
Abstract
“Alheira” is a traditional smoked meat sausage produced in the north of Portugal, representing an important economic resource for the region. This meat product has been subjected to research studies with the aim of detecting the presence of common foodborne pathogens, but, to our knowledge, isolation of emerging foodborne Aeromonas from alheira has never been previously described. Present work attempts to evaluate the Aeromonas species diversity of 84 isolates of Aeromonas spp. collected from 32 alheira samples. All presumptive Aeromonas isolates were subjected to genotyping by enterobacterial repetitive intergenic consensus–polymerase chain reaction analysis. The isolates presenting a different pattern were subjected to gyrB gene sequencing for species classification, and the species A. hydrophila, A. salmonicida, A. caviae, A. media, and A. allosaccharophila were identified. The Aeromonas species diversity found has not been previously described in any other meat product evaluated in previous studies. It is also important to highlight the presence of A. hydrophila and A. caviae because they were previously associated with illness in humans, including gastroenteritis.
Introduction
Several studies investigated the microbiological content of alheira. This product presented high microbial counts, and lactic acid bacteria represented the dominant microflora (Esteves et al., 2008; Ferreira et al., 2006). In addition, several foodborne pathogens such as Staphylococcus aureus, Salmonella spp., Clostridium perfringens, Listeria monocytogenes, Bacillus cereus, and Yersinia enterocolitica have been isolated from this traditional sausage (Esteves et al., 2007a,b, 2008; Ferreira et al., 2006).
Over the past 25 years, mesophilic Aeromonas have received increasing attention as an emergent agent of foodborne disease. In Spain, Aeromonas spp. occupied fourth place among the list of causes of gastrointestinal disease reported each year over the period 1997–2006 (Pablos et al., 2009). Some Aeromonas species are responsible for producing intestinal and extraintestinal infections worldwide, and gastroenteritis represents the most common clinical presentation associated with Aeromonas, followed by localized soft-tissue infections and bacteremia (Abbott et al., 2003; Janda and Abbott, 2010; Schlenker and Surawicz, 2009; Vila et al., 2003). Bacteremia mainly occurs in patients with underlying diseases such as hepatobiliary disorders, cancer, and diabetes (Figueras, 2005).
The genus Aeromonas belongs to the family Aeromonadaceae and currently comprises 26 species: A. allosaccharophila, A. aquariorum, A. bestiarum, A. bivalvium A. caviae, A. encheleia, A. eucrenophila, A. fluvialis, A. hydrophila, A. jandaei, A. media, A. molluscorum, A. piscicola, A. popoffii, A. rivuli, A. salmonicida, A. sanarellii, A. schubertii, A. simiae, A. sobria, A. taiwanensis, A. tecta, A. trota, A. veronii, and also Aeromonas sp. HG11 and Aeromonas sp. HG13 (Enteric Group 501) (Alperi et al., 2010a,b; Beaz-Hidalgo et al., 2009; Demarta et al., 2008; Figueras et al., 2011; Harf-Monteil et al., 2004; Martin-Carnahan and Joseph, 2005; Martínez-Murcia et al., 2008; Miñana-Galbis et al., 2004, 2007). Recently, the name A. diversa was proposed to designate Aeromonas sp. HG13 (Miñana-Galbis et al., 2010). Martínez-Murcia et al. (2011) report that two additional potential new species “A. cavernicola” and “A. lusitana” have been recently recognized and are in the process of being described. This genus consists of facultative anaerobic Gram-negative non–spore-forming motile bacilli or coccobacilli, predominantly motile by a single polar flagellum, which are oxidase and catalase positive, reduce nitrate to nitrite, and are resistant to the vibriostatic agent O/129 (2,4-diamino-6,7-diisopropylpteridina) (Martin-Carnahan and Joseph, 2005). Aeromonas are ubiquitous in nature, being considered autochthonous inhabitants in aquatic environments, and have also been isolated from a wide range of foods, including meat, poultry, milk and milk products, fish and shellfish, and vegetables (Fiorentini et al., 1998; Hazen et al., 1978; Neyts et al., 2000; Pablos et al., 2009). Furthermore, the isolation of Aeromonas spp. from meat products, including sausages, has also been reported (Hudson and De Lacy, 1991; Singh, 1997; Villari et al., 2000). Some Aeromonas spp. are facultative psychrophiles and maintain the ability to express virulence factors at refrigeration temperatures (Kirov et al., 1993; Mano et al., 2000). This fact, along with the ubiquitous nature of Aeromonas spp. and its presence in the food chain, suggested that food is a vehicle for illness caused by Aeromonas spp. (Isonhood and Drake, 2002).
Housekeeping gene sequencing analysis is considered as a high-resolution approach to ascertain phylogenetic relationships in bacteria (Stackebrandt et al., 2002). During the past few years, the sequencing of gyrB gene (encoding the B-subunit of DNA gyrase, a type II DNA topoisomerase) has proved to be a useful marker for Aeromonas species identification and strain differentiation, even when using only a partial gyrB gene sequence (Fontes et al., 2010, 2011; Martinez-Murcia et al., 2008; Saavedra et al., 2006, 2007; Soler et al., 2004; Yañez et al., 2003).
The present study was undertaken with two aims: to isolate and to describe the phylogenetic diversity of Aeromonas species in traditional Portuguese sausage alheira by using the partial gyrB gene sequencing.
Methods
Sample collection and isolation of Aeromonas species
A total of 32 samples of alheira were purchased from local retail establishments, in January and February 2007. They were immediately transported to the laboratory and analyzed within 2 h.
Twenty-five grams of each alheira sample were weighed aseptically, homogenized for at least 2 min in stomacher bags containing 225 mL of alkaline peptone water, and incubated for 24 h at 30°C. The homogenate was streaked on glutamate starch phenol-red agar (GSP; Merck, Whitehouse Station, NJ) plates. After incubation at 30°C for 24 h, yellow colonies were isolated and subcultured into tryptone soya agar (TSA; Oxoid, Hampshire, UK). Oxidase-positive colonies were taken as presumptive Aeromonas isolates. Strains were maintained on TSA (Oxoid) plates at 4°C and were stored in 10% glycerol at −80°C. Culture collection strains used in this study were purchased from the Spanish Collection of Type Cultures (CECT).
DNA extraction
A single colony from fresh cultures was resuspended in 100 μL of TE buffer (Tris-HCl 10 mM, pH 8; and EDTA 1 mM, pH 8), and 200 μL of Chelex 100 (BioRad) was added. The tube was vortexed at high speed for 1 min, incubated at 96°C for 10 min, kept at −20°C for 10 min, and this process was repeated three consecutive times. The tube was again vortexed and centrifuged for 5 min at 12,000×g. The supernatant was transferred to a fresh tube and stored at −20°C.
Enterobacterial repetitive intergenic consensus–PCR (ERIC-PCR)
All isolates from the same sample were subjected to ERIC-PCR in order to recognize indistinguishable isolates and to discriminate dissimilar isolates, which have been chosen for sequencing the gyrB gene. The oligonucleotides selected for ERIC-PCR were ERIC 1R (5’-ATGTAAGCTCCTGGGGATTCAC-3’) and ERIC 2 (5’-AAGTAAGTGACTGGGGTGAGCG-3’) (Versalovic et al., 1991). PCR amplification was performed using a PTC100™ Peltier Thermal Cycler (MJ Research, St. Bruno, Quebec, Canada). Each reaction was performed in a final volume of 25 μL, containing 2.5 μL of 10× PCR buffer, 50 pmol of each primer, 400 μM of deoxynucleotide triphosphates (dNTPs), 1 μL (1U) of Taq DNA polymerase, and 2.5 μL (50 ng) of template DNA. The reaction mixtures were subjected to an initial denaturation for 4 min at 94°C followed by 35 cycles, with a thermal cycling of 94°C for 15 s, 55°C for 30 s, 72°C for 45 s, and a single final extension for 5 min at 72°C. Following amplification, 8 μL of the PCR products were electrophoresed on 1.2% agarose gels in TAE buffer. Electrophoresis was performed for 80 min at 6 V/cm. A 1-kb DNA ladder was used as molecular size marker. Agarose gels were stained on ethidium bromide (0.5 μg/mL), visualized with an ultraviolet lamp, and photographed in the GelDocMega (Wolf Laboratories Limited, Pocklington, UK).
PCR amplification and sequencing of gyrB
PCR amplification was performed using a PTC100™ Peltier Thermal Cycler (MJ Research). Procedures and characteristics of oligonucleotide primers (gyrB3F 5’-TCCGGCGGTCTGCACGGCGT-3’; gyrB14R 5’-TTGTCCGGGTTGTACTCGTC-3’) for the PCR-based amplification and sequencing of the gyrB gene were performed as previously described (Soler et al., 2004; Yañez et al., 2003), except for using the BigDye Terminator V3.1 Cycle Sequencing Kit in the ABI 3100-Avant Genetic Analyzer (Applied Biosystems, Carlsbad, CA), according to the manufacturer's instructions.
Phylogenetic data analysis
The gyrB nucleotide sequences were aligned to previously published reference sequences (Saavedra et al., 2006; Soler et al., 2004; Yañez et al., 2003) by using the Clustal X program, version 1.8 (Thompson et al., 1997). Genetic distances were determined by Kimura's two-parameter model (Kimura, 1980), and an evolutionary tree was constructed by the Neighbour-Joining method (Saitou and Nei, 1987) with the Mega program (Kumar et al., 2001).
Results
In this study, Aeromonas spp. were detected in 10 (31%) of the 32 samples analyzed, and a total of 84 presumptive Aeromonas isolates were obtained. The 84 isolates of Aeromonas spp. were subjected to ERIC-PCR analysis, and 20 isolates presented different and unique patterns. Nevertheless, isolate redundancy (i.e., pairs of strains with identical genetic pattern) was found in the remaining 64 isolates. Six of the 20 different patterns included just one isolate each, three included two, and the number of isolates included in the other 11 different patterns varied between three and 15 (Table 1).
The 20 isolates with different patterns were taken for species identification by gyrB gene sequencing, and we have found 18 isolates with different gyrB sequences. The gyrB sequences obtained (427–486 nt in length) were aligned with those from reference strains of the Aeromonas species, and a phylogenetic tree was constructed (Figs. 1 and S1) (see supplementary Fig. S1 at
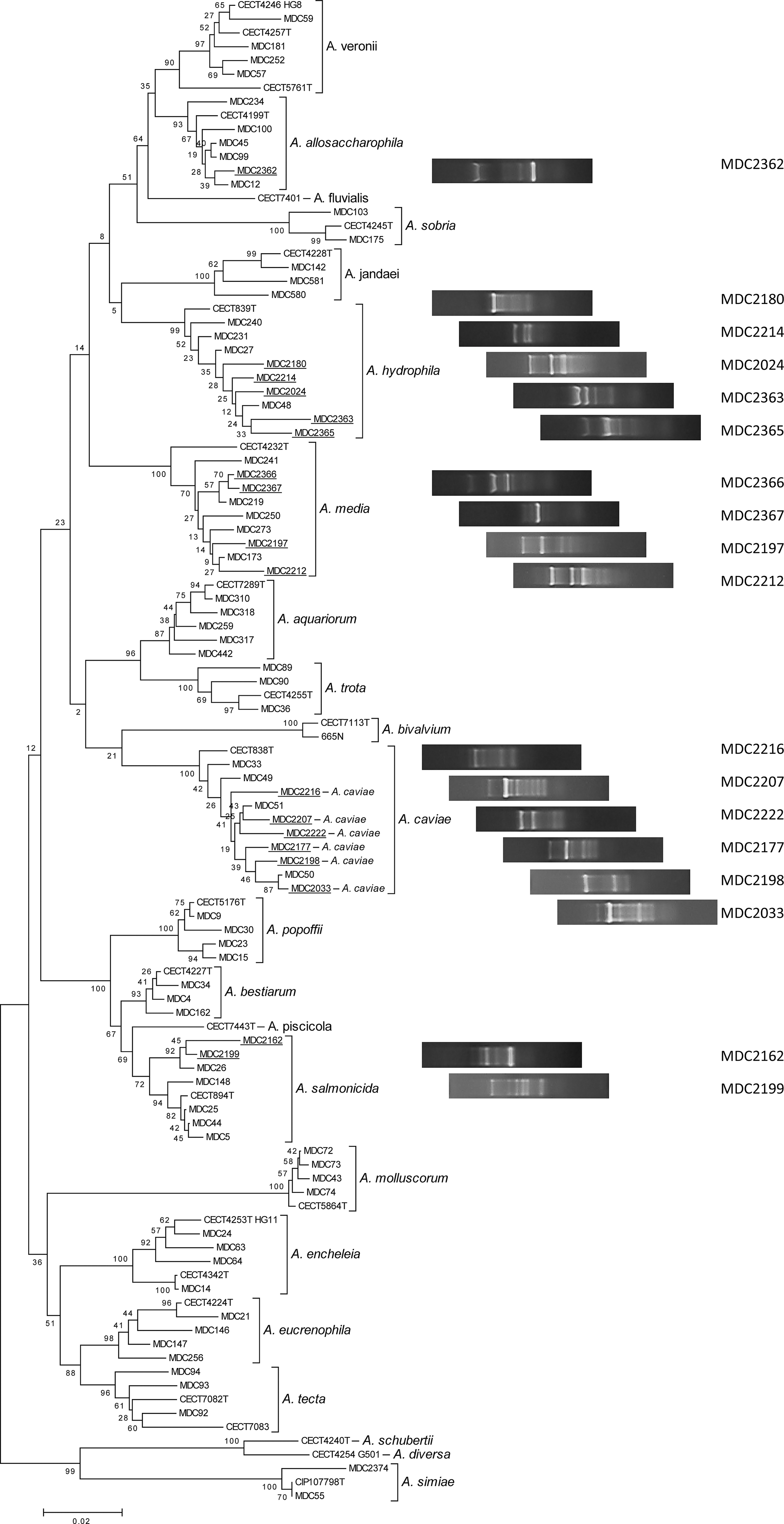
Phylogenetic tree (NJ) showing all the strains of Aeromonas hydrophila, A. salmonicida, A. caviae, A. media, and A. allosaccharophila obtained (underlined) and other known Aeromonas species based on the gyrB gene sequences and the type strains of each corresponding species. Numbers shown next to each node indicate bootstrap values (percentage of 1000 replicates). Enterobacterial repetitive intergenic consensus–PCR (ERIC-PCR) profiles of the strains obtained in this study are shown.
It was found that 60% of the positive samples presented more than one Aeromonas species, and in one of them we isolated and identified three different species: A. salmonicida, A. caviae, and A. media.
Discussion
Aeromonas are ubiquitous in nature, and their presence in food has been reported in an increasing number of studies, from different geographical regions. Aeromonas species have been detected in different food products, including meat products (Hudson and De Lacy, 1991; Neyts et al., 2000; Singh, 1997; Villari et al., 2000). In a study describing the distribution of mesophilic Aeromonas species in raw and ready-to-eat fish and meat products in Switzerland by Gobat and Jemmi (1993), sliced cooked ham, smoked cooked sausage, and mortadella showed a occurrence of 38.2%, 15.6%, and 12.9%, respectively, for Aeromonas spp. In Italy, 25% of salami and raw ham samples were positive for Aeromonas spp., with A. hydrophila and A. caviae being the only species detected in meat products (Villari et al., 2000). Hudson and De Lacy (1991) reported the presence of A. hydrophila and A. caviae in 18% and 4.5%, respectively, of the sausages analyzed. Singh (1997) isolated Aeromonas spp. from all sausage samples analyzed, which belong to the species A. hydrophila, A. caviae, and A. sobria. These Aeromonas species were also detected in pork sausages and ground pork by Okrend et al. (1987).
In the present study, 31% of the alheira samples analyzed, all from the same factory, were positive for Aeromonas spp., and five different Aeromonas species have been identified: A. hydrophila, A. salmonicida, A. caviae, A. media, and A. allosaccharophila.
The gyrB gene sequencing was very useful in this study for the identification of Aeromonas species, which confirms the usefulness of this methodology in discriminating Aeromonas, including the identification of strains of rarely isolated species such as A. allosaccharophila (Saavedra et al., 2007).
Generally, alheira is characterized by high variability in its chemical, physical, and sensory attributes (Patarata et al., 2008), and it is important to note that this traditional product is submitted to a smoking process at low temperature (generally<40°C) (Albano et al., 2008). According to Ferreira et al. (2006), alheira exhibited pH and salt levels insufficient to assure microbiological safety, although this traditional meat product is not considered a ready-to-eat food and is usually eaten after being cooked (roasted or fried). Because of this, even when contaminated with pathogens, including Aeromonas, these would be inactivated. However, very often, alheira is consumed after being submitted to culinary treatments, whose time/temperature combination is insufficient to inactivate pathogens. On the other hand, there is always ample opportunity for post-cooking contamination (Ferreira et al., 2006). In these cases, a contaminated alheira may represent a threat to consumers' health, especially to the young, the elderly, and the immunocompromised.
Conclusion
In this work, the phylogenetic diversity of Aeromonas isolated from alheira has been reported. Despite the increasing number of studies on the occurrence of Aeromonas in food, to our knowledge it is the first time that the presence and diversity of these bacteria are described in this traditional Portuguese meat product. We would like to emphasize the identification of clinically relevant species, i.e., A. hydrophila and A. caviae, among the species isolated. The results demonstrate the usefulness of gyrB sequencing in discriminating Aeromonas species.
Footnotes
Acknowledgments
This work was supported by the Foundation for Science and Technology (FCT Ph.D. grant SFRH/BD/25415/2005 and strategic research project PEst-OE/AGR/UI0772/2011) and by IMPIVA, Generalitat Valenciana, Spain (grants IMIDTA/2007/68, IMIDTA/2008/104, IMIDTP/2009/64). Services were supplied by the Molecular Diagnostics Center (MDC), Orihuela, Spain.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.