
Abstract
Fourteen Campylobacter jejuni genes—porA, cadF, omp18, dnaK, flaC, peb1, peb2, peb3, peb4, ahpC, groEL, tuF, hipO, and Cj0069—were cloned and expressed in Escherichia coli BL21. The recombinant proteins were purified on histidine (His) and glutathione S–transferase (GST) trap columns using the ÄKTA Explorer 100 System. Recombinant proteins were visualized using sodium dodecyl sulfate–polyacrylamide gel electrophoresis and identified using matrix-assisted laser desorption ionization time-of-flight mass spectrometry. The antigenicities of these recombinant proteins were assessed by Western blotting and enzyme-linked immunosorbent assays with anti–C. jejuni immune rabbit sera. Four recombinant proteins, including rGST-PorA, rHis-CadF, rGST-GroEL, and rGST-TuF, demonstrated reactions with both anti-serum and preimmune serum, while rHis-DnaK, rGST-FlaC, rGST-PEB2, rGST-PEB3, rGST-PEB4, and rGST-HipO showed variable antigenicity characteristics to the anti-sera derived from different C. jejuni strains. rHis-Omp18, rHis-PEB1, and rGST-AhpC demonstrated universal and specific antigenities with the entire anti-sera panel tested in this present study, while recombinant rGST-Cj0069 and rHis-DnaK did not react with any of the anti–C. jejuni sera tested. In conclusion, rGST-AhpC may be useful as a potential serodiagnostic antigen for C. jejuni infection.
Introduction
Serological evidence for the diagnosis of Campylobacter enteritis is limited; however, such diagnoses play a crucial role in the study of antecedent causes of GBS, as well as in determining the role of C. jejuni in cases of reactive arthritis. Conventional serological assays for C. jejuni commonly use crude antigenic preparations, such as acid-glycine extracts and whole-cell lysates (Kosunen et al., 1983; Logan and Trust, 1983; Rautelin and Kosunen, 1983; Rautelin and Kosunen, 1987). So far, only a few definite proteinaceous antigens have been characterized from C. jejuni (Burnens et al., 1995; Pei et al., 1991; Schmidt-Ott et al., 2006). A more contemporary approach employs the production of recombinant antigens that are used to identify antigens useful for diagnosing C. jejuni infection. In our laboratory, a previous investigation of whole cell proteins from C. jejuni using Western blotting and immune serum showed that some proteins demonstrate positive reactions with anti–C. jejuni serum (unpublished data). The aim of this present study was to obtain potential candidate proteins that may be useful for the serological diagnosis of C. jejuni infection by assessing the antigenicities of 14 recombinant C. jejuni proteins expressed in Escherichia coli.
Methods
Bacterial strains and culture conditions
The C. jejuni isolates used in this present study were clinical isolates from China, and these were sorted into three groups (A, B, and C) for rabbit immunizations. The background of each C. jejuni isolate is listed in Table 1. The bacteria were cultured on Columbia agar supplemented with 5% sheep blood under microaerophilic conditions (5% O2, 10% CO2, and 85% N2) at 42°C for 24 h. Bacterial cells were harvested from agar plates and resuspended in sterile saline (0.9% NaCl). Then, the cells were washed in saline three times by centrifugation each time at 4,000 r.p.m. for 5 min at 4°C.
GBS, Guillain-Barré syndrome.
Production of anti–C. jejuni sera
For each immunization experimental group, the immunization amount for each C. jejuni isolate was mixed equally. In each group, four adult New Zealand white rabbits were immunized by injection of 1 mL of formalin-killed C. jejuni suspensions at 109 cells/mL (from 24-h cultures) into the ear vein. Three identical doses were given at 10-day intervals. Increases in the titer of the specific antibody were monitored by enzyme-linked immunosorbent assay (ELISA) with the whole cell lysate antigen. The polyclonal antiserum was collected once the titer was greater than 1:100,000. Pre-immune serum was taken and used as control serum. Immunoserum for Helicobacter pylori was obtained from previous immunizations in our laboratory, while immune-sera for Yersinia enterocolitica, E. coli O157:H7, and Vibrio cholerae were gifted by other laboratories in our Institute. In total, 29 clinical serum samples were used for the antigenicity assessments of the potential recombinant proteins, and these included 17 negative and 12 positive sera to anti–C. jejuni antibodies.
Protein expression and preparation
The nucleotide sequence of each target gene was downloaded from the National Center for Biotechnology Information (NCBI) GenBank for C. jejuni isolates ICDCCJ07001 and NCTC11168. Specific DNA comparisons were carried out using the Vector NTI Suite 6 software. The polymerase chain reaction (PCR) products (without the regions encoding signal peptides) of porA, cadF, omp18, dnaK, flaC, peb1, peb2, peb3, peb4, ahpC, groEL, tuF, hipO, and Cj0069 from C. jejuni ICDCCJ07001 or NCTC11168 were cloned into the pMD18-T vector (D103A; Takara, Dalian, China). The genes and primers used for PCR are summarized in Table 2. Target DNA fragments were excised from recombinant T-vectors using BamHI and XhoI, and subsequently cloned into pET30a, pET32a (69015-3; Novagen, Darmstadt, Germany), or pGEX-4T-1 vectors (28-9545-49; GE Healthcare, Little Chalfont, UK). The resultant recombinant plasmids were transformed into E. coli BL21. Successfully transformed E. coli BL21 were screened on Luria-Bertani agar containing 100 μg/mL ampicillin. The expression of each recombinant protein was induced by 200 μg/mL isopropyl b-
NCBI, National Center for Biotechnology Information; MALDI TOF, Matrix-Assisted Laser Desorption/Ionization–Time of Flight; CI, confidence interval.
Protein identification and purification
The proteins were pooled, and 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) was performed. Protein bands were visualized with Coomassie Brilliant Blue G-250 and excised from the gel. In-gel digestion and protein identification was carried out using matrix-assisted laser desorption ionization time-of-flight (MALDI-TOF/TOF) mass spectrometry, as described previously (Zhang et al., 2009). Protein identity was defined when the total protein score confidence interval (CI) percentage was >95%. Target proteins were purified using HisTrap™ HP (GE Healthcare, catalog no. 17-5247-01) and GSTrap™ HP columns (GE Healthcare, catalog no. 17-5282-02) on an ÄKTA Explorer 100 System according to the manufacturers' instructions. All fractions were analyzed by SDS-PAGE, and those containing greatest concentrations of protein were pooled. Protein concentrations were determined by measuring absorbance at 280 nm with a NanoDrop-1000 spectrophotometer (NanoDrop Technologies Inc., Wilmington, DE).
Western blotting and ELISAs
Ten micrograms of each purified protein was subjected to 12% SDS-PAGE and then transferred from the gel to a polyvinylidene fluoride (PVDF) membrane. Transfers were performed for 1 h at 20 V. PVDF was blocked with 5% skim milk in Tris-buffered saline (0.02 M Tris base, 0.14 M NaCl, and 2.7 mM KCl; pH 7.4). Immunoblotting was carried out with the primary antibodies using three groups of rabbit polyclonal antisera at a final dilution of 1:200. The secondary antibody was horseradish peroxidase–labeled goat anti-rabbit IgG (ZB-2301; Zhongshanjinqiao, China) diluted to 1:2,000 in washing buffer. Reactive proteins were detected by diamino benzidine (DAB) staining.
In order to confirm specific reactions, ELISAs were performed between the purified rGST-AhpC protein and the following rabbit immunosera: anti–C. jejuni, anti–H. pylori, anti–Y. enterocolitica, anti–E. coli O157:H7, and anti–V. cholerae. Briefly, 1 μg of purified recombinant rGST-AhpC was coated on to the surface of each well of 96-well immuno-ELISA plates (Corning Costar 2592). The plates were blocked with bovine serum albumin by overnight incubation at 4°C. The immunosera were diluted from 1:200 to 1:102,400, and added to triplicate wells before incubation at 37°C for 1 h. Then, the plates were washed and horseradish peroxidase-conjugated goat anti-rabbit IgG diluted to 1:2,000 was added as the secondary antibody. Following a 1-h incubation, captured antibodies were detected using tetramethyl benzidine as substrate and measuring optical density (OD) at 450 nm. Pre-immunoserum was used as the negative control. A total of 29 selected clinical serum samples were selected for ELISA testing, and these included 17 anti–C. jejuni negative sera from diarrheal patients and 12 anti–C. jejuni positive sera from GBS patients. The sera were diluted to 1:500. The secondary antibody used was goat anti-human IgG (A0170; Sigma, St. Louis, MO) that was diluted to 1:2,000. ELISAs were performed in triplicate according to standard protocols.
Results
Protein expression and identification
The recombinant expression plasmids were identified by enzyme digestion with BamHI and XhoI. The sizes of the inserted DNA fragments were consistent with predictions. Thirteen of the 14 recombinant proteins were expressed as soluble proteins, and only rGST-Cj0069 formed inclusion bodies. MALDI-TOF/TOF analyses showed that the expressed proteins in E. coli BL21 were the targeted proteins from C. jejuni. The characteristics of each recombinant protein are listed in Table 2.
Antigenicity characteristics of the recombinant proteins
In total, 300 mL of anti-serum was collected from each immuno rabbit group. The titers of the immunosera were greater than 1:100,000. Antigenicity characteristics of each recombinant protein were determined using three groups of anti–C. jejuni sera. A selection of Western blot results is shown in Figure 1. The immunoreactivities between the recombinant proteins and the different immunosera are summarized in Table 2. The mean optical densities from ELISAs of immunosera and clinical sera are summarized in Figures 2 and 3, respectively. rGST-AhpC specifically reacted with the anti–C. jejuni sera. The mean and minimum OD values from positive clinical serum were 0.34 and 0.20, respectively, while the mean and maximum OD values of negative clinical serum were 0.06 and 0.13, respectively. Thus, the mean OD value of clinical positive serum was five times greater than clinical negative serum.
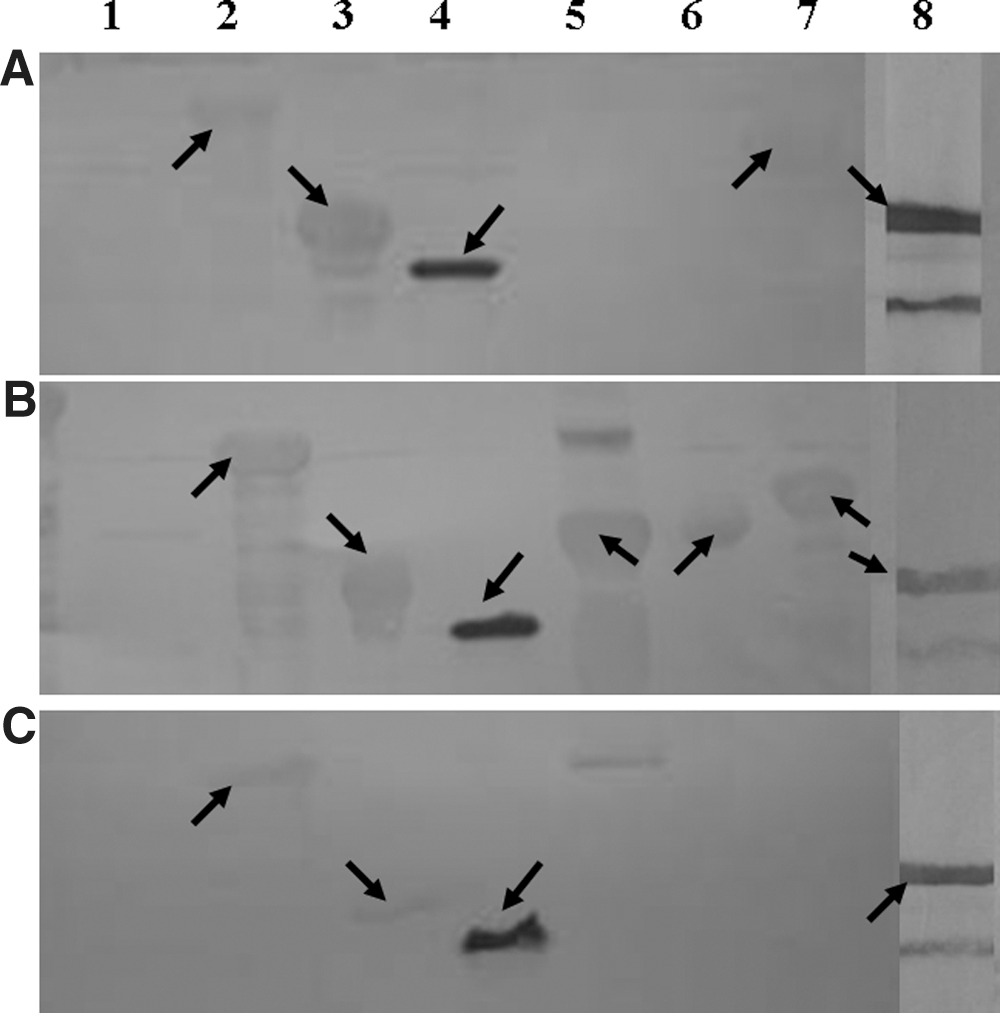
Western blot of some of the recombinant proteins. Purified expressed proteins (10 μg of each) were subjected to 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred from the gel to a polyvinylidene fluoride (PVDF) membrane. Immuno-blotting was carried out with primary antibodies using three groups of rabbit polyclonal anti– Campylobacter jejuni sera at a final dilution of 1:200.

Enzyme-linked immunosorbent assay (ELISA) results of immuno sera. ELISAs were performed between purified rGST-AhpC and anti–Campylobacter jejuni, anti–Helicobacter pylori, anti–Yersinia enterocolitica, anti–Escherichia coli O157:H7, and anti–Vibrio cholera immuno rabbit sera. In each case, 1 μg of purified rGST-AhpC was coated onto the wells of 96-well ELISA plates. The immuno sera were diluted from 1:200 to 1:102,400. Specific IgG antibodies were measured by optical density at 450 nm. ELISAs were performed in triplicate using a standard protocol.
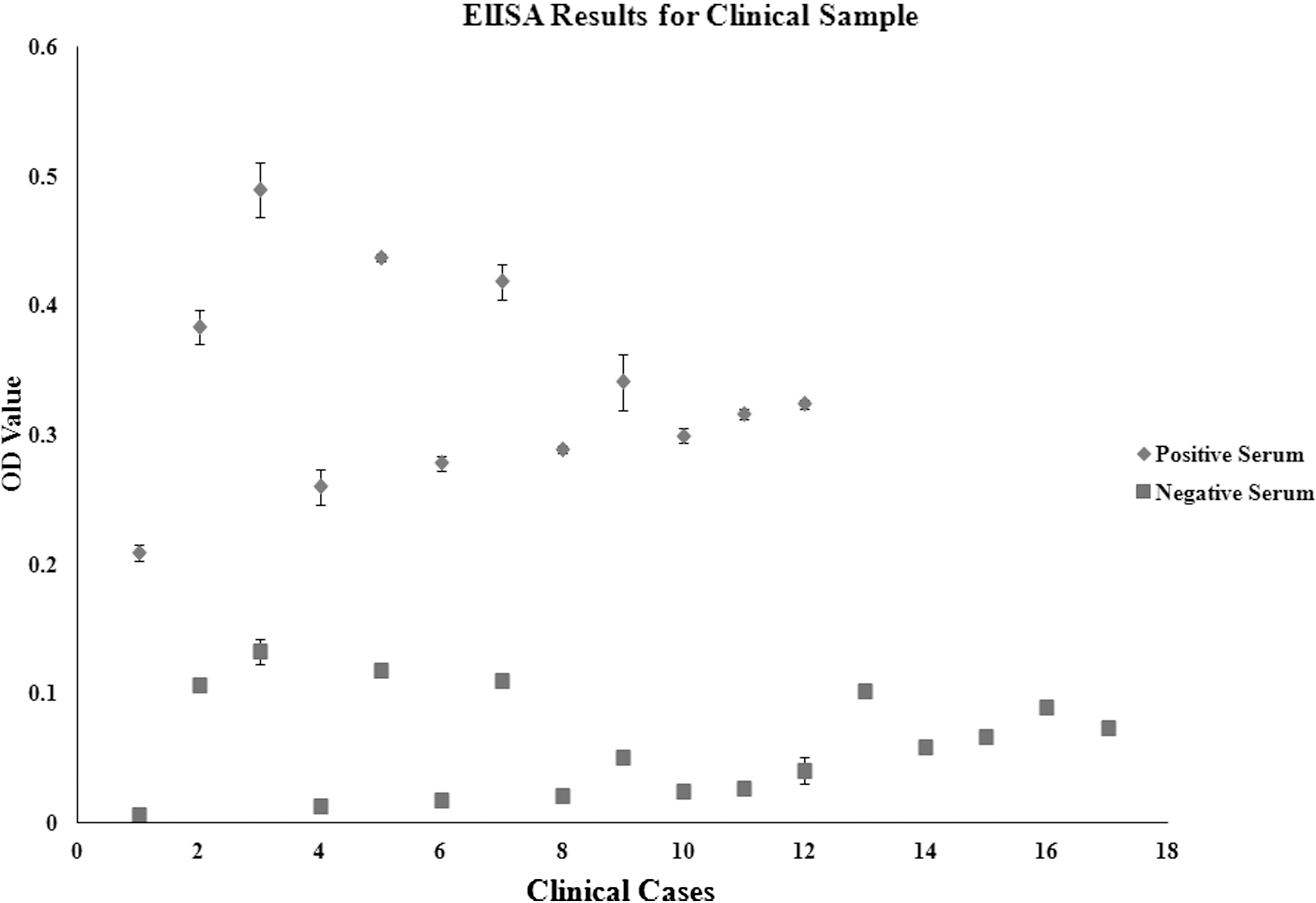
Enzyme-linked immunosorbent assay (ELISA) data from selected clinical samples. ELISA tests were performed between purified rGST-AhpC and 29 sera selected from clinical samples. In each case, 1 μg of purified rGST-AhpC was coated onto the wells of 96-well ELISA plates. The clinical sera were diluted to 1:500. Specific IgG antibodies were measured by optical density at 450 nm. ELISAs were performed in triplicate using a standard protocol.
Discussion
The purpose of this present study was to obtain recombinant protein antigen candidates that may be useful for the serological diagnosis of C. jejuni infection in China. In this present study, three groups of C. jejuni isolates, including seven clinical isolates from China, were used for the production of anti–C. jejuni sera. C. jejuni ICDCCJ07001 (assigned to group A) was the isolate that triggered the GBS outbreak in China in 2007 (Zhang et al., 2010). C. jejuni HB-CJGZHANGB, HB-CJGLXC, and HB-CJGZHANX (assigned as group B) were from sporadic GBS patients caused by prior C. jejuni infection in China (Jiang et al., 2010). C. jejuni BJ-CJD158, BJ-CJD101, and BJ-CJD70 (group C) were from diarrheal patients in China. Although a limited number of isolates was examined, these isolates still exhibited appreciable diversity for the C. jejuni strains found in China, particularly the isolates from GBS patients.
DnaK and GroEL are heat shock family proteins. Aside from the stress response, these proteins play important roles as immunodominant antigens during bacterial infections. Previous reports have indicated that recombinant DnaK is immunoreactive with sera from C. jejuni–infected patients but only in a minority of cases (Thies et al., 1999). The recombinant rHis-DnaK obtained in this present study did not react with any of the anti-sera. GroEL is the most prominent antigen and virulence factor for many enteric pathogens, and it is also a potential immunogen in C. jejuni (Wu et al., 1994). In this present study, rGST-GroEL showed cross-reaction with pre-immunoserum. This result indicates that GroEL might not be suitable as a diagnostic antigen for C. jejuni infection, though it might be useful as a vaccine component.
PorA is the major outer membrane protein of C. jejuni. It is surface exposed and seems to be immunogenic to a certain extent during infection. A previous report indicated that recombinant rGST-PorA provided appreciable protection against colonization by different C. jejuni isolates (Islam et al., 2010). However, in this present study and similar to GroEL, PorA showed cross-reaction with anti-sera and pre-immune serum meaning that it cannot be used as a potential diagnostic antigen for the detection of antecedent C. jejuni infection.
FlaC is a protein secreted by C. jejuni through the flagellar filament without special signals. Thus, it is found predominantly in the extra cellular milieu. It binds epithelial cells and plays a crucial role during cell invasion. An earlier study showed that recombinant FlaC is immunogenic, but it provides only limited protection against C. jejuni in a mouse infection model (Baqar et al., 2008). In this present study, rGST-FlaC showed no immunoreaction with any of the immunosera.
To date, only a few surface antigens of C. jejuni have been well characterized genetically and immunogenically, and these include Omp18, PEB1, PEB2, PEB3, and PEB4 (Burnens et al., 1995; Du et al., 2008; Pei et al., 1991). Purified PEB1 and PEB3 appear to be good candidate proteins to detect antibody responses for C. jejuni infection. However, immunoreaction varies between patient sera and PEB2 and PEB4. In order to verify the antigenicity profiles of PEB1, PEB2, PEB3, and PEB4 from local isolates, recombinant proteins were produced. In this present study, recombinant rHis-PEB1 and rHis-Omp18 (another membrane protein) reacted positively only with sera from C. jejuni–infected patients and not the healthy controls, indicating that these proteins could be used as diagnostic antigens for C. jejuni infection. Meanwhile, the antigenicities of PEB2, PEB3, and PEB4 might be strain dependant.
The hippurate hydrolase gene (hipO), found in C. jejuni only and not in other Campylobacter spp., is typically used as a DNA identification marker for C. jejuni. However, in this present study, rGST-HipO showed antigenicity characteristics with only some of the anti-sera tested.
The protein encoded by Cj0069 has been shown to be more highly expressed in C. jejuni at 37°C compared with 42°C (Zhang et al., 2009). An earlier report showed that recombinant rTEV-Cj0069 could be a sensitive and specific antigen for the detection of anti–C. jejuni IgG antibody in patients infected with C. jejuni (Corso et al., 2011). However, the results of this present investigation showed that negative reactions occurred between recombinant rGST-Cj0069 and the entire panel of immunosera.
Our previous results showed that the elongation factor Tu (TuF), the outer membrane protein cadF, and one subunit of alkyl hydroperoxide reductase (AhpC) presented strong reactions with immuno rabbit serum. In this present study, the immunoblotting results showed that GST-TuF and rHis-CadF cross-reacted with both anti-serum and pre-immune serum, while rGST-AhpC showed a specific reaction with the entire panel of various anti-sera.
The results from the immune sera and the selected clinical samples confirmed the specificity of the reactions between rGST-AhpC and the anti–C. jejuni antibody, which indicates that recombinant rGST-AhpC might be a particularly useful antigen for serologic diagnosis post–C. jejuni infection. However, in this present study, only a few non-Campylobacter pathogen immunosera were included, and cross-reactivity between rGST-AhpC and other Campylobacter spp. remains to be determined. In addition, the limitation of using anti-serum raised by C. jejuni injection for evaluating candidate serologic antigens remains. Further investigations are needed on additional anti–Campylobacter spp. sera and larger sets of sera from clinical cases. AhpC is reported to be an important determinant for the ability of microaerophilic pathogens, such as C. jejuni, to survive oxidative and aerobic stresses (Baillon et al., 1999). This present study demonstrates for the first time that AhpC is a potential serodiagnostic antigen for C. jejuni infection.
Conclusion
In summary, 14 C. jejuni proteins were cloned and expressed. The recombinant proteins rGST-PorA, rHis-CadF, rGST-GroEL, and rGST-TuF are not suitable as serodiagnostic antigens; the recombinant proteins rGST-PEB2, rGST-PEB3, rGST-PEB4, and rGST-HipO demonstrated antigenicity against C. jejuni, but the antigenicity appeared to be strain dependent. Nevertheless, rHis-Omp18, rHis-PEB1, and rGST-AhpC seem to be appropriate serologic diagnostic antigens for the detection of the anti–C. jejuni antibodies. This present study provides a platform for further functional and immunogenic studies of these 14 recombinant proteins from C. jejuni.
Footnotes
Acknowledgments
We are grateful to Dr. Huaiqi Jing and Dr. Biao Kan for kindly providing anti–Y. enterocolitica serotype O:8, serotype O:9, anti–E. coli O157:H7, and anti–V. cholera immuno serum. This work was supported by grants from the General Program of the National Natural Science Foundation of China (grant 81071314) and the Young Scientist Foundation from China CDC (grant 2009A102).
Disclosure Statement
No competing financial interests exist.