
Abstract
The aim of this study was to identify Salmonella serotypes infecting cattle in Pennsylvania, to compare infection rates for the predominant serotype, Salmonella enterica serotype Cerro, with the infection rates for the same serotype in humans, and to study the clonal diversity and antimicrobial resistance for this serotype in cattle from 2005 to 2010. Clonal diversity among the selected isolates was studied using pulsed-field gel electrophoresis (PFGE) and repetitive (rep)–polymerase chain reaction (PCR). Salmonella Cerro showed the single largest increase as a cause of cattle infections over the study period. The proportional distribution of Salmonella Cerro serotype among laboratory-submitted Salmonella positive cases in cattle was 36.1% in the year 2010 compared to 14.3% in 2005. A simultaneous decrease in serotype Newport infections was also observed in cattle (25% in 2005, to 10.1% in 2010). Studies of clonal diversity for cattle and human isolates revealed a predominant PFGE type but showed some variability. All tested isolates (n=60) were susceptible to sulfamethoxazole-trimethoprim, but 2% of cattle isolates (n=1/50) and 20% of human isolates (n=2/10) showed resistance to tetracycline and sulfisoxazole. One human isolate showed additional resistance to ampicillin and gentamicin. This study suggests an increase in Salmonella Cerro infections in the cattle population and a decrease in Salmonella Newport infections. The increase in Cerro infections appears to be restricted to the cattle population, but occasional human infections occur.
Introduction
S
Salmonella enterica serotype Cerro is infrequently isolated from humans in foodborne infections (CDC, 1985, 2008). The prevalence of this serotype however, in cattle has been on the rise. Cummings et al. (2010) found 46% of dairy herds to be infected with Salmonella Cerro, while recording overall positive rate of 77% for Salmonella in the study herds. A 2007 National Animal Health Monitoring System Dairy study conducted in the United States also recorded a marked (10%) increase in overall Salmonella infections within a 5-year study period (CEAH, 2009).
This study examined non-typhoidal Salmonella serotypes prevalent in cattle to determine the prevalence of Salmonella serotype Cerro, the serotype of interest. The study Salmonellae were isolated from cattle specimens submitted to the veterinary diagnostic laboratory in Central Pennsylvania for animal disease diagnosis. We also compared the incidence of salmonellosis due to Salmonella serotype Cerro in humans to that in cattle, including a study of clonal diversity and antimicrobial resistance for selected isolates in similar temporal and geographic settings.
Materials and Methods
Bacterial isolation and serotyping
Salmonella isolates were recovered from animal tissues or feces submitted to the Pennsylvania Department of Agriculture's Veterinary Laboratory for animal disease diagnosis from dead or clinically ill dairy cattle. Salmonella was isolated by standard culture methods with pre-enrichment using a Gram-negative (GN) enrichment broth, followed by overnight selective tetrathionate broth enrichment (1:10 dilution), and plating onto brilliant green agar with novobiocin and xylose lysine tergitol 4 (XLT-4) selective media (Carter, 1990). Plates were incubated at 37°C for 18–24 h. After biochemical identification and laboratory confirmation of recovered isolates as Salmonella, isolates were serogrouped with antisera (BD, Franklin Lakes, MD). Isolates were subsequently serotyped at the National Veterinary Services Laboratories (Ames, IA), using published methods (Ewing, 1986). Isolated Salmonella from human illness encountered in hospitals and commercial laboratories within the state were forwarded to the Pennsylvania Department of Health's laboratory for serotyping. Serial isolations from the same individual or from a farm with multiple animals were recorded as a single occurrence of a particular serotype for this analysis. For comparing serotype data in cattle at the national level, data were provided by National Veterinary Services Laboratories (Ames, IA).
Resistance to antimicrobials
Cattle (n=50) and human (n=10) Salmonella Cerro isolates were tested against six antimicrobials, including ampicillin 10 μg, cephalothin 30 μg, gentamicin 10 μg, sulfisoxazole 0.25 mg, sulfamethoxazole/trimethoprim 23.75/1.25 μg, and tetracycline 30 μg using disk-diffusion technique following Clinical and Laboratory Standards Institute (CLSI) guidelines. CLSI interpretive values were used to determine breakpoints (CLSI, 2008, 2011).
Pulsed field gel electrophoresis (PFGE)
PFGE was performed on 30 selected Salmonella Cerro cattle isolates according to the Centers for Disease Control and Prevention (CDC) PulseNet protocol described by M'ikanatha et al. (2010). Patterns were analyzed using BioNumerics v. 5.1 software (Applied Maths, Saint-Matins-Latem, Belgium). Cattle isolates for analysis were chosen representing wide geographic areas in the study area based on zip codes and year of isolation. All human isolates (n=10) were typed by PFGE unless re-isolated from the same patient, and data was submitted to the PulseNet database.
Repetitive (rep)–polymerase chain reaction (PCR) typing
Rep-PCR (based on repetitive extragenic palindromic sequence amplification) was used for genomic fingerprinting (Ben-Darif et al., 2010). Total bacterial genomic DNA was isolated from the same cattle (n=30) and human Salmonella Cerro strains (n=10) using the Qiagen DNeasy extraction kit, following the manufacturer's instructions (Qiagen, Germantown, MD). The DiversiLab Salmonella DNA fingerprinting kit (BioMerieux, Duraham, NC) was used for automated rep-PCR molecular typing using AmpliTaq DNA polymerase (Applied Biosystems, Austin, TX) and PCR conditions according to the manufacturer's instructions. The rep-PCR amplicons were separated using an Agilent 2100 Bioanalyzer on the DiversiLab microfluidics DNA chip. DNA fingerprints were analyzed with DiversiLab software using the manufacturer's instructions.
Results
Retrospective analysis of Salmonella serotypes showed changes in serotype prevalence in cattle samples examined at the Pennsylvania Department of Agriculture‘s Veterinary Laboratory from 2005 through 2010, with a shift towards a higher isolation rate for Salmonella Cerro (Table 1a). Percent distribution for the top five serotypes within Salmonella-positive cases was determined based on total number of Salmonella positives isolated in a particular year. In 2005, serotypes Newport and Typhimurium were seen as the dominant cattle serotypes in Pennsylvania, accounting for >50% of Salmonella infections. However, since 2007, Salmonella Newport isolations in cattle have shown a considerable decline (10% in 2010 compared to 29% in 2007; Table 1a). This trend is also reflected nationally among Salmonella serotypes recovered from cattle suffering from clinical disease (Table 1b). Both Pennsylvania and national serotype prevalence data point towards the emergence of Salmonella Dublin in cattle. In 2007, less than 5% of Salmonella isolations reported from the laboratory were due to Salmonella Typhimurium, but this percentage had increased to 25% after 2009 (Table 1a). Recovery of Salmonella Cerro has shown a marked increase paralleling the drop in serotypes Newport and Typhimurium. Table 1a also shows that Salmonella Cerro isolation rate was highest in the year 2008 in Pennsylvania and remained high in 2010. A similar trend for Salmonella Cerro isolation was observed at the national level.
Predominantly from clinical disease cases submitted for disease diagnosis. Salmonella Typhimurium includes variant 5-.
The number of Salmonella isolates (n) recovered and analyzed during the study period.
Data represents isolates submitted to National Veterinary Services Laboratories identified as clinical cases.
The number of Salmonella isolates (n) recovered and analyzed during the study period.
All Salmonella Cerro isolates from cattle (n=50) tested for resistance against six antimicrobials showed in vitro susceptibility to sulfamethoxazole/trimethoprim, gentamicin, ampicillin, and cephalothin, whereas only 2% of isolates were resistant to sulfonamides and tetracycline. Clonality of all 30 selected isolates from cattle when examined with rep-PCR showed less than 2% difference (data not shown). PFGE analysis using the enzyme XbaI was slightly more discriminatory, resulting in six different subtypes, with one profile more dominant than others. Additional bands were observed in several isolates as shown for isolates labeled M14 and M15, pointing to further diversity within the Cerro serotype (Fig. 1). PFGE patterns for cattle isolates showed no clustering with the geographic location (based on zip code), age, or year of isolation as depicted in Figure 1 with the representative set.
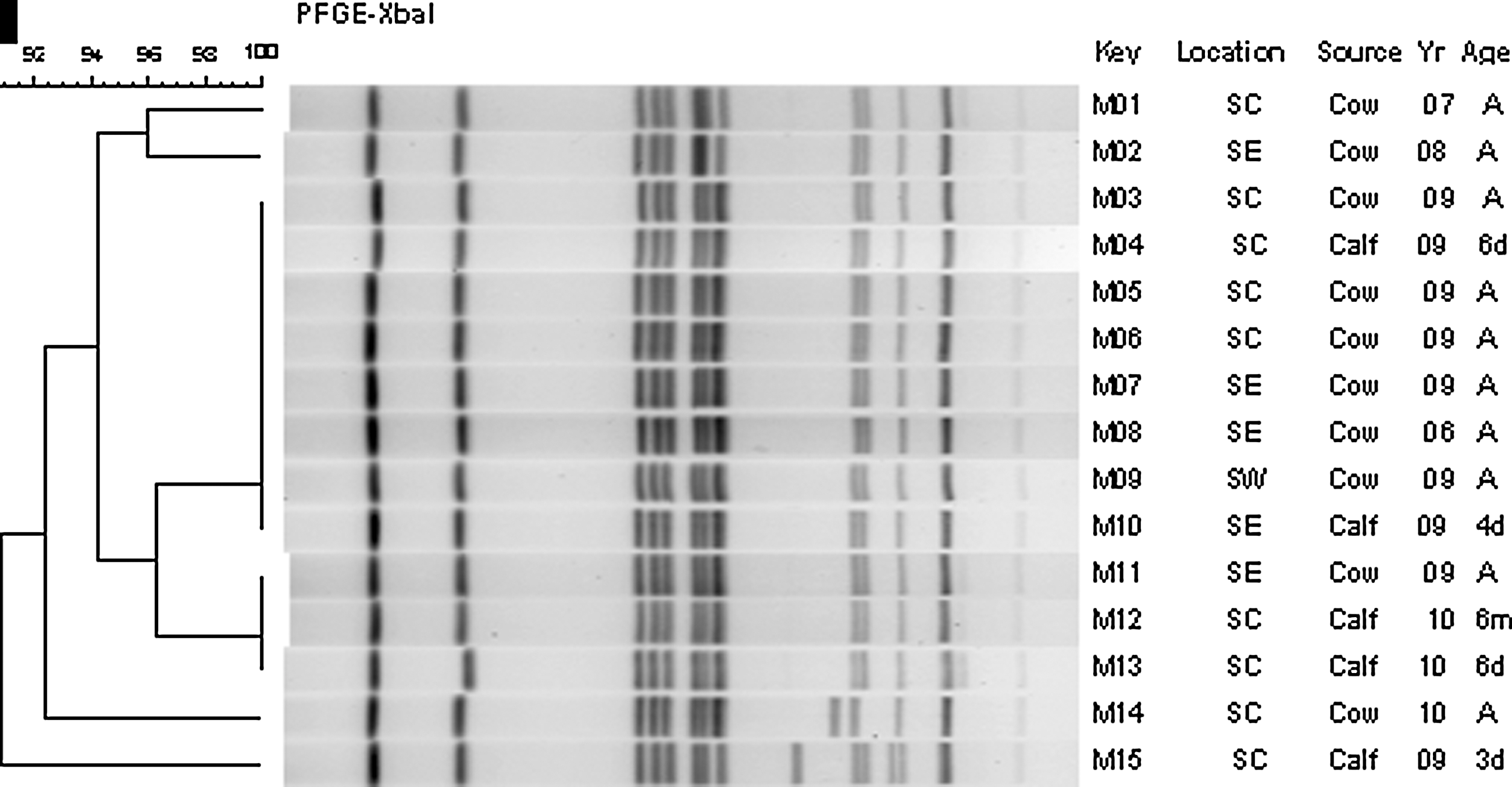
Characterization of Salmonella Cerro isolates from cattle by pulsed-field gel electrophoresis (PFGE) with one-enzyme analysis (XbaI). Dendrogram analysis is on the left, gel image is in the middle, and identity of selected isolates is on the right. Dendrogram analysis also shows percent identity between different nodes. Farm location (shown at region level, based on zip code: SC, south-central; SE, south-east; SW, south-west), source, year of isolation (Yr), and age of animals (A, adult; m, month; d, day) are included with the key.
Serotype Cerro isolates recovered from humans in Pennsylvania during the same period did not show any major increase despite the higher isolation rate observed in cattle (Table 1c). The human isolates examined for antibiotic resistance showed susceptibility to same antimicrobials as observed with cattle isolates except two human isolates that were resistant to tetracycline and sulfonamides. One of these two isolates also showed resistance to ampicillin and gentamicin. The PFGE profile for this isolate was the dominant pattern, JCGX01.0001 (7/10 human isolates showed this pattern), reported in the PulseNet database. This PFGE profile in PulseNet was similar to the pattern exhibited by a number of cattle Salmonella Cerro isolates (e.g., isolates M3–M10; Fig. 1). The other PFGE profiles for human isolates were JCGX01.0002, JCGX01.0031, and JCGX01.0033 (data not shown). Also, the rep-PCR profile for the isolate showing additional antibiotic resistance was indistinguishable (less than 2% different) from that of most other cattle (represented by isolates M03 and M05; Fig. 2) or human isolates (represented by H01). Only one human isolate (H03) showed a distinct rep-PCR profile (Fig. 2), but the PFGE PulseNet profile of this isolate was also the dominant PFGE type (JCGX01.0001), with no additional resistance identified against the tested antibiotics.
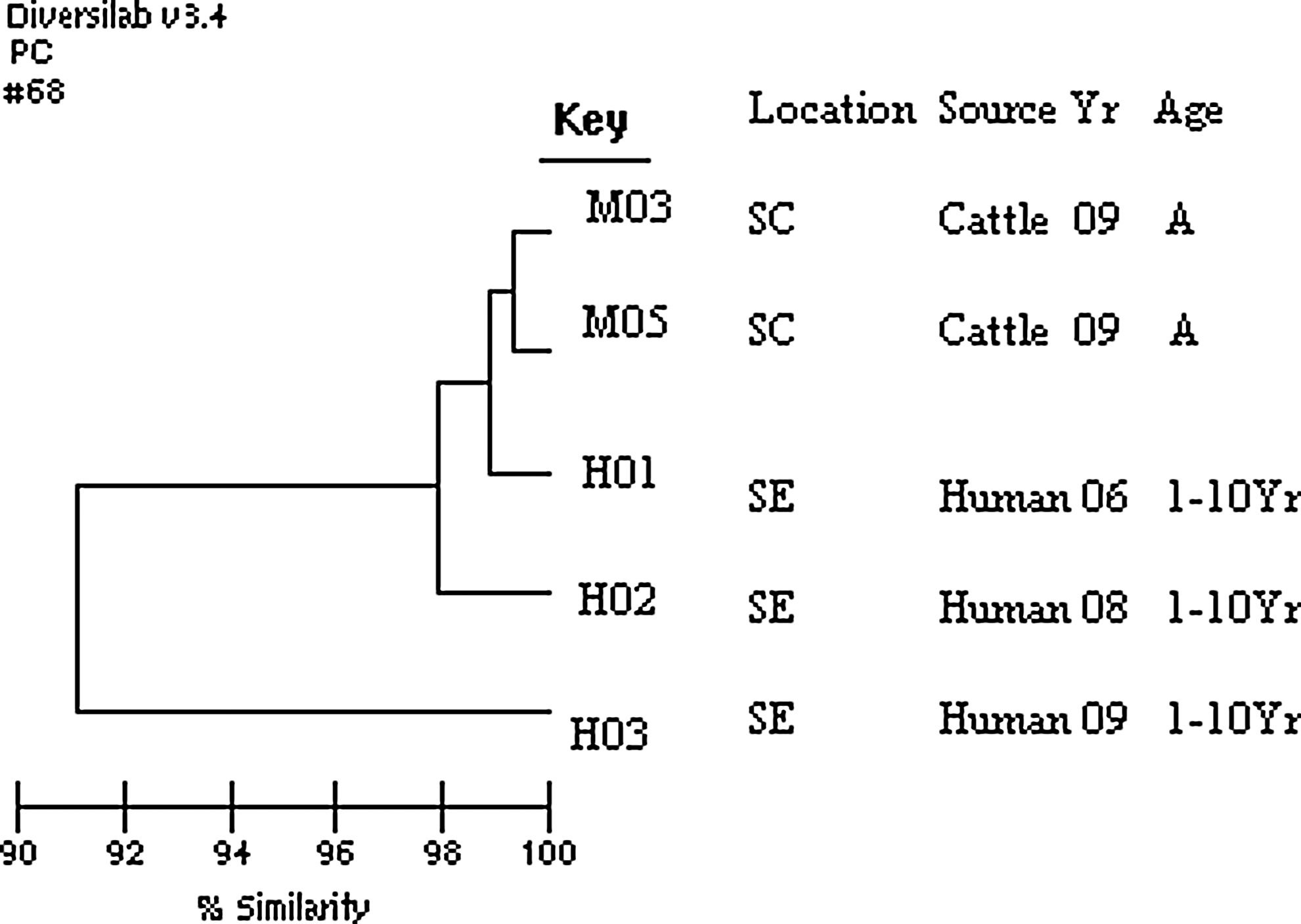
Repetitive (rep)–polymerase chain reaction (PCR) profile of Salmonella serotype Cerro isolates from cattle and humans. Dendrogram relationship among electrophoretic profiles for selected Salmonella serotype Cerro recovered from both cattle and human obtained using Diversilab rep-PCR analysis. Location (shown at region level, based on zip code: SC, south-central; SE, south-east), source, year of isolation (Yr), and age (A, adult; Yr, year) are included with the key. Percent similarity is shown at the bottom of the dendrogram.
The number of Salmonella isolates (n) recovered from patients and analyzed during the study period by the Pennsylvania Department of Health.
Discussion
The study shows a higher recovery rate for Salmonella Cerro in cattle when laboratory-recovered Salmonella isolates were examined. The increase in Salmonella Cerro isolations appears to be restricted to the cattle population with occasional reports of human infection. Analysis of geographic distribution of Salmonella Cerro in cattle showed no clustering by study location and wide distribution based on PFGE analysis. Mammina et al. (2000) discussed infrequent isolations of Salmonella Cerro in the last 30 years from wide-ranging sources, including hen eggs, duck eggs, captive crocodiles, cattle herds, and humans. More recently, emergence of Salmonella Cerro in dairy cattle as an important infection has been described (Van Kessel et al., 2007, 2012; Hoelzer et al., 2011). The CDC reports 447 human cases in the past 30 years, with only four cases between 1968 and 1984. In 1985, a cluster of human salmonellosis cases caused by Salmonella Cerro was identified and associated with consumption of carne seca, an ethnic dried beef product (CDC, 1985). Review of the national PulseNet database shows a few other clusters of Salmonella Cerro PFGE types in humans that are similar to those reported from cattle (CDC, 2010b), but the epidemiologic significance of these matching PFGE types is still unclear.
Besides the higher cattle isolation rate for Salmonella Cerro in Pennsylvania, national data also indicated higher prevalence rate for this serotype. The analysis reported in this study is based on laboratory-submitted diagnostic cases, which did vary from year to year. Variations in the top five serotypes recovered in different years could not have been influenced solely by the laboratory submission patterns. We believe the temporal distribution pattern is representative of the prevalence of serotypes in enteric disease investigations. Subclinical infections with Salmonella Cerro and other Salmonella have been reported on a dairy farm previously (Huston et al., 2002; Fossler et al., 2004; Van Kessel et al., 2007). In a longitudinal study that focused on Salmonella serotypes on a dairy farm, Van Kessel et al. (2007) reported that Salmonella Cerro replaced other prevalent Salmonella serotypes that were responsible for causing clinical disease. The infection with Salmonella Cerro was found to be more of a commensal nature on this dairy farm. The available studies have not fully established the role of Salmonella Cerro in causing clinical disease in cattle because the isolations are also tied to asymptomatic infections (Van Kessel et al., 2007, 2012). Cummings et al. (2010) reported an emerging trend where the likelihood/odds ratio of Salmonella Cerro infection was much higher in diseased animals than in healthy animals. An in-depth investigation of diarrheal diseases in cattle, where Salmonella Cerro was recovered, revealed that on many occasions Clostridium perfringens and other Salmonella, Escherichia coli, and Rotavirus were isolated in addition to Salmonella Cerro. Occasionally, Salmonella Cerro was also recovered from animals that died due to non-infectious causes, pointing further to subclinical or asymptomatic infections.
For the majority of cattle isolates in this study, antimicrobial susceptibility and rep-PCR patterns showed similarities for Salmonella Cerro, even though the isolates were recovered from different farm locations and at different time intervals. One previous study found less than 10% resistance for tested antimicrobials when Salmonella Cerro isolates recovered from cattle were examined (NARMS, 2010). Antimicrobial resistance in one human isolate was higher, but other isolates exhibited resistance patterns similar to the cattle isolates. The clonal nature of the isolates was additionally supported by the finding of one dominant PFGE type. The differences among analyzed isolates with PFGE were found to be minor and only found in a few studied isolates. Van Kessel et al. (2007) reported isolation of indistinguishable isolates from animals on neighboring farms. Hoelzer et al. (2011) also reported a single predominant PFGE type among dairy cattle. Examining the geographic distribution of isolates over the study period suggested that the predominant PFGE type was widely distributed in the study area with no evidence of clustering among the analyzed isolates. Use of additional molecular methods can complement PFGE and rep-PCR, and thus expand our understanding of the epidemiology of these infections.
Interestingly, emergence of the Cerro serotype in cattle appears to have coincided with an observed decrease in Salmonella Newport infections. Salmonella Newport infections, including those involving multi-drug-resistant (MDR) strains, have been associated with severe enteritis in both cattle and humans prior to the emergence of serotype Cerro (Rankin et al., 2002; CDC, 2002). Salmonella serotypes in Pennsylvania recovered from humans, particularly Salmonella Newport (5.1–6.4%) and Cerro (<0.17%), did not show major changes and appear not to be impacted by much higher isolation rates of Salmonella Cerro in cattle. The lower rate of infection in humans despite high isolation rates of Salmonella Cerro from cattle suggests that either cattle are not presently a major source for human infections or such infections may not always result in clinical illness and are thus not captured by public health surveillance.
In humans, Salmonella Typhimurium, Salmonella Entertidis, Salmonella Newport, Salmonella Javiana, and Salmonella Heidelberg remain the most frequently recovered five serotypes (CDC, 2010a). Natural cycling of serotypes in the cattle population or selection pressures due to interventions including vaccination may account for changes in the prevalence of the serotypes. Of note, one of the vaccines used for protecting against Salmonella Newport is a siderophore receptor and porin (SRP) vaccine consisting of extracts from Salmonella Newport. Variable success has been reported with the use of such vaccines (Hermesch et al., 2008; Heider et al., 2008; Dodd et al., 2011). Coincidentally, local and national data suggests that the prevalence of Salmonella Newport has declined considerably in cattle, corresponding with the increase in Salmonella vaccine use. The drop in Salmonella Newport infections, whether due to increased vaccine use or natural cycling of Salmonella serotypes in cattle, has coincided with the higher incidence for Salmonella Cerro infections. It is also possible that some Salmonella vaccines, although stated to be broadly protective, are not effective against all O types. Farzan et al. (2010) reported that Salmonella Typhimurium vaccine use in pigs resulted in the reduced incidence for Salmonella Typhimurium but led to increased shedding of Salmonella Cerro in the vaccinated group. Another possibility is that under selection pressure a virulent serotype is replaced by a less virulent serotype or a serotype that is not impacted by vaccine immunity. To understand the cycling of biotypes, studies need to be conducted using experimental infection models in addition to continuous monitoring of trends or shifts in the Salmonella serotypes in animal and human populations. This will provide a better understanding of the molecular mechanisms involved in competitive exclusion, the effects of vaccines, and pathogenicity specific serotypes in Salmonella infections.
Footnotes
Acknowledgments
We thank Dr. Matthew Erdman and Brenda Morningstar from National Veterinary Services Laboratories, Iowa, for providing serotyping service, sharing national data, and reviewing the manuscript; Ann Ohme, Barbra Porcher, and Julia Livengood for technical assistance; Dr. Helen Acland, Dr. Barbra Corson, and Dr. Mark Walter for allowing access to pathology reports; and Dr. Craig Shultz and Dr. Steve Ostroff for critically reviewing the manuscript.
Disclosure Statement
No competing financial interests exist.