
Abstract
The objective of this study was to determine the effect of sampling (swab samples compared to destructive samples) on isolation rates of human pathogenic Yersinia enterocolitica from pig tonsils. Moreover, the relative efficiency of different rapid, routinely applicable isolation methods was evaluated. Therefore, swab and destructive samples from tonsils of 120 pigs at slaughter were analyzed in parallel using direct plating and different enrichment methods. Salmonella-Shigella-desoxycholate-calcium chloride (SSDC) agar, cefsulodin-irgasan-novobiocin (CIN) agar, and Yersinia enterocolitica chromogenic medium (YeCM) were used as selective agar media. For enrichment, irgasan-ticarcillin-potassium chlorate (ITC) broth and peptone-sorbitol-bile (PSB) broth were incubated at 25°C for 48 h. Overall, 55 tonsils (45.8%) were positive for Y. enterocolitica bioserotype 4/O:3. Recovery was significantly higher using the destructive method compared to the swabbing method. Direct plating resulted in 47 and 28 Y. enterocolitica–positive destructive and swab samples, respectively. Alkali treatment of PSB and ITC enrichment broths significantly increased recovery of pathogenic Y. enterocolitica from destructive tonsil samples. The performance of YeCM for qualitative and quantitative isolation of pathogenic Y. enterocolitica from pig tonsils was equal to SSDC and CIN. In conclusion, direct plating and ISO 10273:2003 with minor modifications are suitable and rapid methods for isolation of pathogenic Y. enterocolitica from destructive tonsil samples.
Introduction
Y
Culture methods for recovery of Y. enterocolitica rely on initial enrichment followed by plating on selective agar plates. Enrichment at 4°C for several weeks is successful for isolation from different sample types, but for isolation of pathogenic yersiniae, enrichments in selective media at higher temperatures for shorter time periods have been shown to be as effective as the prolonged cold enrichments (de Boer and Nouws, 1991; De Giusti et al., 1995; Kwaga et al., 1990). Isolation of pathogenic Y. enterocolitica from naturally contaminated samples without enrichment is seldom successful (Fredriksson-Ahomaa and Korkeala, 2003); however, for tonsil samples, direct plating is a fast alternative as in most samples background flora on a selective agar medium is minimal and the number of pathogenic Y. enterocolitica is sufficiently high (Fredriksson-Ahomaa et al., 2007; Van Damme et al., 2010). A disadvantage of current conventional media is the difficulty to distinguish pathogenic Y. enterocolitica from non-pathogenic yersiniae and many other Gram-negative bacteria (de Boer, 1992; Fondrevez et al., 2010). To facilitate this differentiation, Weagant (2008) recently designed a chromogenic agar medium, called Y. enterocolitica chromogenic medium (YeCM).
Besides sensitive culture methods, adequate sampling procedures are also fundamental for the success of isolation and, therefore, crucial in monitoring programs. When monitoring Y. enterocolitica in pigs at slaughter, tonsils are regarded as the sample of choice (Fredriksson-Ahomaa et al., 2001; Nesbakken et al., 2003). Swab samples from tonsils have been used in several studies (Andersen et al., 1991; Fondrevez et al., 2010; Kechagia et al., 2007), since sampling and processing is easier and less time-consuming compared to tissue samples. Therefore, this study evaluates if swab samples have a good predictive value for the presence of pathogenic Y. enterocolitica in pig tonsils. Moreover, when screening a large number of animals, short and efficient isolation procedures are indispensable. This study will thus compare different rapid isolation methods to determine the most efficient method for recovery of pathogenic Y. enterocolitica isolates from pig tonsils.
Methods
Sample collection and preparation
In a Belgian abattoir, tonsils from 120 fattening pigs were cut out aseptically immediately after removal of the plug set and the surface area of both tonsils was swabbed using a dry sterile cotton swab. The swab sample was placed in 10 mL of peptone-sorbitol-bile broth (PSB), and for the destructive method, 11 g of tonsillar tissue was homogenized in 99 mL PSB. One and 10 mL of the swab and tissue homogenate was transferred into 9 and 90 mL of irgasan-ticarcillin-potassium chlorate (ITC) broth, respectively. Three milliliters of PSB-homogenate was used for direct plating, and the remaining homogenate was used for enrichment at 25°C for 48h.
Isolation and enumeration
All samples were examined using (i) direct plating and (ii) different enrichment methods (Fig. 1). Three selective agar media were used: (1) Salmonella-Shigella-desoxycholate-calcium chloride agar (SSDC; Conda, Madrid, Spain); (2) cefsulodin-irgasan-novobiocin agar (CIN; Oxoid, Basingstoke, United Kingdom) and (3) Yersinia enterocolitica chromogenic medium (YeCM; prepared according to Weagant [2008]).
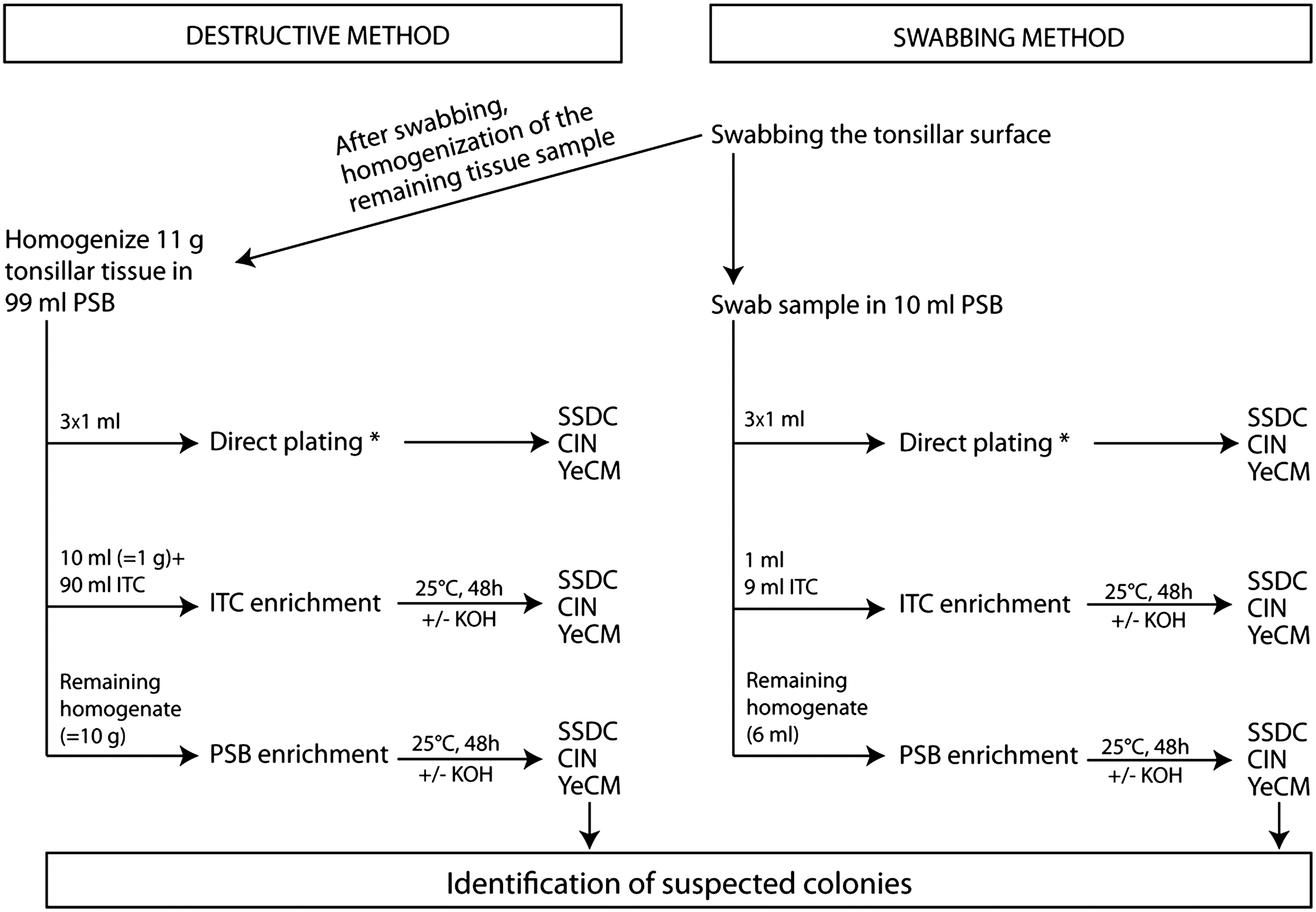
Schematic overview of the applied procedures for isolation of pathogenic Yersinia enterocolitica from pig tonsils. PSB, peptone-sorbitol-bile; ITC, irgasan-ticarcillin-potassium chlorate; SSDC, Salmonella-Shigella deoxycholate calcium chloride; CIN, cefsulodin-irgasan-novobiocin; YeCM, Y. enterocolitica chromogenic medium. *Direct plating: 1 mL of PSB homogenate is plated on each of the three selective agar media: 0.9 mL is spread plated over two plates, and 0.1 mL is inoculated on a third plate using the spiral plate machine.
(i) For direct plating, 0.4 and 0.5 mL of the homogenate was spread plated on two plates of each of the three agar media. Moreover, 0.1 mL of the homogenate was plated on each of the selective plates using a spiral plate machine (Eddie Jet, IUL Instruments, Barcelona, Spain). All plates were incubated at 30°C for 24 h and examined for typical Yersinia colonies using a stereomicroscope with Henry illumination. Suspected colonies were enumerated and identified as described below.
(ii) After 48 h of enrichment in ITC and PSB at 25°C, a loopful was streaked directly and after alkali treatment onto each of the three different agar plates. All inoculated selective plates were incubated at 30°C for 24 h. Suspected colonies were identified as described below.
Biochemical identification and biotyping
Suspected Yersinia colonies were grown on plate count agar (PCA) and inoculated in urease broth, Kligler Iron agar, and tryptic-casein-soy broth and incubated at 30°C for 24 h. Isolates showing typical reactions were further identified using lysine and ornithine decarboxylase, sucrose fermentation, and Voges-Proskauer reaction at 30°C. Isolates showing atypical reactions were tested using API 20E strips (Bio-Mérieux, Marcy l'Etoile, France), incubated at 25°C for 18–20 h. Y. enterocolitica isolates were biotyped based on the revised scheme of Wauters et al. (1987) using esculin hydrolysis, indole production, and xylose fermentation. Y. pseudotuberculosis isolates were biotyped using melibiose, citrate, and raffinose (Tsubokura and Aleksic, 1995) and sent to the Belgian reference laboratory for Y. pseudotuberculosis (UCL, St-Luc Hospital, Brussels) for serotyping.
Molecular identification
The pathogenicity of Y. enterocolitica isolates was confirmed using a multiplex PCR with primers targeting the chromosomal virulence genes ail and yst and the plasmid virulence gene virF according to Harnett et al. (1996). A single PCR was performed to identify Y. enterocolitica serotype O:3 using primers targeting the rfbC gene (Weynants et al., 1996). Y. pseudotuberculosis isolates were identified using a PCR assay targeting the inv-gene (Nakajima et al., 1992).
DNA was extracted using PrepMan reagent (Applied Biosystems, Foster City, CA) according to the manufacturer's instructions, and 1 μL of the supernatant was used as a template in each PCR assay. DNA amplification was performed in a 50-μL reaction mixture containing 2.5 U GoTaq® Flexi DNA Polymerase, 1×green reaction buffer, 1.5 mM MgCl2 (Promega, Madison, WI), 200 μM of each dNTP (Invitrogen, Carlsbad, CA), and 0.5 μM of each primer. PCR was performed in a Veriti™ 96-Well Thermal Cycler (Applied Biosystems). Each cycle consisted of pre-denaturation at 94°C for 5 min, 32 cycles of denaturation at 94°C for 30 s, primer annealing at 55°C for 30 s, and extension at 72°C for 1 min, followed by a final extension at 72°C for 7 min. For inv, only the annealing temperature (58°C) was changed. PCR products were electrophoresed in agarose gels, stained with ethidium bromide, and visualized by ultraviolet light.
Statistical analysis
Statistical analysis was carried out using STATA 11.0 (Stata Corporation, College Station, TX). Differences between isolation methods were examined using a random-effects logistic regression analysis. The mean load was calculated for each agar medium separately, computing the mean of the log-values, and taking into account the countable plates only. Counts on the three different agar plates were compared using a random-effects negative binomial regression analysis. Samples that were negative for Y. enterocolitica by direct plating using the three different agar media were not included in the analysis. Values of plates that were not countable were replaced by the average count on the particular agar medium. For plates that were negative by direct plating, a value of 0 log10 colony-forming units (CFU) per gram or per surface area was used.
Results
The effect of sampling on isolation of pathogenic Y. enterocolitica from pig tonsils was evaluated by analyzing in parallel swab samples and destructive tissue samples from 120 slaughter pig tonsils. In total, Y. enterocolitica bioserotype 4/O:3 was isolated from tonsils of 55 slaughter pigs (45.8%). Recovery using destructive samples was significantly higher compared to swab samples (OR=14.0 [3.6–54.7]) (Table 1). For 16 tonsils, only the destructive sample was positive, whereas for three tonsils, Y. enterocolitica was isolated from the swab sample only.
NT, not tested; SSDC, Salmonella-Shigella deoxycholate calcium chloride; CIN, cefsulodin-irgasan-novobiocin; YeCM, Y. enterocolitica chromogenic medium; ITC, irgasan-ticarcillin-potassium chlorate; PSB, peptone-sorbitol-bile.
There was no significant difference in recovery between the three agar plates for any of the isolation procedures (p>0.05). For the majority of enrichment procedures, alkaline treatment resulted in a significant higher recovery (p<0.01) compared to isolation without alkaline treatment. However, alkaline treatment did not enhance recovery after ITC enrichment of swab samples (p>0.05).
For destructive samples, more samples were positive using ITC compared to PSB enrichment. This difference was significant for all plates without alkaline treatment (p<0.001); however, it was not the case when using alkaline treatment (p>0.05). Direct plating on SSDC was more efficient (p=0.045) compared to isolation on SSDC after ITC enrichment without alkaline treatment. Only one sample that was positive after ITC-SSDC was not detected using direct plating on SSDC. Isolation rates after ITC enrichment on CIN and YeCM plates without alkaline treatment were also lower compared to direct plating on CIN (p=0.008) and YeCM (p=0.001), respectively. Nevertheless, for destructive tonsil samples, there was no significant difference between direct plating and any of the enrichment methods when using alkaline treatment (p>0.05).
In contrast to destructive samples, more swab samples were positive after PSB enrichment compared to ITC enrichment (p<0.05). Recovery on SSDC and CIN after PSB-enrichment and alkaline treatment was higher than after direct plating on SSDC (p=0.003) and CIN (p=0.010), respectively. This difference was not significant for YeCM (p=0.097).
Direct plating of tonsils also allowed quantification of enteropathogenic Yersinia spp. The mean load of Y. enterocolitica bioserotype 4/O:3 in destructive samples yielding enumeration data was 4.23±0.99 log10 CFU/g (n=29) using SSDC, 4.13±0.93 log10 CFU/g (n=38) using CIN, and 4.25±0.79 log10 CFU/g (n=37) using YeCM agar plates. For countable swab samples, the number of pathogens on the complete surface area was 2.72±1.09 log10 CFU (n=16) using SSDC, 2.62±0.99 log10 CFU (n=24) using CIN, and 2.55±1.10 log10 CFU (n=22) using YeCM agar plates. No significant difference between counts on the three different agar plates was observed (p>0.05). As such, only the frequency distribution of Y. enterocolitica counts using CIN agar plates is shown in Figure 2. Twelve samples that were contaminated with more than 102 CFU per gram tonsillar tissue in the destructive sample were negative by the swab sample.
Frequency distribution of Yersinia enterocolitica bioserotype 4/O:3 counts in tonsil samples based on countable cefsulodin-irgasan-novobiocin (CIN) agar plates using direct plating (n=38 for destructive samples, n=24 for swab samples).
Altogether, 963 Y. enterocolitica bioserotype 4/O:3 isolates were recovered, of which 10 isolates (all originating from the same pig) were ornithine negative. All isolates were positive for the ail and yst gene, and in 780 isolates (81.0%), the virF gene was detected (pYV+). For isolates recovered after ITC and PSB enrichment, 77/393 (19.6%) and 62/337 (18.4%) isolates did not contain the virF gene (pYV-), respectively.
Y. pseudotuberculosis bioserotype 1/O:2 was isolated from the destructive tonsil sample of one pig (0.83%) after direct plating on CIN and YeCM agar plates, on which enumeration revealed a concentration of 3.66 and 3.45 log10 CFU/g tonsillar tissue, respectively. The two Y. pseudotuberculosis isolates were positive for the inv and virF genes, but negative for the ail, yst and rfbC gene.
Discussion
Evaluation of sampling methods for pig tonsils showed a higher frequency of Y. enterocolitica bioserotype 4/O:3 using destructive samples (43.3%) compared to swab samples (32.5%). Our results are in agreement with the results by Nesbakken (1985), who also found more positive tonsillar tissue samples compared to swab samples, applying different isolation methods. Moreover, several highly contaminated destructive samples were negative when the corresponding swab sample was analyzed. As Y. enterocolitica are located in the tonsillar crypts and lymph noduli (Shiozawa et al., 1991; Thibodeau et al., 1999), swabbing the tonsillar surface conceivably fails picking up these bacteria. Since swab samples do not represent internal contamination accurately, they are likely to underestimate the actual carriage of pathogenic Y. enterocolitica in pig tonsils. The International Standard Organization (ISO) method for isolation of presumptive pathogenic Y. enterocolitica from food samples (ISO 10273:2003) is also recommended for pig tonsils (EFSA, 2009). This study shows that the ISO method can be used for tonsil samples. However, some minor modifications are suggested. First, 5-day PSB enrichment was omitted in this study, as for destructive tonsil samples 5-day enrichment is less efficient than 2-day PSB enrichment (Van Damme et al., 2010). Second, ITC provided better results than PSB for destructive samples, whereas for swab samples, PSB enrichment resulted in more positive samples compared to ITC. Remarkably, 15 PSB-positive swab samples that were negative after ITC enrichment were either negative by direct plating or had counts less than 20 CFU/surface area (data not shown). Thus, this lower recovery using ITC compared to PSB enrichment for swab samples may rather be due to the smaller sample fraction that is used for ITC than to an inferior efficiency of this medium. Third, ISO 10273:2003 prescribes the use of alkali treatment after PSB enrichment, but not after ITC enrichment. Nevertheless, our data suggest that alkaline treatment after ITC enrichment is beneficial for isolation of pathogenic Y. enterocolitica from destructive tonsil samples, including SSDC agar. Nine destructive samples that were positive by direct plating were not detected by the ITC-SSDC method. However, when using alkaline treatment, this number was reduced to three. As a conclusion, direct plating or additional KOH treatment after ITC enrichment is thus recommended instead of the conventional ITC-SSDC method.
SSDC agar plates are usually used in combination with ITC enrichment, as recovery is usually higher compared to the ITC-CIN combination (de Boer and Nouws, 1991; Wauters et al., 1988). However, in this study, there was no difference between the agar plates after enrichment in ITC. For tonsil samples, plating on CIN after ITC enrichment seems to be equally good as SSDC for isolation of Y. enterocolitica serotype O:3.
Isolation of pathogenic Y. enterocolitica from tonsillar tissue using enrichment procedures generally results in more positive samples compared to direct plating (Fredriksson-Ahomaa et al., 2001; Korte et al., 2004). However, in this study, recovery using direct plating yielded similar results as enrichment procedures, provided quantitative data, and resulted in the most rapid identification of positive samples. This discrepancy with some other studies might be attributed to different approaches of plating. In total, we used 1 mL of the initial homogenate for direct plating, which corresponds to a theoretical detection limit of 10 yersiniae per gram. Moreover, spread plating and spiral plating may result in a different outcome compared to traditional streaking using a loop. As the former generally leads to numerous well-isolated colonies due to a homogeneous spread of bacteria, isolation of pathogenic yersiniae is less complicated. When applying the direct plating method for isolation of pathogenic Y. enterocolitica from tonsils samples, spread plating or spiral plating is thus preferable over traditional streaking.
The performance of YeCM was equal to the standard agar plates SSDC and CIN. When using short isolation procedures for isolation of pathogenic Y. enterocolitica from pig tonsils, as in this study, the benefit of discriminating pathogenic from non-pathogenic Y. enterocolitica seems to be limited. Nevertheless, YeCM might be beneficial when a large quantity of non-pathogenic yersiniae is expected, and thus when differentiation of pathogenic from non-pathogenic strains is more problematic. For instance, the applied enrichment procedure can influence the detection of non-pathogenic yersiniae, resulting in a higher recovery when prolonged cold enrichment is used compared to selective enrichment and direct plating (de Boer and Nouws, 1991; Kwaga et al., 1990; Laukkanen et al., 2009). Additionally, contrary to pig tonsils, in environmental samples and food products, non-pathogenic strains are predominant, whereas the numbers of pathogenic Y. enterocolitica are low (Fredriksson-Ahomaa and Korkeala, 2003; Kwaga et al., 1990). These conditions complicate isolation of pathogenic strains, and therefore, the use of a selective medium for pathogenic Y. enterocolitica, as for instance YeCM, can be of interest.
About 80% of Y. enterocolitica bioserotype 4/O:3 isolates was PCR-positive for the virF gene. As from many pigs both pYV+and pYV- isolates were recovered, analyzing a limited number of isolates per pig may lead to an underestimation of pYV+pigs. The plasmid is said to be lost easily under in vitro conditions, particularly during repeated culturing in the laboratory and during culturing at 37°C. However, Li et al. (1998) showed that the plasmid is relatively stable when culturing at 32°C, and observed only a slight loss of the plasmid after 9 days of culture in brain-heart infusion broth. In the present study, a considerable proportion of plasmid-negative isolates were detected after direct plating, even though cultures were never incubated at temperatures above 30°C and DNA was extracted no later than 5 days after sampling. Moreover, the proportion of pYV+/pYV- Y. enterocolitica isolates after enrichment was similar to the proportion obtained by direct plating. Thus, enrichment in ITC or PSB at 25°C for 48 h did not seem to cause an apparent selection towards pYV+or pYV- strains. Logue et al. (2006) observed no significant difference in duration of lag phase or growth rate at 25°C between a plasmid bearing and plasmid cured strain, which may suggest that there is no overgrowth by plasmid negative strains during enrichment at 25°C. Hence, it is still unclear in which stage the plasmid is lost.
Conclusion
Sampling methodologies strongly influence the outcome of detection and enumeration of human pathogenic Y. enterocolitica from pig tonsils. Although swab samples are easier and faster compared to destructive samples, the use of homogenization of tissue samples increased the isolation frequency. For direct isolation of pathogenic Y. enterocolitica from destructive tonsil samples, we found spread plating to be a suitable method. Not only does it result in a rapid identification of positive samples, it also provides a measure of bacterial counts. Moreover, we found ISO 10273:2003 with minor modifications to be a rapid and routinely applicable isolation method for the recovery of pathogenic Y. enterocolitica from destructive tonsil samples.
The isolation methods investigated here were only tested for tonsil samples and should not be regarded as valid for isolation of pathogenic Y. enterocolitica from other matrices.
Footnotes
Acknowledgments
Sébastien Morio and Annelies Wachtelaer are acknowledged for their technical assistance. We thank the slaughterhouse personnel for their cooperation in sampling. This research was funded by a doctoral fellowship of Bijzonder Onderzoeksfonds (BOF09/24J/064) of Ghent University.
Disclosure Statement
No competing financial interests exist.