
Abstract
Broiler digestive tract fungal communities have gained far less scrutiny than that given corresponding bacterial communities. Attention given poultry-associated fungi have focused primarily on feed-associated toxin-producers, yeast, and yeast products. The current project focused on the use of pyrosequencing and denaturing gradient gel electrophoresis (DGGE) to identify and monitor broiler digestive fungal communities. Eight different treatments were included. Four controls were an Uninfected-Unmedicated Control, an Unmedicated-Infected Control, the antibiotic bacitracin methylene disalicylate plus the ionophore monensin as Positive Control, and the ionophore monensin alone as a Negative Control. Four treatments were two probiotics (BC-30 and Calsporin) and two specific essential oil blends (Crina Poultry Plus and Crina Poultry AF). All chickens except the Unmedicated-Uninfected Control were given, at 15 days of age, a standard oral Eimeria inoculum of sporulated oocysts. Ileal and cecal digesta were collected at pre–Eimeria infection at 14 days of age and at 7 days post–Eimeria infection at 22 days of age. Extracted cecal DNA was analyzed by pyrosequencing to examine the impact of diet supplements and Eimeria infection on individual constituents in the fungal community, while DGGE was used to compare more qualitative changes in ileal and cecal communities. Pyrosequencing identified three phyla, seven classes, eight orders, 13 families, 17 genera, and 23 fungal species. Ileal and cecal DGGE patterns showed fungal communities were clustered mainly into pre- and post-infection patterns. Post-infection Unmedicated-Uninfected patterns were clustered with pre-infection groups demonstrating a strong effect of Eimeria infection on digestive fungal populations. These combined techniques offered added versatility towards unraveling the effects of enteropathogen infection and performance enhancing feed additives on broiler digestive microflora.
Introduction
F
Due to their ubiquitous nature, fungal hyphae, mycellia, spores, and their byproducts can be ingested by broilers during preening, feeding, and drinking. Beneficial dietary fungi and byproducts potentially offer nutritive value, stimulate host immunity, and exert selective pressure on digestive bacteria resulting in improved production (Buts and De Keyser, 2006; Gao et al., 2008; Willis et al., 2009). Probiotic and prebiotic properties of Saccharomyces spp., Aspergillus spp., and Candida spp. have been reported, and findings have encouraged their inclusion in some food animal and human dietary formulations (Anuradha and Rajeshwari, 2005; Balevi et al., 2009; Biggs and Parsons, 2008; Gao et al., 2008; Lee et al., 2006).
Traditional fungal culturing and isolation techniques can require extended growth periods with highly specific media. The development of molecular techniques has streamlined fungal detection and quantification. Communal DNA or molecular probes can be used to target species as well as to facilitate the monitoring of entire fungal communities (Gharizadeh et al., 2004; Goebes et al., 2007; Griffiths et al., 2006; May et al., 2001; Scupham et al., 2006; Urubschurov et al., 2008). Scupham et al. (2006) estimated that fungi comprise 0–10%, with a median of 2%, of biota associated with murine cecal biofilm, and that fungi sparsely populated digesta outside the ceca. Buts and De Keyser (2006) demonstrated that murine mucosal Saccharomyces boulardii increased levels of digestive enzymes, stimulated immune function, and increased levels of polyamines that stimulate gene expression and protein translation. Urubschurov et al. (2008) suggested diet as the main source of fungi in weaned pigs and the type of feed ingested affects digestive yeast community diversity.
Investigations regarding poultry digestive fungal communities, aside from the toxin-producing varieties, have focused mainly on prebiotic and probiotic applications. Yeast, yeast cell walls, and yeast cell wall–derived mannanoligosaccharides have been exhaustively researched in food animals as anti-enteropathogens and production enhancers. Aspergillus, Candida, and Saccharomyces are routinely given to food animals as probiotics (Anuradha and Rajeshwari, 2005; Balevi et al., 2009). Aside from these beneficial isolates and the vast knowledge base regarding toxigenic fungi, very little is known regarding poultry digestive fungal communities. A major limiting factor has been the difficult and laborious task of conventional culturing. Molecular techniques have been adapted for the use of a single community DNA preparation for differentiation of general and specific bacterial, protozoan, plant, and fungal genetic targets. Sample DNA assayed in the current study was collected during a related study in which we examined the impact of Eimeria infection and treatment with two commercial probiotics and two essential oil blends (EO) to minimize this impact on ileal and cecal bacterial populations (Hume et al., 2011). Mixed Eimeria infection was selected as a model to study intestinal modifications caused by a common pathogen that can cause profound changes in intestinal microbial communities (Hume et al., 2006, 2011; Oviedo-Rondón et al., 2006). Coccidiosis is one of the most endemic enteric diseases in broiler production (Dalloul and Lillehoj, 2005). Pyrosequencing and denaturing gradient gel electrophoresis (DGGE), body weight gain (BWG), and feed conversion (FC) values were employed to examine these effects. In the current study, pyrosequencing and DGGE were applied to community DNA preparations to profile poultry ileal and cecal fungi. As far as the authors could determine, this is the first molecular attempt to elucidate broiler digestive tract fungal populations. The objectives were to identify digestive fungal populations and examine the effects of dietary probiotics and EO, along with a mixed Eimeria infection, on digestive fungal communities. Pyrosequencing was applied to examine the impact of feed additives and Eimeria infection on individual constituents in the fungal community, while DGGE was used for comparison of more gross changes in community constituents.
Methods
Broiler husbandry
A total of 384 1-day-old male Ross 708 chickens were randomly placed in 48 floor pens (8/pen) as previously described (Hume et al., 2011). Broilers were reared according to safe practices authorized by the Animal Use Protocol Committee of North Carolina State University.
Diets and treatments
Eight treatments (Table 1) (Hume et al., 2011) included four controls: Uninfected-Unmedicated (UU), Unmedicated-Infected (UI), the antibiotic bacitracin methylene disalicylate (BMD; Alpharma Inc., Animal Health, Bridgewater, NJ) at 50 g/ton plus the ionophore monensin at 90 g/ton (Coban; Elanco Animal Health Division of Eli Lilly & Co., Indianapolis, IN) as Positive Control (PC), and Coban as Negative Control (NC). Four treatments included the following feed additives: two probiotics, BC-30 (BC30; Ganeden Biotech, Mayfield Heights, OH) and Calsporin (CAL; Quality Technology International, Inc., Elgin, IL); and two specific EO, Crina Poultry Plus (CPP; DSM Nutritional Products, Parsippany, NJ) at 300 ppm and Crina Poultry AF (CPF; DSM Nutritional Products) at 100 ppm. The four treatment diets also contained monensin. Birds were fed a balanced diet (NRC, 1994) and water ad libitum (Table 2) (Hume et al., 2011).
Hume et al., 2011.
All treatments and diets contained phytase enzyme (Ronozyme P CT; DSM Nutritional Products, Parsippany, NJ) at 185 g/ton to provide 925 FYT (phytase units) and ionophore monensin at 90 g/ton (Coban-90; Elanco Animal Health, Greenfield, IN) in the starter and grower diets (1–35 days of age) of all treatments. Each treatment was assigned to six pens with six male Ross 708 chickens each.
Bacitracin methylene disalicylate at 50 g/ton (Alpharma Inc., Animal Health, Bridgewater, NJ).
BC-30 is a probiotic containing Bacillus coagulans GBI-30, 6086 at 2.3×107 cfu/g of feed (Ganeden Biotech, Mayfield Heights, OH).
Calsporin contained B. subtilis C-3102 at 3.6×107 cfu/g of feed (Quality Technology International Inc., Elgin, IL).
Crina Poultry Plus and Crina Poultry AF are two specific essential oil blends (DSM Nutritional Products, Parsippany, NJ).
Treatment: UU, Unmedicated-Uninfected Control; UI, Unmedicated-Infected Control; PC, Positive Control; NC, Negative Control; BC30, BC-30 Probiotic; CAL, Calsporin Probiotic; CPP, Crina Poultry Plus 300 ppm; CPF, Crina Poultry AF 100 ppm.
Cl. Sphaerospermum, Cladosporium sphaerospermum; H. pseudokoningii, Hypocrea pseudokoningii; Mi. oncoperae, Microhilum oncoperae; My. Cinctum, Myrothecium cinctum; Penicillium adametzioides, Penicillium adametzioides; V. incurvum, Verticillium incurvum.
nd, Not detected.
Coccidia infection
All chickens except those in the UU control group were gavaged at 15 days of age to introduce a standard oral Eimeria inoculum (0.5 mL/bird) of sporulated oocysts of mixed coccidia species: E. acervulina (2×105/mL), E. maxima (2.5×104/mL), and E. tenella (5×104/mL). The inoculum was provided by Dr. Mark C. Jenkins (Animal Parasitic Diseases Laboratory, ARS, USDA). Oocytes had been determined to be non-resistant to any ionophore. The Eimeria doses selected were observed in previous experiments to cause mild intestinal lesions similar to the ones observed under commercial conditions of broiler production.
Sampling and DNA extraction
Broilers were sacrificed by cervical dislocation. Ileal and cecal digesta were collected aseptically by excision, and by voiding of contents into sterile sample tubes, from six broilers at pre–Eimeria infection per treatment group at 14 days of age and from six additional broilers post–Eimeria infection per group at 22 days of age. Samples were lyophilized and stored at −80°C until DNA extraction from 0.25 g each (Hume et al., 2003, 2011).
Pyrosequencing
Tag-encoded FLX amplicon pyrosequencing (bTEFAP) was performed as described previously using fungal SSU sequencing and primers SSUFungiF 5’TGGAGGGCAAGTCTGGTG’3 and SSUFungiR 5’TCGGCATAGTTTATGGTTAAG’3 (Research and Testing Laboratory, Lubbock, TX) (Dowd et al., 2011). Initial generation of the sequencing library utilized a one-step polymerase chain reaction (PCR) with a total of 30 cycles and a mixture of Hot Start and HotStar high-fidelity taq polymerases with average read length of 400 bp. Tag-encoded FLX amplicon pyrosequencing analyses utilized Roche 454 FLX instrument with Titanium reagents; titanium procedures were performed at the Research and Testing Laboratory based upon RTL protocols (
Fungi diversity data analysis
Following sequencing, all failed sequence reads, low-quality sequence ends and tags and primers were removed, and sequence collections were depleted of non-fungal ribosome sequences and chimeras using B2C2 (Gontcharova, 2010), as described previously (Bailey et al., 2010). To identify fungi in the remaining sequences, sequences were denoised, assembled into clusters, and queried using a distributed BLASTn .NEt algorithm (Dowd et al., 2005) against a database of high-quality 28s fungal sequences derived and curated monthly from National Center for Biotechnology Information (NCBI). Database sequences were characterized as high quality based upon similar criteria utilized by RDP ver 9 (Cole et al., 2009). Using a .NET and C# analysis pipeline, the resulting BLASTn outputs were compiled and were validated using taxonomic distance methods, and data reduction analysis was performed as described previously (Bailey et al., 2010).
Fungal identification
Based upon the above BLASTn-derived sequence identity (percent of total length query sequence which aligns with a given database sequence) and validated using taxonomic distance methods, the fungi were classified at the appropriate taxonomic levels based upon the following criteria. Sequences with identity scores, to known or well-characterized 28S sequences, greater than 97% identity (<3% divergence) were resolved at the species level, between 95% and 97% at the genus level, between 90% and 95% at the family level, between 85% and 90% at the order level, between 80% and 85% at the class level, and between 77% and 80% at the phyla level. After resolving based upon these parameters, the percentage of each fungal ID was individually analyzed for each sample providing relative abundance information within and among the individual samples based upon relative numbers of reads within each. Evaluations presented at each taxonomic level, including percentage compilations, represent all sequences resolved to their primary identification or their closest relative (Bailey et al., 2010).
Denaturing gradient gel electrophoresis
Amplification targeted a highly variable region of fungal 28S ribosomal DNA and was run according to Sandhu et al. (1995). Template DNA was added to Ex Taq Hot Start Mix (TaKaRa Bio USA, Mountain View, CA, a division of Clontech Laboratories, Inc., Madison, WI; vendor Fisher Scientific, Pittsburg, PA) containing 20 pmol of each universal fungal primers U1 (5’-GTGAAATTGTTGA AAGGGAA-3’) and U2 (5’-
Results
Pyrosequencing
Three phyla were identified, with a total of 392 hits, seven classes, eight orders, 13 families, 17 genera, and 23 species. Ascomycota and Basidiomycota were detected in Eimeria pre- and post-infected broilers, while Microsporidia was detected only in post-infected broilers. Twenty of the 23 total species were detected in pre-infected broilers, with the distribution of detected sequences being limited to three treatment groups: UU, CPP, and CPF. Seventeen of the 20 species detected in pre-infected broilers came from the group given CPP (Table 2). A Cladosporium sp. and Cladosporium sphaerospermum were detected in UI control broilers, and Candida khmerensis was found in CPF. Those species at greater than 10% of the detected population were Alternaria japonica, Alternaria sp., Candida kmerensis, Cladosporium sp., Cladosporium sphaerospermum, and Embellisia sp. Only eight species sequences were detected in ceca from post–Eimeria infected broilers (Table 3). Again, detected and identified sequences were seen in three treatment groups: UI control, CAL, and CPP. Seven species sequences were detected at levels 10% and above: Alternaria japonica, and Alternaria sp., Embellisia sp., Engyodontium album, Malassezia restricia, Systenostrema alba, and Trema sp. Sequences given the identity of known genus and the designation of species refers to a unique species sequence that was identified, but did not match a known fungal species sequence in the data base. A few of the sequences identified have plant, insect, or undomesticated animal disease potential, but are likely filling vastly different niches in the broiler digestive tract.
Treatment: UU, Unmedicated-Uninfected Control; UI, Unmedicated-Infected Control; PC, Positive Control; NC, Negative Control; BC30, BC-30 Probiotic; CAL, Calsporin Probiotic; CPP, Crina Poultry Plus 300 ppm; CPF, Crina Poultry AF 100 ppm.
E. album, Engyodontium album.
nd, Not detected.
Denaturing gradient gel electrophoresis
Analysis of PCR amplicons on agarose gel revealed positive amplification from only seven DNA samples: ileal UI post–Eimeria infection, and cecal post-infection UI, PC, CAL, BC30, CPP, and CPF. All other amplicon mixtures (10 μL of each) were subjected to a second PCR to enhance visualization of the sparse results from the primary amplification. The five samples that gave positive results with the first PCR were similarly examined by a second reaction. This second amplification was to determine if any qualitative changes might occur following the first PCR amplification. Resulting band patterns from those reactions were similar to those from the first amplicon patterns.
Ileal Eimeria pre- and post-infection fungal amplicons (Fig. 1) were clustered separately with a 34.6% SC. Pre-infection patterns, with the exception of CPF, were clustered together with a 77.3% SC. The pattern for pre-infection CPF was most unlike (18.2% SC) any pre- and post-infection pattern. Patterns from post-infection ilea shared a 74.1% SC. The EO groups at pre- and post-infection clustered together with moderate to relatively high %SC. Clustering of PC with BC30 was seen at pre- and post-infection with greater pattern similarity seen at post-infection.

Dendrogram of 28S rDNA denaturing gradient gel electrophoresis (DGGE) amplicon patterns of ileal fungal contents in broilers at 14 days of age Eimeria spp. pre-infection (Il Pr) and at 22 days of age, 7 days after infection (Il Po). Percentage similarity coefficient (bar) of band patterns is indicated by their grouping on the dendogram. Treatment: UU, Unmedicated-Uninfected Control; UI, Unmedicated-Infected Control; PC, Positive Control; NC, Negative Control; BC30, BC-30 Probiotic; CAL, Calsporin Probiotic; CPP, Crina Poultry Plus; CPF, Crina Poultry AF.
Similar to arrangements seen in ileal and cecal MC (Fig. 2) were clustered mainly into pre- and post-infection patterns. Pre-infection UU and UI were clustered together and shared a 93% SC, while UI shared a 95.8% SC with the two groups given EO. The post-infection UU was clustered with the pre-infection groups, but with a very low 43.4% SC.
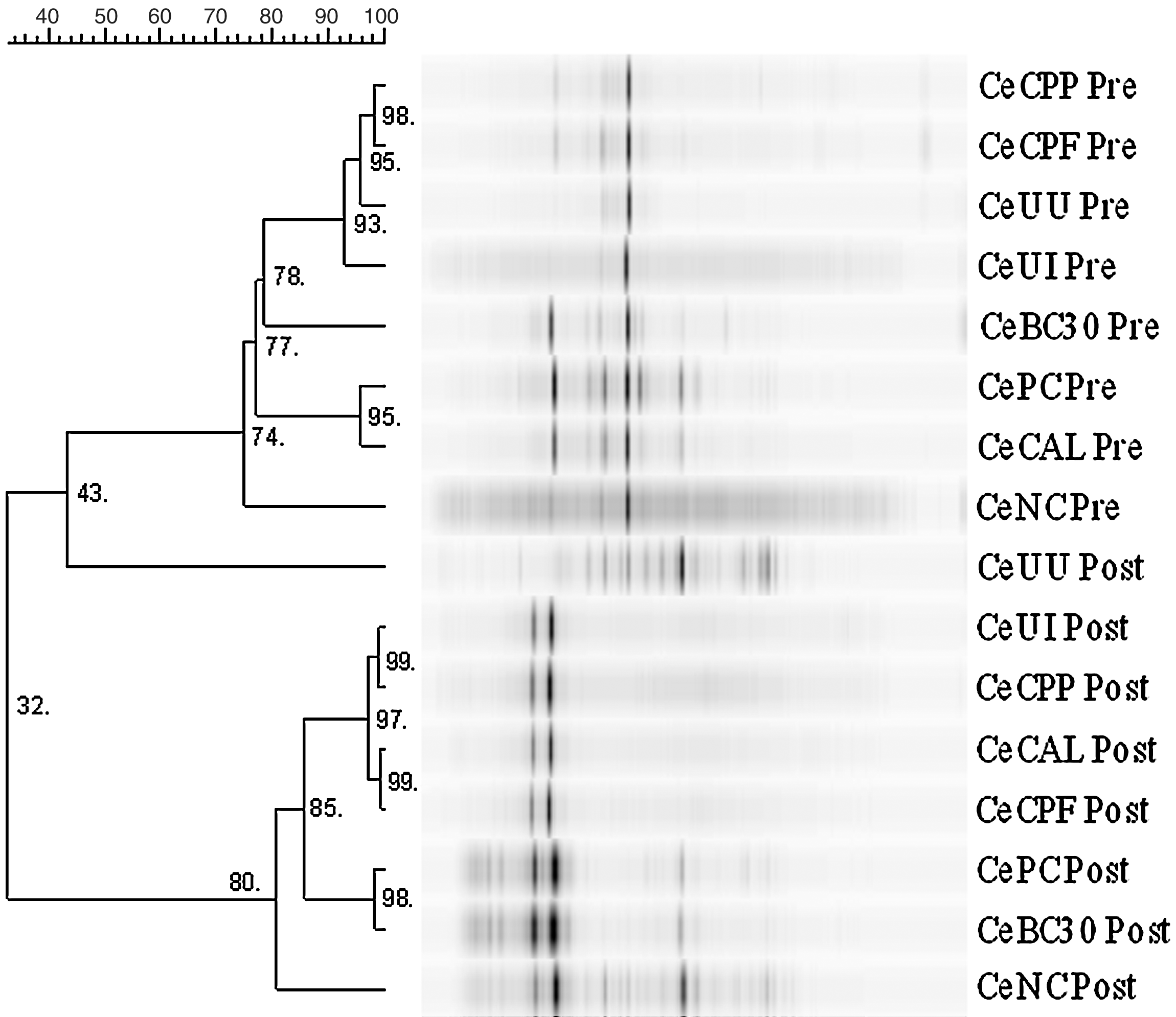
Dendrogram of 28S rDNA denaturing gradient gel electrophoresis (DGGE) amplicon patterns of cecal fungal contents in broilers at 14 days of age Eimeria spp. pre-infection (Il Pr) and at 22 days of age, 7 days after infection (Il Po). Percentage similarity coefficient (bar) of band patterns is indicated by their grouping on the dendogram. Treatment: UU, Unmedicated-Uninfected Control; UI, Unmedicated-Infected Control; PC, Positive Control; NC, Negative Control; BC30, BC-30 Probiotic; CAL, Calsporin Probiotic; CPP, Crina Poultry Plus; CPF, Crina Poultry AF.
Discussion
In the current study, relative abundance of broiler ileal and cecal fungi were examined following treatment with two commercial probiotics and two EO as feed additives and Eimeria infection. This current study is a companion to the study in which we examined the effects of treatment and infection on bacterial populations, BWG, and FC (Hume et al., 2011). Twenty-three species sequences were detected in pre–Eimeria-infected chicks at 14 days of age, while only eight were detected 7 days later at post-infection. This decline in species may be related to the maturing of the cecal microflora towards a complement of bacteria more antagonistic to fungi than those found in the younger chicks. Eimeria infections could also reduce microbial diversity in broiler cecal contents (Hume et al., 2011). Additionally, one must always consider potential PCR bias that may be introduced by combinations of template, possible PCR inhibitors co-isolated with template DNA, and reaction mixture components (Wintzingerode et al., 1997; Poltz et al., 1998; Anderson et al., 2003).
The majority of detected fungal species were seen in broilers given Eimeria and no feed additives, and mainly in chickens fed the same basal diet, but with CPP. Components of CPP (i.e., benzoic acid, thymol, eugenol) exhibit in vitro and dose-dependent antibacterial and antifungal properties (Kandil et al., 1994; Vázquez et al., 2001; Cueva et al., 2010). These antibiotic effects of CPP may be suppressing the growth of antagonists that would normally outcompete the detected fungal species. The final effect for the broiler host seems to be positive, since CPP broilers had significantly better feed utilization than UI control broilers up to 14 days of age (Hume et al., 2011). The CPF formulation is similar to CPP, with the exception that CPF does not contain benzoic acid and relatively less CPF (100 vs. 300 ppm) was added to the feed. Groups UU and UI were not feed additives. However, the two Cladosporium spp. were observed in UI at pre–Eimeria infection, but not in UU, when they were essentially the same treatment up to 14 days. The two Malassezia spp., Systenostrema alba, and the Trema sp. detected at post-infection were not evident in UU. These differences may be directly attributed to effects of mixed Eimeria infection. The mixed Eimeria spp. infection used caused mild lesions in the jejunal, ileal, and cecal mucosa, and almost no lesions were observed in the duodenum. Mucosal lesions indicated changes in intestinal physiology that can be linked to reduction in diversity and number of fungal populations as they were observed in bacterial populations in other studies (Hume et al., 2006, 2011; Oviedo-Rondón et al., 2006).
Broilers fed dietary BC30 and the two EO treatments had BWG better than broilers in the UI treatment, similar to the PC and NC, but lower than the UU control group (Hume et al., 2011). Feed utilization of the two probiotic treatments and CPF group was similar to UU, control groups PC and NC, and better than UI. Higher numbers of fungal populations did not indicate better feed utilization post-infection. Identifying the treatment attributes eliciting these responses and their implications will require further study.
Pyrosequencing was very sensitive at detecting shifts in individual sequences, whereas DGGE was able to detect qualitative shifts in entire populations. Scupham et al. (2006) estimated the fungal community as a fraction of the total murine cecal biofilm microflora. It was apparent that fungal DNA was present in these broiler digesta. However, target sequence quantity was very low. A subsequent PCR of 10 μL of amplicon mixture from the initial amplification was used for samples that did not yield a positive result in the initial PCR. This second PCR gave positive bands on agarose gels for all apparently negative samples and distinct bands following DGGE. Re-amplification can result in PCR artifacts, heteroduplex formation, or bias from low template copy number (Chandler et al., 1997; Thompson et al., 2002; Syvyk et al., 2008). However, the DGGE band patterns were very similar in profile to those seen in the previously positive samples. Additionally, re-amplification of cecal samples that gave positive results in the first PCR resulted in band patterns identical to the first assays.
Ileal and cecal DGGE patterns, respectively, formed two main clusters: pre– and post–Eimeria infection. The fungal cluster pattern somewhat resembles the effect of Eimeria infection exhibited by bacterial clustering (Hume et al., 2006, 2011; Oviedo-Rondón et al., 2006). The effect of Eimeria on cecal fungal communities can be seen with the clustering of post-infection UU with the pre-infection patterns. Common to these groups is that they were not infected with Eimeria. Ileal microbial communities are more transient than cecal fermentative communities. Therefore, far fewer fungi were found in broiler ileal contents than in ceca. Also, due to the expected ileal transience, there would likely be less of an effect of Eimeria infection on ileal than on cecal communities.
The combined techniques of pyrosequencing and DGGE offered added versatility towards unraveling the effects of enteropathogen infection and performance enhancing feed additives on broiler digestive fungal flora. Results show modifications of fungal communities resulting from feed additives prior to infection with a standard mixed Eimeria spp. Nevertheless, none of the feed additives evaluated resulted in modifying the digestive fungal populations to resemble those seen in uninfected broilers post-infection. This result offers preliminary insights into broiler digestive fungal communities, and the effects of pathogen infection and performance enhancing diet supplements on these communities.
Footnotes
Acknowledgments
Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture or North Carolina State University.
Disclosure Statement
No competing financial interests exist.