial susceptibility of isolates was determined by using an approved standard broth microdilution method (CLSI, 2006). The experiment used commercially prepared CAMPY plates (Sensititre; TREK™ Diagnostic Systems Inc., Westlake, OH), and samples were tested according to the manufacturer's instructions. Breakpoint values were based on Clinical and Laboratory Standards Institute (CLSI) and by the U.S. National Antibiotic Monitoring System (NARMS). Minimum inhibitory levels were determined using the SensiTouch (TREK™ Diagnostic Systems Inc). The quality control organism C. jejuni American Type Culture Collection (ATCC) 33560 was used to verify the quality of the plates (CLSI, 2006).
PFGE
PFGE of C. coli to determine genomic DNA fingerprints was performed using the Centers for Disease Control and Prevention Pulse Net Protocol (Ribot et al., 2001). Briefly, C. coli isolates were grown for 48 h under microareophilic conditions at 42°C on blood agar plates (trypticase soy agar with 5% defribrinated sheep blood [Remel, Lenexa, KS]). Bacterial cell suspensions were prepared in phosphate-buffered saline (PBS; EMD Chemicals Inc., Darmstadt, Germany) to an optical density (OD) at 610 nm of 0.57 to 0.82. The cell suspension was then transferred to a microcentrifuge tube containing proteinase K (20 mg/mL) and mixed gently. Agarose-embedded cells were then lysed with cell lysis buffer (CLB; 50 mM Tris and 50 mM EDTA [pH 8.0], 1% sarcosyl [Sigma, St. Louis, MO], 0.1 mg/mL proteinase K), and intact genomic DNA was digested with 50 U of the SmaI restriction enzyme (Fermentas, Glen Burnie, MD) for 24 h at 30°C. The fragments were then separated using the CHEF-DR III PFGE system (Bio-Rad Laboratories, Hercules, CA). The PulseNet standard marker strain, Salmonella enterica serovar Braenderup H9812, was used as a molecular reference digested with XbaI. PFGE results were analyzed using the Bionumerics software version 4 (Applied-Maths, Kortrijk, Belgium), and the banding patterns were clustered using Dice coefficients with a 2% band position tolerance and 1.5% optimization. All isolates with a similarity index of 92% or greater were grouped together based on the similarity of the control strains and out groups (data not shown).
MLST
MLST was performed on the purified DNA for each of the following seven housekeeping genes as described (Dingle et al., 2001; Miller et al., 2005): aspA (aspatase), glnA (glutamine synthetase), gltA (citrate synthase), glyA (serine hydroxy methyl transferase), pgm (phospho glucomutase), tkt (transketolase), and uncA (ATP synthase alpha subunit). Ready-to-Go PCR beads (GE Healthcare Life Sciences, Piscataway, NJ) were used for all PCR amplifications. PCR reaction conditions were initial denaturation at 95°C for 5 min, followed by 30 cycles of the following: 95°C for 1 min, primer annealing at 55°C for 90 sec, and extension at 72°C for 1 min. The PCR products were purified by using a Qiaquick Multiwell PCR purification kit (Qiagen). Sequencing reactions were conducted in a volume of 20 μL containing 1 μL of purified PCR product, 2 μL of primer (10 pmol/μL), 1.5 μL of sequencing buffer (Beckman Coulter, Fullerton, CA), 2 μL of DTCS Quick Start Master Mix (Beckman Coulter), and 13.5 μL of molecular grade water. Thermal cycling conditions for sequencing reactions were set up according to the manufacturer's instructions. Unincorporated dye terminators were removed by ethanol precipitation, and the sequenced products were separated and detected with a CEQ 8000 Genetic Analysis System (Beckman Coulter). Alleles and sequence types (STs) were assigned by submitting each DNA sequence to the Campylobacter MLST database. Additionally, the Clustal W software (Larkin et al., 2007) was used to perform the sequence alignments.
Results
Antimicrobial susceptibility
The most common antimicrobial resistance phenotypes for the five sample types were as follows: (1) farm: pan-susceptible [n=3], and tetracycline (Te) [n=2]; (2) post-evisceration: azithromycin (Ar), Clindamycin (Ca), Erythromycin (Er) and Te [n=3]; (3) hide: Te [n=7]; (4) carcass: Te [n=7]; and (5) meat: Te [n=2] (Table 1).
Several pigs only had multidrug resistance at the farm or at post-evisceration, and not in other isolates from the same animal (pigs 2, 4, 6, 26, 43, and 76). Two pigs had multidrug-resistant C. coli observed in farm isolates and meat isolates (pigs 10 and 31).
PFGE
The PFGE analysis revealed seven distinct genotypic clusters using a cut-off of 92% similarity (Fig. 1). There were two clusters (B and G) that contained isolates from meat and other isolates collected upstream in the processing. Cluster B contains four isolates that were from different pigs and sources, including farm, post-evisceration, and meat. There were two STs in cluster B (ST-1056 and ST-1107), both of which were within the same clonal complex (Fig. 2). The isolates were mostly multidrug resistant, with one isolate being resistant to only tetracycline. Three out of the four isolates were recovered from the same cohort of animals at slaughter. Cluster G contains six isolates from multiple pigs (two from pig 43) and multiple sources (hide, carcass, and meat), all of which were ST-1056 and all but one were resistant only to tetracycline. The other isolate was pan-susceptible and collected from different cohorts.

Dendrogram of pulso-types created by pulsed-field gel electrophoresis clustered using the dice coefficient. Thirty-four isolates recovered from cryopreservation from Campylobacter coli isolates from farm, post-evisceration, hide, carcass, and final meat product from 11 finishing swine. C5560, C6356, and C6005 are samples that did not originate from this study; they were included only as an out-group.
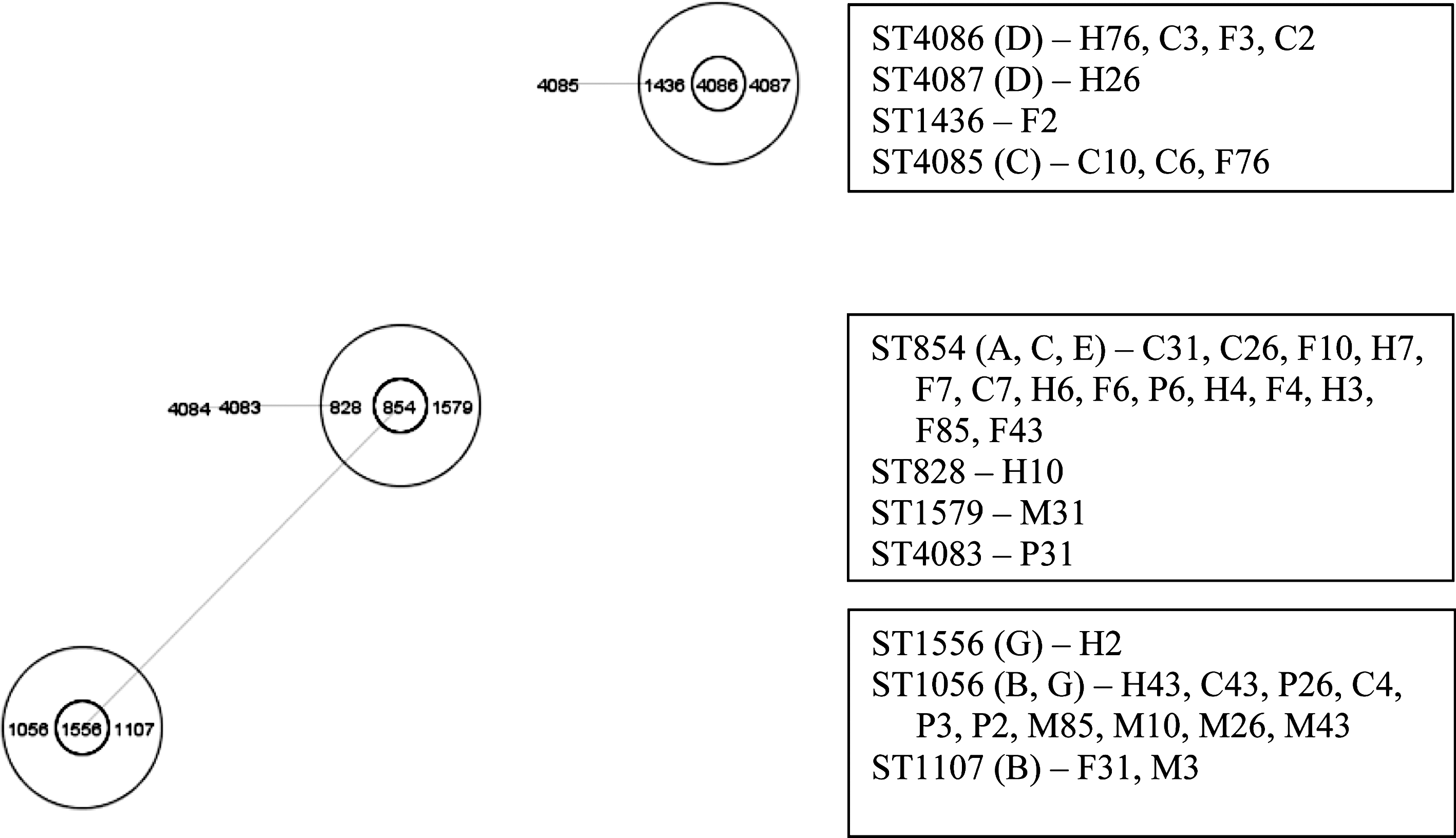
Based Upon Related Sequences analysis for all the sequence types. The sequence type in the center is the founding sequence type for each of the three clonal lineages. The sequence types within the larger circle indicate single locus deviants (SLD) and the lines represent double locus variants (DLV). The boxes describe the source (F, farm; P, post-evisceration; H, hide; C, carcass; M, meat) and pig for each sequence type. The letters in parentheses indicate the pulostype cluster.
MLST
A total of 13 different STs were identified in this study. Among them, ST-854 (n=14) and ST-1056 (n=10) were the most common. These predominant STs were identified among samples from the farm, post-evisceration, hide, carcass, and meat, which indicates their persistence. Five new STs that were not reported previously were identified (ST-4083, ST-4084, ST-4085, ST-4086, and ST-4087).
Discussion
Campylobacter isolates from meat were phenotypically similar and are within the same PFGE cluster (Fig. 1) and MLST clonal lineages (Fig. 2) as isolates collected from farm, post-evisceration, hide, and carcass. This indicates that the isolates on the meat most likely originated from contamination from the feces or hide of the pigs. This study could not determine if the meat was contaminated directly from the feces or the hide, or from the persistence of strains on the equipment and/or hands of the workers at the harvest facility.
The high frequency of resistance to tetracycline (77%) and erythromycin (28%) was similar to results previously reported by our group (Thakur et al., 2005b). Wang et al. (2011) observed a lower percentage of erythromycin resistance in C. coli isolated from retail meat (mostly chicken breasts); the researchers also observed an agreement between MLST and PFGE, but they did not observe any correlation between antibiotic resistance patterns and MLST and PFGE types. The previous study also found PFGE to be more discriminatory than MLST; this was also observed in the present study.
Other studies have observed a high host association with C. coli and swine and also relatively little diversity as far as the majority of isolates belong to the clonal complex 828 (Lang et al., 2010; Sheppard et al., 2010; Ogden et al., 2009); this was also seen in the present study. Also, when the STs found in this study were compared to the global database on the MLST website, all but two STs (1579 and 1056) included isolates deposited from swine. Several STs from this study also were associated with human samples (1556, 1056, 828, and 854).
Some STs exhibited multiple resistance patterns. For instance, ST-1056 exhibited patterns that included pan-susceptible; tetracycline (Te); or multidrug resistance with azithromycin (Ar), clindamycin (Ca), naladixic acid (NI), and Te. ST-854 predominantly exhibited pan-susceptible or Te. This is not entirely unexpected, since MLST genotyping focuses on housekeeping genes, whereas antimicrobial resistance could be encoded in regions with greater plasticity, including plasmids. However, this emphasizes the importance of using a combination of phenotype and genotype to fully characterize Campylobacter isolates for epidemiologic classification. A study conducted by Miller et al. (2007) typed a total of 488 C. coli strains from four different food sources (cattle, chicken, swine, and turkeys). Of the four sources, they observed that swine had the most diverse STs, with 82 unique STs identified within 185 swine samples. Our results found a similar type richness of isolates originating from 11 pigs when compared to those of Miller et al. Also, when comparing our isolate STs to those recovered by Miller et al., several STs were identical (ST-854, ST-828, and ST-1107). We previously reported (Thakur et al., 2005a) the clonal diversity of Campylobacter with as high as 65 unique STs recovered from 100 swine isolates. The most common STs we previously reported that were also present in the current study were ST-854, ST-828, and ST-1056. This study supports early findings of the diverse nature of C. coli isolated from swine. In this study, we did not find correlation between persistence and multidrug resistance. Therefore, we do not have any evidence to support or refute that the use of antimicrobials and the selection of multidrug-resistant strains contributes to persistence of C. coli strains.
Conclusion
Despite high genotypic diversity of C. coli on farm, retail meat products were contaminated with isolates of the same STs and PFGE groups as isolates collected from previous stages, indicating persistence of strains from pre- to post-harvest. This finding could have important implications that the control of campylobacteriosis in the food systems (i.e., swine production systems) need to begin on farm.
Footnotes
Acknowledgments
This work was supported by the Food Safety Research and Response Network USDA CSREES Special Research grant 2003–34475–13066 for Food Safety Research and Response Network (FSRRN) and The National Pork Board grant 08-263. We also acknowledge the technical support provided by USDA ARS BEAR Unit. This manuscript made use of the Campylobacter jejuni Multi Locus Sequence Typing website (
Disclosure Statement
No competing financial interests exist.