
Abstract
Several listeriosis outbreaks have been linked to the consumption of fresh or processed produce in recent years. One major determinant of the listeriosis risk is the ability of a food to support growth of Listeria monocytogenes during storage. However, data regarding the ability to support growth of L. monocytogenes are scarce or nonexisting for many produce commodities. Here we synthesize the available data regarding growth behavior of L. monocytogenes on produce, compare the growth data with listeriosis outbreak data, and evaluate the adequacy of the data for predictive modeling. Growth rates and maximum L. monocytogenes population densities differed markedly among produce commodities, and post-harvest processing had a considerable effect on growth dynamics for certain commodities such as tomatoes. However, data scarcity prevented reliable estimation of growth rates for many commodities. Produce outbreaks seemed frequently associated with processed produce and often involved storage under suboptimal conditions (e.g., at room temperature for several hours or for several months in the refrigerator) or environmental cross-contamination after processing. However, no clear associations between high growth rates of L. monocytogenes on fresh produce and outbreaks were detected. In conclusion, produce commodities differ in the supported growth rate of L. monocytogenes, the maximum attainable L. monocytogenes population density, and possibly in the impact of post-harvest processing, but data are currently insufficient to predict growth behavior, and the listeriosis risk appears to be also governed by additional factors.
Introduction
L
L. monocytogenes growth during storage is thought to be an essential contributor to most outbreaks (Codex alimentarius Commission, 1999; FDA/FSIS, 2003; FAO/WHO, 2004). However, food commodities vary in their ability to support growth of L. monocytogenes (FDA/FSIS, 2003). Growth is affected by environmental and food characteristics such as temperature, pH, water activity, sodium chloride concentration, presence of organic acids, and competitive microflora. Several models have been derived that predict growth as a function of time, temperature, and commodity characteristics (see, for example, George et al., 1996; Tienungoon et al., 2000; Carrasco et al., 2006; Valero et al., 2007b; Mejlholm et al., 2010). Such models can be instrumental in determining the optimal formulation, processing, and storage conditions to minimize growth of L. monocytogenes. L. monocytogenes growth models have been validated for numerous food commodities, including deli meats, deli salads, and seafood, but a comprehensive evaluation for produce is so far missing (van Gerwen and Zwietering, 1998; te Giffel and Zwietering, 1999). Predictive microbial models for growth of L. monocytogenes on produce require adequate data for a representative number of commodities and storage conditions. Accurate prediction of L. monocytogenes growth behavior on produce may be complicated by inherent differences in plant microarchitecture, presence of naturally occurring antilisterial substances, and the natural microflora (Nguyen-The and Lund, 1992; Bisha et al., 2010; Noriega et al., 2010). Growth behavior may also be impacted by post-harvest processing such as cutting, dicing, chlorine washes, or packaging under modified atmosphere (MAP) (Beuchat and Brackett, 1990b; Beuchat and Brackett, 1991; Delaquis et al., 2002).
To characterize growth of L. monocytogenes on produce and to evaluate potential public health implications, we reviewed the scientific literature to identify outbreaks of L. monocytogenes associated with produce and summarized the available data regarding L. monocytogenes growth behavior on produce.
Materials and Methods
Data sources
We performed a systematic literature search in the NCBI Pubmed database (
Initially, all available data were extracted from the manuscripts. When data were presented as graphs, maximum growth rates were estimated using log-linear approximation. If multiple graphs were presented in a figure and distinction was uncertain, the highest and lowest maximum growth rates were approximated to capture the range of reported values. Experiments performed at temperatures below 4°C were excluded because of data scarcity and model validity concerns. Moreover, because bacterial inactivation is not studied here, negative growth rates were set to zero. Measures of variability were often not or insufficiently reported and were therefore not considered further.
A second analysis only focused on data presented in tables or ComBase. After commodities that evidently did not support growth (e.g., dried apricots) were excluded, primary growth models were fit for 14 uncooked or cooked commodities (Fig. 1) using DMfit (available at
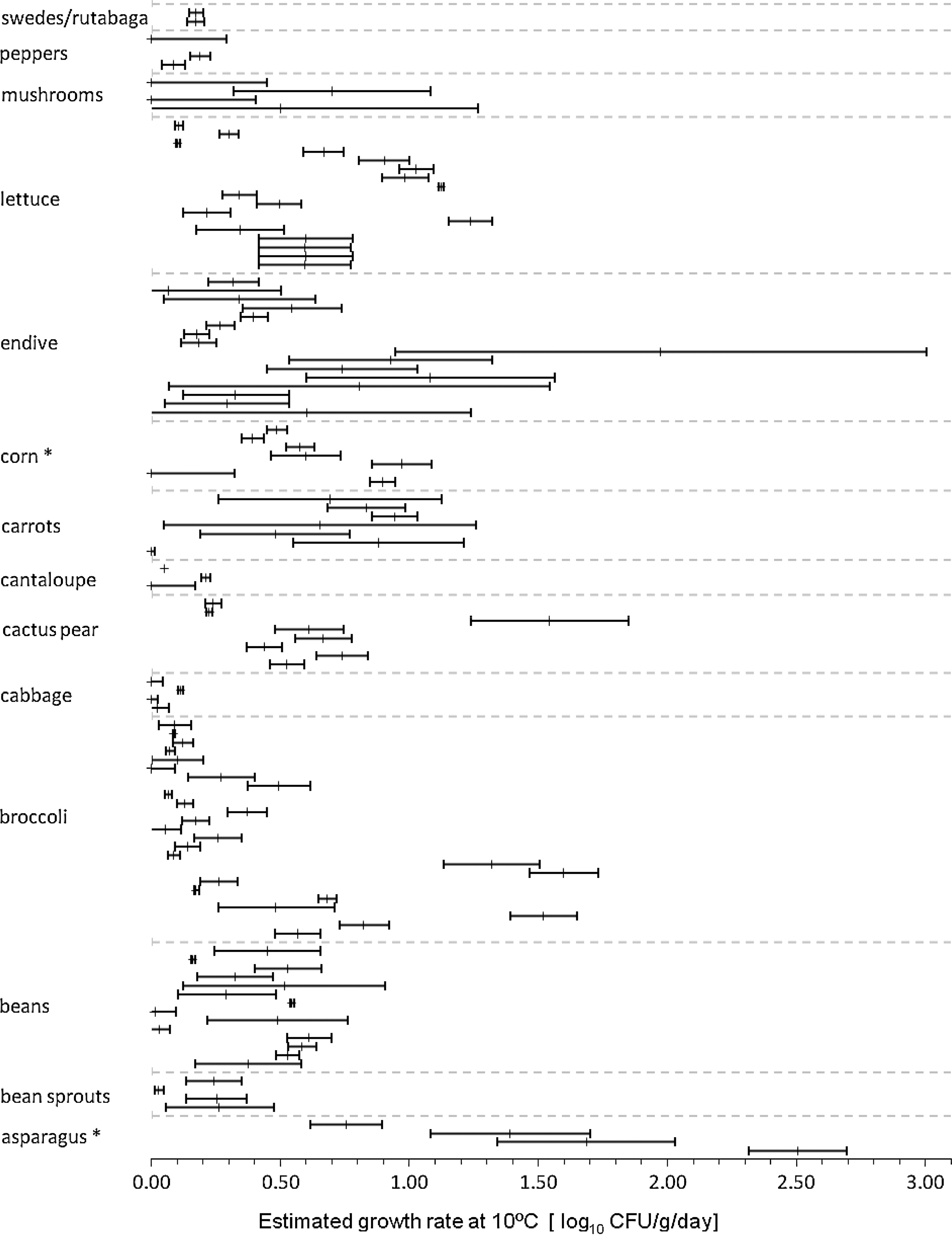
Maximum daily growth rates at 10°C and corresponding standard errors for a variety of produce commodities that support Listeria monocytogenes growth. Values were truncated at zero. Each bar represents one experiment. Commodities heat-treated prior to the growth experiment are indicated by asterisks. All other commodities were used without cooking. CFU, colony-forming units.
Commodity-specific water activity values were generally not available. Water content (USDA, 2011) was evaluated as proxy, but after initial analysis appeared inadequate and is therefore not reported. Produce pH was rarely reported in the growth studies. Values reported by Bridges and Mattice (1939) or Atwater and Woods (1896) served as proxies because no newer, comprehensive surveys of produce pH were available.
Semiquantitative analysis
Data extracted from graphs were only used semiquantitatively as the visual determination of the exponential growth phase may have introduced errors. Consistent with international guidelines (EC 2073/2005 and EC 1441/2007) defining limited growth as a less than 0.5 log10 colony-forming unit (CFU) per gram increase during the normal shelf life, results were categorized as supporting growth if the estimated growth exceeded 0.5 log10 CFU during 10 days of storage at 5°C. This time–temperature combination appeared to be a reasonable approximation of the normal shelf life for many produce commodities (USDA, 2004). For many commodities, growth experiments were performed at multiple temperatures. Growth rates were standardized using the square root model first described by Ratkowsky et al. (1982), using a theoretical minimum temperature of −1.18°C as previously described (FDA/FSIS, 2003). Because growth dynamics differed vastly among studies and replicates, post-harvest processing effects were only evaluated using studies that explicitly compared these treatments, and temperatures that apparently did not support growth were not analyzed further. Lag times depend on pre-experimental conditions (Dufrenne et al., 1997; Crepet et al., 2009) and were therefore not further analyzed.
Quantitative analysis
Maximum growth rates obtained through primary growth models were standardized to 10°C using the Ratkowsky model using a theoretical minimum temperature of −1.18°C (Ratkowsky et al., 1982; FDA/FSIS, 2003). The choice of 10°C was made as this storage temperature appeared reasonable for cold-sensitive (e.g., bell peppers) and cold-resistant (e.g., broccoli, carrots) commodities (USDA, 2004). After exclusion of data that could not be adequately fit, this resulted in a dataset comprising 119 primary growth rate estimates. Of these, a total of 65 observations also allowed comparison of maximum bacterial population densities.
To test whether L. monocytogenes growth rates significantly differ among produce commodities, a general linear model that considered temperature (in °C) as a continuous variable and commodity as an ordinal variable was fit to the primary growth rate data using the general linear model (GLM) procedure available in SAS version 9.2 (SAS Institute, Cary, NC). The Tukey–Kramer adjustment for multiple comparisons was used to correct for multiple comparisons.
Results
L. monocytogenes growth behavior on produce
Commodities differed in the ability to support growth, and results differed among studies and processing forms (Table 1). It is notable that growth of L. monocytogenes on cantaloupe melon rind was not observed, whereas L. innocua has been reported to grow on rind (Ukuku and Fett, 2002; Behrsing et al., 2003). Seven studies compared growth on intact and damaged or cut produce. One study, on tomatoes, reported growth only on intact produce (Beuchat and Brackett, 1991), while four studies, on carrots, cabbage, peppers, and cantaloupe melon, reported growth exclusively on cut, shredded, or damaged produce (Beuchat and Brackett, 1990a; Han et al., 2001; Ukuku and Fett, 2002; Ells and Truelstrup Hansen, 2010). The remaining two studies did not find differences in the ability to support growth of L. monocytogenes, reporting growth on both intact and cut cabbage (Beuchat and Brackett, 1990b) or growth on neither whole nor cut strawberries (Flessa et al., 2005).
Experiments conducted at temperatures below 4°C were excluded.
Based on USDA National Nutrient Database for Standard Reference data for fruits or vegetables, respectively.
Limited growth defined as ≤0.5 log10 colony-forming units growth in 10 days at 5°C.
Includes both whole and cut or shredded commodity.
Summarizes data for different sprout varieties (e.g., alfalfa, mung bean).
ComBase, Combined dataBase for predictive microbiology (available at
Of the studies evaluating the impact of chlorine, four studies, performed on tomatoes, sprouts, and lettuce (two studies), reported no differences in growth rates (Beuchat and Brackett, 1990b, 1991; Lee et al., 2002; Li et al., 2002), whereas two studies, on carrots and green peppers, reported increased growth in chlorine-treated produce (Beuchat and Brackett, 1990a; Han et al., 2001). None of the studies reported decreased growth in chlorine-treated produce. Washing in warm chlorinated water (47°C) favored growth during subsequent storage (Delaquis et al., 2002).
Of the studies evaluating the impact of packaging under passive MAP, four studies, conducted on mushrooms and endive (three studies), reported increased growth rates for L. monocytogenes in MAP compared with atmosphere-packaged produce (Aytac and Gorris, 1994; Bennik et al., 1996; Gonzalez-Fandos et al., 2001) (Ringle et al., 1991, cited by Nguyen-the and Carlin, 1994). Studies on asparagus, broccoli, cactus pear, carrots, cauliflower, sprouts, lettuce, and tomatoes, in contrast, did not detect significant differences in growth rates among MAP and atmosphere-packaged produce (Berrang et al., 1989; Beuchat and Brackett, 1990b, 1991; Aytac and Gorris, 1994; Beuchat and Doyle, 1995; Corbo et al., 2005). None of the studies reported a decreased growth rate in MAP produce.
Growth rates and maximum achievable population densities on fresh or fresh-cut produce
For 17 of the 119 primary data sets, the complete model of Baranyi and Roberts (1994) provided adequate fit according to the goodness of fit test available in the DMfit software. Standardized estimated growth rates varied among and within commodities (Fig. 1). It is notable that estimated growth rates on bean sprouts ranged from 0.03 to 0.3 log10 CFU/g/day, and respective estimates on cabbage and cantaloupe melon ranged from 0 to 0.1 log10 CFU/g/day and 0 to 0.2 log10 CFU/g/day, even though some excluded studies indicated potentially higher growth rates (Leverentz et al., 2003, 2004; Penteado and Leitão, 2004). Maximum achievable population densities varied among commodities and replicates and may be partially temperature-dependent (Fig. 2). On cantaloupe melon, reported maximum population densities only ranged from 3 to 5 log10 CFU/g. Data were scarce for cabbage and coleslaw, but also indicated values of approximately 3–5 log10 CFU/g. Population densities and growth rates varied considerably among studies (Fig. 3).
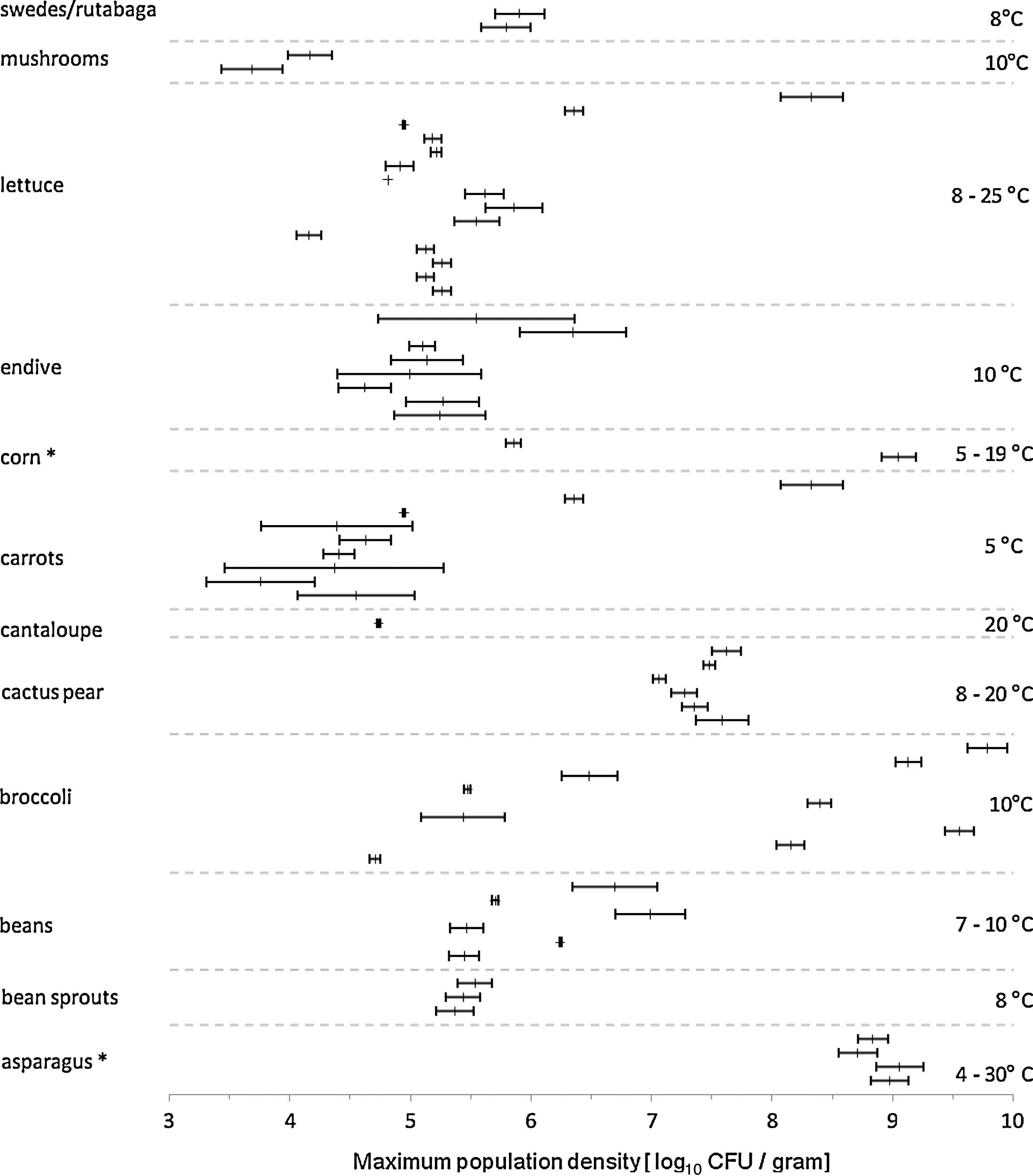
Maximum achievable population density and corresponding standard errors for a variety of produce commodities that support Listeria monocytogenes growth. Each bar represents one experiment. Commodities heat-treated prior to the growth experiment are indicated by asterisks. All other commodities were used without cooking. Temperatures at which growth experiments were performed are indicated on the right. CFU, colony-forming units.

Maximum achievable population density and estimated maximum growth rate (EGR) at 10°C. Data generated in the same study are marked by identical symbols. CFU, colony-forming units.
The commodity effect on growth proved significant in the GLM (p<0.001), confirming significant differences in growth rates among commodities. In particular, growth rates on melons were significantly lower than for the other produce commodities (Table 2).
Based on Tukey–Kramer multiple-testing adjustment; commodities with the same group number are not significantly different from each other (p<0.05).
CFU, colony-forming units; LS, least squares.
Comparison of outbreak and growth data
Review of the available outbreak data revealed that several outbreaks were associated with processed produce (Table 3). Several of the fresh produce commodities implicated in listeriosis outbreaks, such as coleslaw, celery, alfalfa sprouts, and cantaloupe melon, only support modest growth rates and comparably low population densities. However, data were very scarce, and further studies are urgently needed.
Outbreak occurred in a hospitalized or institutionalized population or hospitalization was part of the case definition.
A dash indicates not applicable or information not available.
Discussion
Growth behavior differs among commodities and is affected by processing
Most produce commodities support growth of L. monocytogenes, even though some commodities may be heated before consumption, thereby reducing the public health risk. Cooked or pasteurized produce usually supports rapid growth to high population densities and may represent a risk after post-processing contamination, especially if not refrigerated and consumed without heating. Maximum achievable population densities on fresh produce are highly variable, occasionally reaching values exceeding 109 log10 CFU/g, and may at least be partially temperature dependent.
Processing can affect growth behavior, even though the exact impacts on a commodity remain difficult to predict. Cutting, dicing, or damaging of the produce surface can disseminate pathogens in the produce tissue and generally increases the ability to support growth of L. monocytogenes, except for tomatoes, where the low-pH juice inhibits growth (Beuchat and Brackett, 1990a, 1991; Han et al., 2001; Ukuku and Fett, 2002; Ells and Truelstrup Hansen, 2010). Whether packaging under modified atmosphere increases the ability of L. monocytogenes to grow by inhibiting competing microflora remains to be determined, and the effect may differ by commodity, atmosphere composition, and storage conditions (Berrang et al., 1989; Beuchat and Brackett, 1990b, 1991; Beuchat and Doyle, 1995; Bennik et al., 1996; Gonzalez-Fandos et al., 2001; Corbo et al., 2005; Ringle et al., 1991; cited by Nguyen-The and Carlin, 1994). However, packaging under modified atmosphere evidently does not inhibit growth of L. monocytogenes, and the prolonged shelf life may in itself increase the listeriosis risk. Given these findings, risk assessments should consider commodity and processing form and account for the maximum population density and growth rate.
Data on L. monocytogenes growth behavior is scarce for most produce commodities
For many produce commodities, estimation of growth parameters that adequately consider factors such as storage condition or processing form is currently impossible because of data limitations. Growth parameters for L. monocytogenes on leafy greens have been estimated and used in risk assessments (Crepet et al., 2009; Carrasco et al., 2010; Franz et al., 2010; Danyluk and Schaffner, 2011). Yet, the impact of post-harvest processing remains difficult to estimate, even for leafy greens (Beuchat and Brackett, 1990b; Li et al., 2002). Produce pH, free water activity, and composition and concentration of background microflora are rarely characterized in growth studies, requiring use of external estimates. It is unclear how well such proxies resemble the conditions during the growth experiment—food chemistry can be affected by produce variety, growing season, storage conditions, and processing, and actual values may differ considerably from the proxies (USDA, 2004). Bacterial inactivation dynamics on produce are complex and may be impacted by factors such as sublethal temperatures or background microflora (Ross et al., 2008; Zhang et al., 2010). Differences in inactivation rates therefore may further complicate the analysis. Overall, the available data currently appear insufficient to build adequate predictive growth models for L. monocytogenes on produce, and attempts to fit secondary growth models to the data proved unsuccessful (data not shown [available from the authors on request]).
It is notable that complete primary growth models could only be fitted to a small fraction of datasets, likely reflecting suboptimal study design (e.g., number and spacing of data points, experiment length). Considerations of the behavior of nonlinear models can guide experimental design to optimize the informational value of the collected data. Such theoretically based experimental designs have been derived for several bacterial growth models, including the Ratkowsky model (Bernaerts et al., 2000), the Baranyi model (Grijspeerdt and Vanrolleghem 1999), and the cardinal temperature model with inflection points (Bernaerts et al., 2005). Optimal experimental design can reduce the number of required data points if the growth data follow the assumed growth model. Basing study design on such theoretical considerations and existing data may improve precision of parameter estimates and permit cost-effective estimation of growth parameters.
Many produce commodities associated with outbreaks only permit limited growth
The 1981 coleslaw outbreak represents one of the first described foodborne listeriosis outbreaks (Schlech et al., 1983). Produce-associated L. monocytogenes outbreaks often occurred in commodities that seem to support only limited growth, even though the number of outbreaks is relatively small, and underreporting and limited recognition of produce as a potential source of listeriosis may have biased the set of reported outbreaks. Nonetheless, this finding appears surprising. The ability to support bacterial growth, the growth rate, and the maximum population density are thought to be major drivers of the listeriosis risk because comparably high bacterial doses have to be ingested to cause disease in humans (FDA/FSIS, 2003; FAO/WHO, 2004). It is remarkable, however, that in many documented outbreaks the produce was consumed in multi-ingredient foods and often extensively handled prior to consumption. The growth behavior of L. monocytogenes in the multi-ingredient food may differ from that on the implicated produce, and processing may alter growth behavior further. It is important that in several instances storage under suboptimal conditions (e.g., at room temperature for several hours or for several months in the refrigerator) or environmental contamination contributed to the outbreak (Junttila and Brander, 1989; Salamina et al., 1996; Simpson, 1996; Aureli et al., 2000; Ozfoodnet, 2006). Taken together, these observations may indicate that other factors such as perhaps contamination patterns, extensive handling, and consumption habits contribute to the listeriosis risk associated with produce consumption (Hoelzer et al., 2012).
Conclusions
Produce commodities differ in their ability to support growth of L. monocytogenes and maximum population densities are highly variable. Although the growth behavior of L. monocytogenes on leafy greens has been well studied, data for most other produce commodities are scarce. Only a limited number of studies have evaluated the impact of post-harvest processing, but tentative evidence for produce-specific differences in the impact of such processing exists. It is important that produce-associated outbreaks appear to occur predominantly in processed produce. We did not find evidence to support a paramount importance of growth behavior on the likelihood with which a produce commodity is associated with L. monocytogenes outbreaks. Further research is needed to understand the determinants of growth of L. monocytogenes on produce, to derive reliable predictive models, and to comprehensively assess the determining factors of the listeriosis risk associated with produce consumption.
Footnotes
Acknowledgments
This work was supported in part by appointments to the Research Participation Program at the Center for Food Safety and Applied Nutrition administered by the Oak Ridge Institute for Science and Education through an interagency agreement between the U.S. Department of Energy and the U.S. Food and Drug Administration.
Disclosure Statement
No competing financial interests exist.