
Abstract
Osmoregulated periplasmic glucans (OPGs) are synthesized by the members of the family Enterobacteriaceae when grown under low osmotic growth conditions. Enteropathogens such as Shigella flexneri spend considerable time outside the host environment such as irrigation waters where low nutrient low osmolarity conditions normally may exist. We recently demonstrated that OPGs of S. flexneri are required for optimal growth under low osmolarity low nutrient conditions. Based on homology of the OPG biosynthesis genes to those of Escherichia coli, the presumptive function of opgC and opgB genes is to add succinate and phosphoglycerol residues respectively on OPGs, rendering them anionic. Using lambda-red recombination procedure, we constructed opgB, opgC, and opgBC mutants of S. flexneri. The mutant strain defective in opgC and opgB genes synthesized neutral OPGs. The OPGs without any anionic charges were beneficial for the organism's growth in hypo-osmotic media. However, with the loss of anionic charges from OPGs, mutants were compromised in their ability to combat stress caused by anionic detergents in hypo-osmotic growth conditions. Cloned wild-type genes opgB, opgC, and opgBC, when mobilized to respective opg mutants, simultaneously restored anionic charges to OPGs and tolerance to detergents. The data indicate that anionic charges on the OPGs contribute towards overcoming the stress caused by anionic detergents such as sodium dodecyl sulfate and sodium deoxycholate.
Introduction
D
The high tolerance levels that enteric bacteria exhibit towards anionic detergents appears to be a cooperative multifunctional effect of several cellular functions (Nickerson et al., 1992; Rajagopal et al., 2002). Anionic charges of osmoregulated periplasmic glucans (OPGs) of Escherichia coli have been postulated to contribute toward the electrostatic repulsion of SDS (Rajagopal et al., 2003a). OPGs are a heterogeneous group of branched glucose polymers primarily located in the periplasmic space of Gram-negative bacteria (Kennedy, 1982; Kennedy et al., 1988). OPGs play an important but poorly defined role in symbiotic as well as pathogenic host-microbe interactions (Bohin, 2000; Bohin et al., 2007). Plant, as well as animal pathogenic bacteria defective in the synthesis of OPGs are either avirulent or exhibit suboptimal virulence towards their hosts (Arellano-Reynoso et al., 2005; Bhagwat et al., 1996, 2009). We recently demonstrated that S. flexneri synthesizes anionic OPGs and that disruption of the OPG biosynthesis operon, opgGH, resulted in long lag periods and slower growth under hypo-osmotic growth conditions (Liu et al., 2010). Based on DNA homology among E. coli and S. flexneri, it is expected that anionic characteristics of Shigella OPGs would be the function of opgC and opgB genes, which add succinate and phosphoglycerol residues, respectively, onto OPGs (Bohin and Lacroix, 2007; Lacorix et al., 1999).
Using lambda-red recombination procedure (Liu et al., 2010; Ranallo et al., 2006), we constructed opgB, opgC, and opgBC mutants of S. flexneri. This study offers direct genetic evidence for the role of anionic charges of OPGs from S. flexneri in overcoming tolerance towards anionic detergents such as SDS and sodium deoxycholate (DOC).
Methods
Bacterial strains, growth conditions, and media
S. flexneri 2a wild-type strain 2457T (Kotloff et al., 1995) was routinely grown on LB media and Tryptic Soy Agar plates containing 0.05% Congo Red (w/v). Where appropriate, ampicillin (100 μg mL−1) and chloramphenicol (25 μg mL−1) were added to growth media for selection. Escherichia coli strains carrying plasmids pKD3 and pCP20 (Datsenko et al., 2000) were obtained from the E. coli Genetic Stock Center, Yale University. The plasmid pKM208 was obtained from Kenan Murphy (Murphy et al., 2003). S. flexneri containing plasmids pCP20 or pKM208 were incubated at 30°C as they carry temperature-sensitive replicons.
The growth rates of wild-type and opg mutants were determined in growth media such as Luria broth (LB) and LB with no salts (LBNS) (osmolarity of 407±4 and 85±4 mosmol l−1, respectively)(Bhagwat et al., 2009). SDS sulfate (0–2%) and DOC (0–0.1%) were added to growth media. Osmolarity of growth media was measured with a Wescor vapor pressure osmometer (model 5500, Wescor, Inc., Logan UT). LBNS broth diluted 1:8 in distilled water was used as low nutrient no salt (LNNS) medium (36±3 mosmol l−1) (Bhagwat et al., 2009). Growth was measured (as A600, every 20 min) using a Bioscreen C automatic turbidometric analyzer (GrowthCurves USA, Piscataway, NJ) as described earlier (Bhagwat et al., 2009; Liu et al., 2009). Growth was analyzed at 30°C with continuous shaking. To assess the effect of osmotic stress on growth, media were supplemented with varying amounts of salt (NaCl or KCl) or buffered with HEPES (50 mM, pH 7.1).
Mutagenesis protocol
Mutagenesis was performed using lambda-red protocol as described earlier (Liu et al., 2010; Ranallo et al., 2006). Primers designed based on reference genome sequence of S. flexneri 2a strain 2457T (GenBank accession number AE014073) for generating opgB and opgC mutations have been described (Liu et al., 2010). The S. flexneri opgB mutant was used as a host strain to generate opgC mutation, which resulted in opgBC double mutant strain by standard lambda-red protocol (Ranallo et al., 2006). After initial polymerase chain reaction (PCR) screen, mutations were confirmed by determining DNA sequence of the flanking region of individual genes.
To clone the opgB and opgC genes, the DNA region flanking the 450-bp upstream and 20-bp downstream region at the opgB and opgC loci from the wild-type strain were amplified using a Phusion Fidelity PCR kit (New England Biolabs, Ipswich, MA) following manufacturer's instructions. The amplified DNA fragments were cloned in pGEM-T vector and then in a low-copy pACYC177-based vector (pPQSL_2.0) (Bhagwat et al., 2009) to obtain pLN5. Construct pLL3 carries wild-type opgGH genes cloned in pPQSL_2.0 (Liu et al. 2010). pLN5 and pLL3 were mobilized to opg mutants by electroporation for complementation studies (Bhagwat et al., 2006).
Isolation, purification, and characterization of OPGs
To maximize the yield of OPGs, cells were grown in LBNS medium (85±4 mosmol l−1), and OPGs were extracted using the charcoal adsorption procedure and eluted with aqueous pyridine as previously described (Bhagwat et al., 2009; Lequette et al., 2004). The charged OPGs were further fractionated from total OPGs by ion exchange chromatography using a DEAE-cellulose (Whatman DE53, Sigma Chemicals, St. Louise, MO) column and eluted in a step-wise gradient of 0.05, 0.1, 0.15, 0.2, and 0.5 M NaCl (Liu et al., 2010). The unbound fraction was referred to as neutral OPGs. Succinate content from hydrolyzed OPGs was estimated as described earlier (Bhagwat et al., 2009; Lacorix et al., 1999).
Results and Discussion
OPGs of S. flexneri are required to overcome detergent stress in low osmolarity media
Recently, we demonstrated that OPGs of S. flexneri are needed for optimal growth initiation, especially in low nutrient low osmolarity media (LNNS, osmolarity 36±3 mosmol l−1) (Liu et al., 2010). We set out to examine if OPGs would also play a role in detergent tolerance under low nutrient/low osmolarity conditions. However, under low nutrient conditions, an opgGH mutant was unable to tolerate detergent stress and failed to grow even at 0.1% concentration of SDS (data not shown). Tolerance to SDS is reported to be an energy dependent process (Aspedon et al., 1993; Nickerson and Aspedon, 1992). As a result, poor nutrient conditions in LNNS medium coupled with inability to synthesize OPGs, even at low SDS concentration, proved fetal for opgGH mutant. In full strength LB medium without salts (LBNS, osmolarity 85+4 mosmol−1), the wild-type and opgGH mutant strain had identical growth characteristics; the opgGH strain entered logarithmic growth phase (A600=0.5) at 595±7 min compared to 600±11 min for the wild-type. In the presence of 0.85% SDS, opgGH mutant strain needed significantly longer time (980±22 min) to enter logarithmic growth phase in comparison with the wild-type or opgGH mutant strain complimented with opgGH genes on a plasmid (pLL3) (651±9 and 661±11 min, respectively). These data indicate that the ability to synthesize OPGs is of distinct advantage to overcome SDS stress.
Higher SDS concentrations imposed additional stress on cells entering logarithmic growth phase, and this was true for both the wild-type and opgGH mutant strains (Fig. 1B). However, it took 1100 min for the opgGH mutant to enter log-phase, as compared to 700 and 711 min for the wild-type and opgGH (pLL3) strains, respectively, at 1% SDS in LNNS medium. The opgGH mutant strain failed to grow in LNNS media at SDS concentration > 1.0%. It is noteworthy that under osmolarity conditions where OPG synthesis is repressed (∼400 mosmol l−1) (Bhagwat et al., 2009; Liu et al., 2010), wild-type and opgGH mutant strains were indistinguishable in their ability to begin logarithmic growth phase at SDS concentrations as high as 2% (Fig. 1B, open symbols). The opgGH mutant was impaired in overcoming SDS stress only in LBNS medium (but not in LB medium, osmolarity 407±4 mosmol l−1). The data indicate that, under OPG synthesis permitting conditions (i.e., LBNS medium), the opgGH mutant failed to tolerate detergent stress; this supports the notion that OPG synthesis in low osmolarity media by Shigella might be a beneficial property to overcome SDS stress.
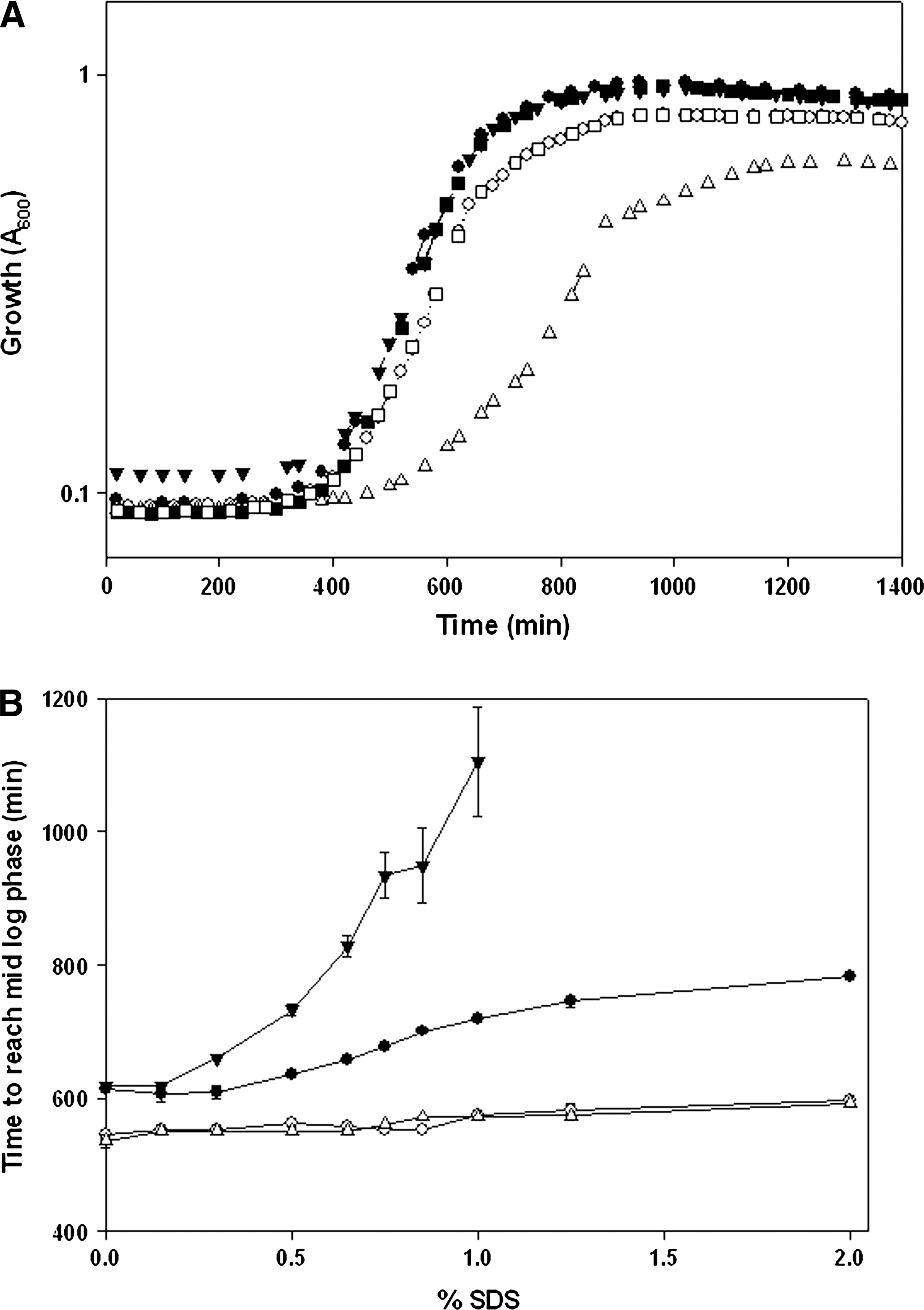
Growth characteristics of Shigella flexneri opgGH to sodium dodecyl sulfate (SDS) in low and high osmolarity media.
Construction of Shigella strain synthesizing neutral OPGs
We hypothesized that anionic charges on OPGs may be acting as a repellent to fend off anionic detergents (such as SDS) entering periplasmic space (Rajagopal et al., 2003a). Based on DNA homology with E. coli, we mutated two genetic loci, opgC and opgB, assumed to be involved in transferring succinate and phosphoglycerate residues, respectively, to OPGs (Bohin and Lacroix, 2007; Lacorix et al., 1999). The effect of opgBC mutations on anionic charges of OPGs was evaluated by estimation of succinic acid residues on OPGs and by DEAE-cellulose chromatography (Fig. 2); due to technical difficulty, phosphoglycerol contents were not examined. The majority of the OPGs synthesized by the opgBC mutant were neutral, as 84% OPGs did not bind to DEAE-cellulose (Fig. 2B). In comparison, 99% of the OPGs of the wild-type strain were retained on DEAE-cellulose; hence, only 1% fraction was neutral (Fig. 2A). Mobilization of wild-type opgBC genes on a low-copy plasmid (pLN5) to the opgBC mutant restored anionic charges to OPGs (Fig. 2C). Succinic acid contents of OPGs from wild-type, opgB, and opgBC (pLN5) strains were 0.69±0.14, 0.61±0.21, and 0.81±0.26 μmol per μmol of OPGs, respectively; no succinic acid was detected in OPGs isolated from opgC and opgBC mutants. These observations confirmed the predicted function of succinylation of OPGs by the opgC gene. Moreover, there appears to be no other gene contributing to anionic substitutions other than opgB and opgC.
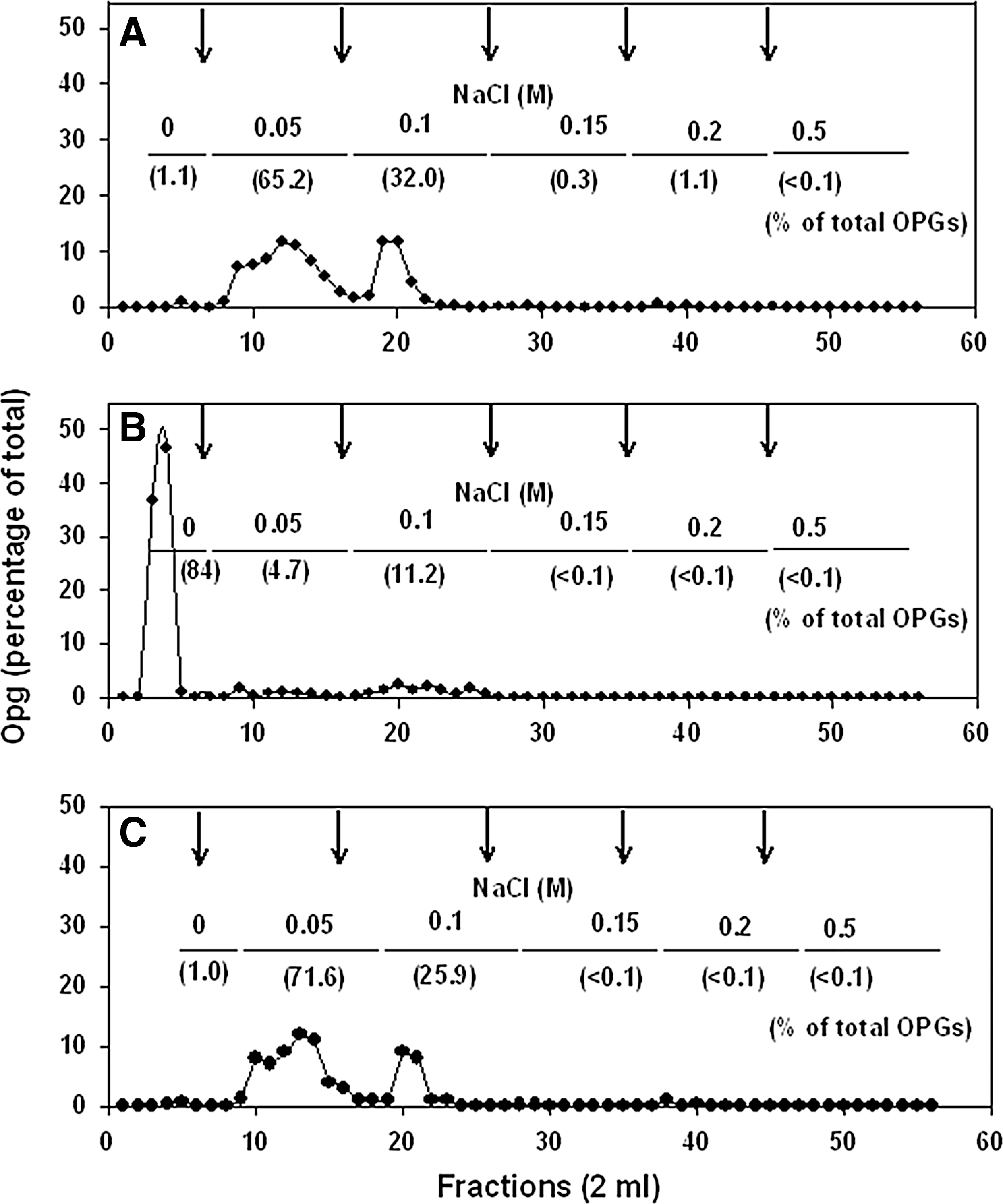
Fractionation of total osmoregulated periplasmic glucans (OPGs) obtained after BioGel P4 chromatography on DEAE-cellulose mini column (1.0 by 1.5 cm). Two milligrams of total OPGs were dissolved in 250 μL of 7% 1-propanol and applied to column. After eluting neutral OPGs in 10 mL of 7% 1-propanol, step gradient of NaCl in 7% 1-propanol was applied in 20-mL volume (0.05, 0.1, 0.15, 0.2, and 0.5 M NaCl, indicated by arrows), and 2-mL fractions were collected and analyzed for total hexose. Horizontal lines indicate pooled fractions; % OPGs eluted at the indicated NaCl concentration are shown below and above each line, respectively.
Anionic substituents of OPGs are needed to overcome SDS stress
Lack of succinic acid residues or phosphoglycerate residues had a significant impact on Salmonella's abilities to tolerate SDS, and contributions due to either anionic residue appeared to be roughly equivalent (Fig. 3, closed and open circles, opgB and opgC strains, respectively). Moreover, the strain that synthesized neutral OPGs (i.e., opgBC) was unable to grow at SDS concentration of > 0.85% (Fig. 3, closed triangles; p<0.001). Like the opgGH mutant, the opgBC mutant was also able to tolerate SDS up to 2% in LB media without significant delay in the onset of logarithmic growth (Fig. 3, open triangles). Mobilization of pLN5 expressing wild-type opgBC to the opgBC mutant restored anionic charges (Fig. 2C) as well as its ability to tolerate SDS at wild-type levels (Fig. 3, closed squares).
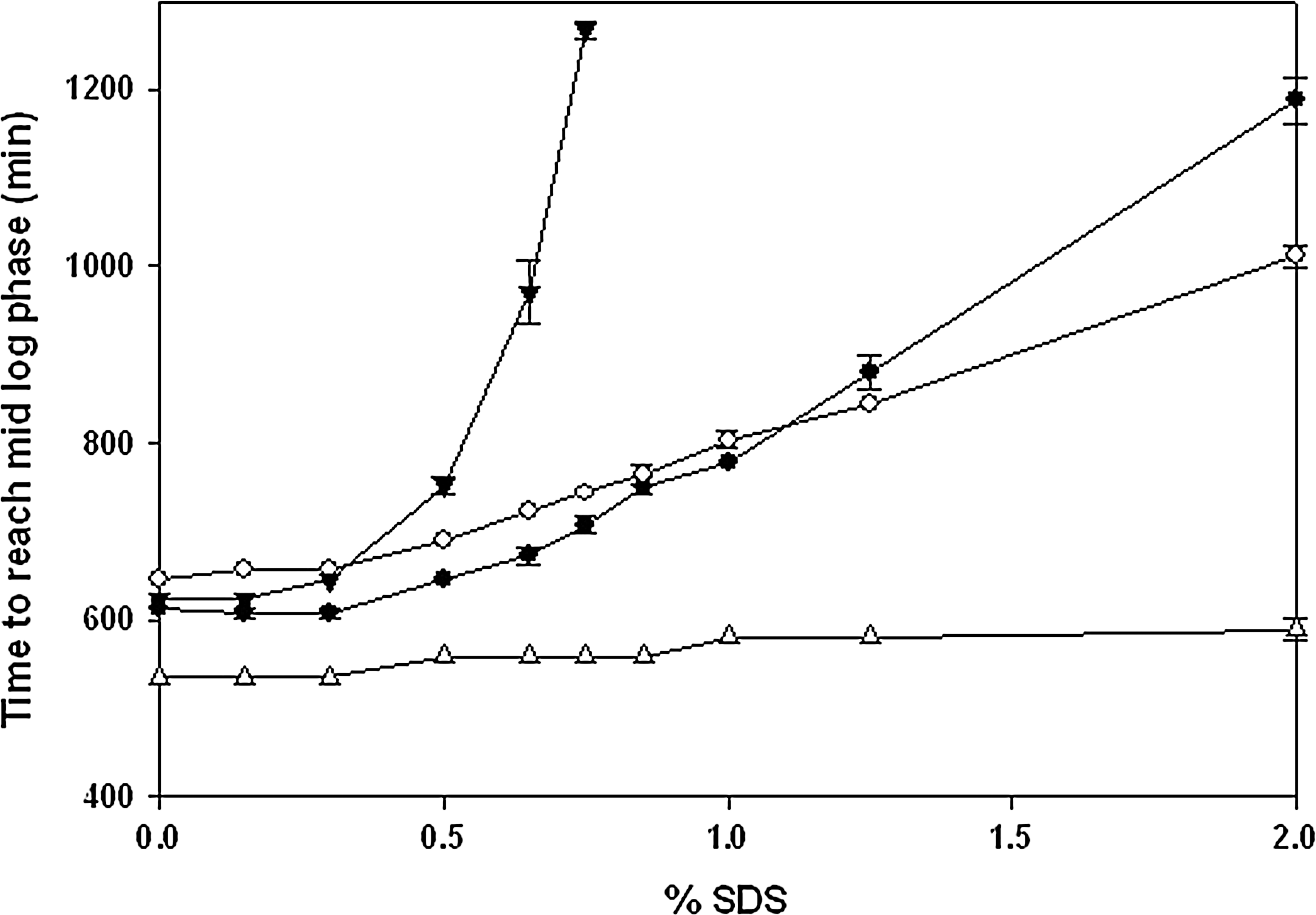
Effect of opgB, opgC, and opgBC mutations on the ability of Shigella to tolerate sodium dodecyl sulfate (SDS). Time to reach mid-exponential growth phase (A600 0.4) was recorded in the presence of varying concentrations of SDS. Cells grown in low osmolarity medium (LBNS), opgB (●), opgC (○), opgBC (▽), and opgBC (pLN5)(■); and cells grown in LB, opgBC (△). Experiments were repeated at least three times, and error bars indicate standard deviation of mean values (not shown when smaller than symbol).
Another anionic detergent stress that is commonly encountered by Shigella in the host gastrointestinal environment is that of DOC (Bopp, 1999; Lampel et al., 2001). A subset of proteins that were identically induced in response to SDS and DOC have been identified in Enterococcus facecalis (Flahaut et al., 1996). We examined how various opg mutants tolerated DOC in low osmotic media (Fig. 4). Like SDS, the presence of DOC exerted significant stress on opgGH and opgBC mutants. Contributions from succinate and phosphoglycerate residues to overcome DOC stress were comparable, while mobilization of opgGH and opgBC genes to respective mutant restored their tolerance to DOC.
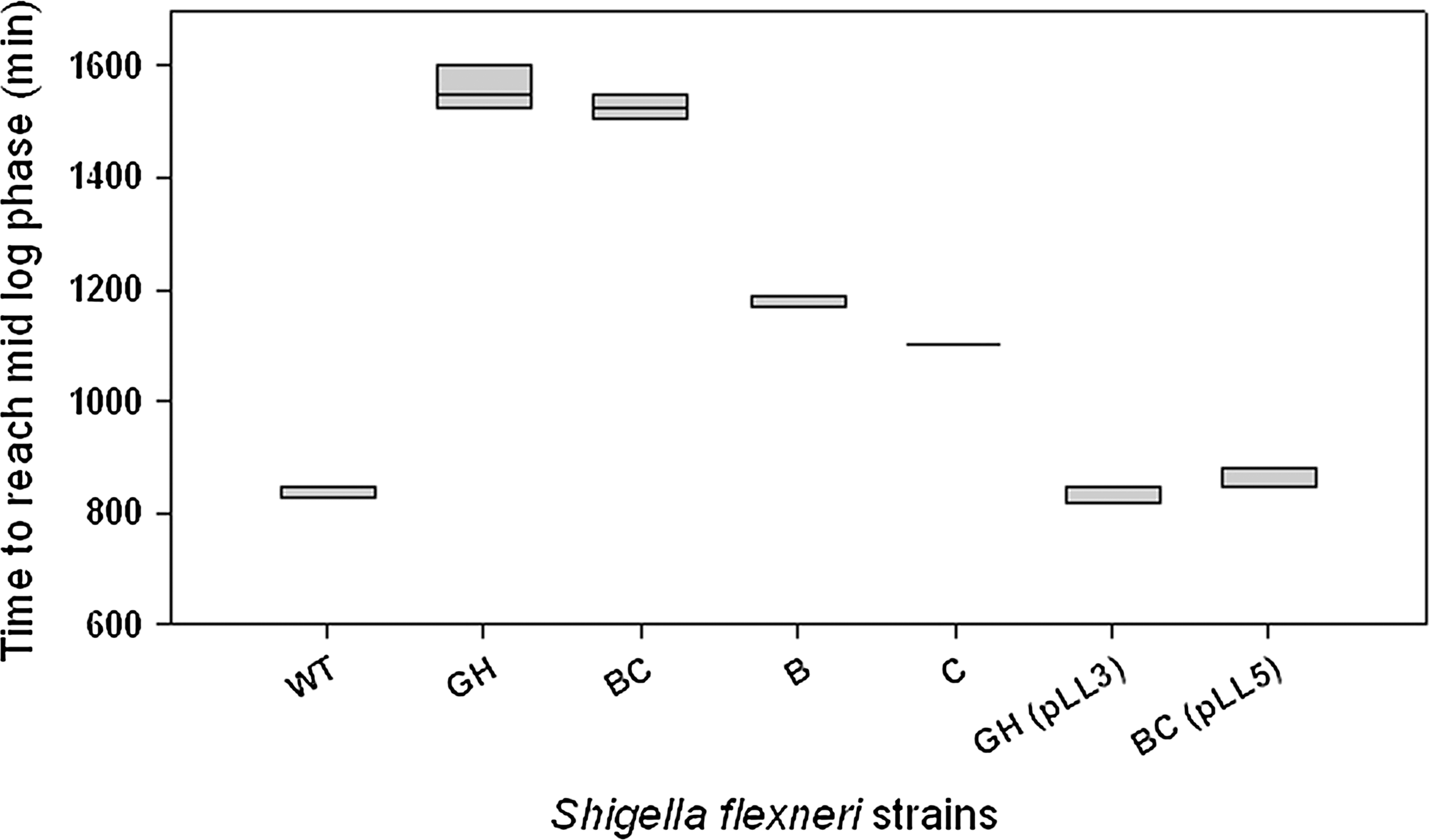
Influence of opg mutations on the ability of Shigella strains to overcome anionic detergent sodium deoxycholate. Cells were grown in low osmolarity medium (LBNS) in presence of 0.1% sodium deoxycholate (DOC), and time to reach mid-log exponential growth phase was recorded. Experiments were repeated at least three times and plotted as a box plot showing 5th and 95th percentile values; line within the box indicates mean (not shown where box size was too small).
While detergents induce membrane damage, bile salts induce both membrane damage and DNA damage. Based on current literature, the interaction between membrane and DNA damage is not fully understood (Merritt et al., 2009). Several gene families contribute towards bile and detergent resistance, such as lipopolysaccharide, efflux pump, porins, and regulatory genes (Gunn, 2000). Mutation in opgBC genes did not result in DOC or SDS sensitivity as severe as has been reported in mutants of the aforementioned gene families. In normal LB media, opg mutants were able to tolerate DOC and SDS, and were indistinguishable from wild-type cultures. However, only under hypo-osmotic growth conditions were anionic OPGs needed to achieve optimal growth, and to overcome SDS and DOC stress. Our results provide the first genetic evidence that anionic charges on OPGs, a structural component of the periplasmic space of S. flexneri, are directly involved in overcoming resistance of anionic detergents such as SDS and DOC.
Footnotes
Disclosure Statement
No competing financial interests exist.