
Abstract
Human anisakidosis is a recognized Arctic zoonosis that is directly related to the consumption of traditional Inuit foods, particularly raw fish. The epidemiology of infections with the zoonotic anisakid nematodes Anisakis simplex and Pseudoterranova decipiens was investigated from August 2007 to July 2009 in Inuit-harvested fish and marine mammals from Inuit regions of Nunavik, Nunavut, and Nunatsiavut, Canada. Fish were tested for anisakid larvae using the pepsin-HCl digestion method, and the stomachs or stomach contents of beluga whales, walruses, and three seal species were examined for anisakids. Anisakids were found in seven of eight fish species, as well as in ringed seals (18.2%; 31/170), bearded seals (75.0%; 12/16), and beluga whales (78.9%; 15/19), but not walruses (0%; 0/15). In fish, the odds of being infected with A. simplex and/or P. decipiens was 68.6 (95% confidence interval, 11.6–627.7) times higher in marine fish than in anadromous fish, after adjusting for length of fish. Negative binomial models were created for animal species with large enough sample sizes and parasite prevalence estimates to assess risk factors associated with anisakid abundance. In seals, the only risk factor significantly associated with increasing anisakid abundance was increasing length (p<0.01), while in beluga whales, the only significant risk factor was year of capture (p=0.03). In fish, length was the variable most commonly associated with increased anisakid larval abundance, with longer fish having significantly higher larval abundances than shorter fish of the same species. The presence of A. simplex and P. decipiens in bearded seals, ringed seals, and beluga whales from Inuit hunting grounds suggests that they likely act as definitive hosts for these parasites in these environments. With respect to zoonotic disease risk associated with Inuit country foods, among the species of fish examined, Atlantic tomcod, polar cod, and sculpins represented the greatest risk of foodborne disease from A. simplex and P. decipiens.
Introduction
A
Humans are accidental hosts in the life cycle (Sakanari and McKerrow, 1989), typically ingesting L3 larvae while consuming raw, insufficiently cooked, smoked, or marinated flesh from infected fish (JongYil et al., 2005). As such, only L3 larvae are of public health significance in humans. Anisakidosis presents in two forms: non-invasive and invasive. Non-invasive anisakidosis occurs when the parasite does not penetrate the gastrointestinal mucosa. This form is often asymptomatic, although it can be associated with “tingling throat syndrome,” wherein anisakids migrate up the esophagus and are coughed out by the patient. Invasive anisakidosis occurs when L3 larvae embed themselves into the mucosa of the stomach or intestine. Within a week after consuming larvae, patients may experience nausea, vomiting, diarrhea, and/or abdominal pain (Sakanari and McKerrow, 1989). In chronic cases, nausea and vomiting may last up to 2 years (Ko, 2006). Clinical signs and symptoms of anisakidosis include edema in the gastric mucosa, epigastric pain, vascular occlusion, diffuse abdominal pain that mimics a gastric ulcer, and pseudo tumoral formation (Bouree et al., 1995; Takabe et al., 1998). In rare instances, an acute allergic response associated with ingestion of larvae or physical contact with an infected fish can cause a fatal anaphylactic reaction (Audicana et al., 2002).
Seroprevalence studies of anisakid infection in human populations are uncommon, with only two published peer-reviewed reports (Møller et al., 2007; Kim et al., 2011). Based on data from case reports, anisakidosis occurs more frequently in areas where cultural practices involve eating raw fish (Sakanari and McKerrow, 1989). Japan, where raw fish is widely consumed, has the highest incidence of anisakidosis, with approximately 2000 cases diagnosed annually (Ko, 2006). In Canada, a case was reported in a 50-year-old Quebec man after he consumed raw Pacific salmon (Couture et al., 2003), and in a 40-year-old woman, also from northern Quebec, who became ill after eating raw Arctic charr (Bhat and Cleland, 2010). Sporadic cases have been reported in the United Kingdom, continental Europe, Iceland, Chile, Australia, and New Zealand (Sakanari and McKerrow, 1989; Skírnisson, 2006; Ugenti et al, 2007; Shamsi and Butcher, 2011).
Anisakids are zoonotic pathogens of interest in the Arctic and subarctic as fish are a prominent part of the Inuit diet (Butler Walker et al., 2006; Johnson et al., 2009) and are traditionally consumed raw, or prepared in ways that do not kill the larvae, such as smoking or light salting (McDonald et al., 1990; Treshchev and Serdyukov, 1997; Food Safety Network, 2009). Despite this, little data exist on the prevalence of, and risk factors for, anisakids in fish and marine mammals from Canadian waters, particularly among stocks used as food sources by Inuit.
The objectives of the present study were to investigate the prevalence of A. simplex and P. decipiens in eight fish species, beluga whales, walruses, and three seal species that are part of the Inuit diet in the eastern Canadian Arctic, and to determine potential risk factors for anisakid abundance in these animals. Anisakid abundance in marine mammals was investigated because, as potential definitive hosts of A. simplex and P. decipiens, they play an important role in the transmission of these parasites to fish used as food sources by Inuit.
Materials and Methods
Sample collection
Animals to be tested for anisakids were determined in conjunction with community collaborators from the Nunavik Hunters Fishers and Trappers Association and Makivik Corporation in Kuujjuaq, Nunavik, Canada, and local representatives from Sikumiut Environmental Management Ltd. and hunters from Nain, Nunatsiavut, Canada. Consultations were used to identify the animal species commonly consumed in these Inuit communities. The resulting list served as the basis for the sampling scheme.
Samples (i.e., marine mammal stomach/stomach contents and whole fish) were collected from August 2007 to July 2009. Approximately 40 hunters were trained in sample collection techniques, provided with sampling kits and asked to record on standardized forms where the animal was caught and, if it was a marine mammal, its sex and species. Hunters froze all fish as soon as possible after capture, but were not asked to record the time between capture and freezing. Hunters were provided with financial compensation for submitted samples. The study was approved by the University of Guelph Animal Care Committee and the Nunatsiavut Government Research Advisory Committee.
In total, eight fish species, three seal species, beluga whales, and walruses were examined (Table 1). Samples were either brought fresh to the laboratories in Nain or Kuujjuaq by hunters, or shipped frozen. Upon receipt by either laboratory, all samples were stored at −20°C prior to analysis.
Laboratory methods
Prior to dissection, fish were weighed, sexed, measured for length, and the otoliths or scales collected for age determination. Age determination was conducted by a Department of Fisheries and Oceans (DFO) biologist; salmon and whitefish were aged using scale readings (Anonymous, 1984); all other fish were aged from otoliths using the break-and-burn method (Das, 1994; Cailliet et al., 2001). Fish were dissected and the visceral organs and coelomic cavity macroscopically examined for anisakid larvae (McGladdery, 1986). In all species, a 20-g sample of muscle tissue was taken from the ventral side of each fish, from which anisakids were collected using the pepsin-HCl digestion method (Lunestad, 2003). All anisakid larvae collected by both detection methods were counted and combined for analysis. Detection in seals was carried out by macroscopic examination of stomach contents for anisakids, as well as examination of the stomach wall for embedded anisakids. All adult and larval anisakids were counted and collected. Due to lack of freezer space and shipping costs, only 500 mL of stomach contents (solid and liquid phases) were collected from beluga whales and walruses. Beluga whales were aged by counting growth rings in cementum annuli in teeth (Morris, 1972) at the Nunavik Research Centre, Makivik Corporation, Kuujjuaq, Nunavik, QC, Canada, and at Matson's Laboratory, Milltown, Montana. Walrus were aged by classes (pup, juvenile, adult) given by the Nunavik walrus hunters.
All collected anisakids were immediately placed in a solution of physiological saline (154 mol/L) and examined under a light microscope at 100× magnification. Any encapsulating tissue was removed, and genus identification was conducted based on morphology (Oshima, 1972; Fagerholm, 1982; Berland, 1989). All anisakids were then placed in 70% ethanol in water for fixation, storage, and shipping. Species identification was performed on all anisakids using polymerase chain reaction-restriction fragment length polymorphism (RFLP); species-specific DNA fragments were sequenced and compared to previously identified RFLP sequences deposited in GenBank (Kijewska et al., 2009).
Statistical analysis
Data for A. simplex and P. decipiens were analyzed together as this research was approached from a human health perspective; both are zoonotic parasites. Descriptive statistics were calculated for each host species and used to estimate mean abundance, burden range, and prevalence of L3 larvae (fish), or L3, L4 larvae, and adults (marine mammals). For fish, larvae detected in the coelomic cavity and muscle tissue were combined for analysis as we did not know when fish had been frozen and therefore did not know in which fish larvae could have migrated into the muscle tissue. Any data presented on anisakid location in fish would therefore be unlikely to be a true representation of the likelihood of finding larvae in the muscle tissue. Adult and larval anisakids from marine mammals were not differentiated for analysis because a future publication will address larval stages. Mean abundance was defined as the mean number of A. simplex and P. decipiens in each host species, including the zero values of uninfected hosts. Burden range was defined as the minimum (excluding zero counts) and maximum number of A. simplex and P. decipiens, combined, in each host species. Prevalence was defined as the percentage of all hosts in a species that had at least one A. simplex or P. decipiens.
A multivariable exact conditional logistic regression model was used to compare the odds of anisakid infection in anadromous fish (Arctic charr [Salvelinus alpines], Atlantic salmon [Salmo salar], Atlantic whitefish [Coregonus huntsman], and brook trout [Salvelinus fontinalis]) versus marine fish (Atlantic tomcod [Microgadus tomcod], polar cod [Boreogadus saida], and sculpins). To assess factors impacting parasite burden in different species, negative binomial models of the combined counts of A. simplex and P. decipiens were built for each of the following species: Arctic charr, Atlantic tomcod, polar cod, ringed seal (Pusa hispida), and beluga whale (Delphinapterus leucas). Due to small sample sizes and similar biology, diet, and sample locations, data from the two sculpin species (longhorn sculpin [Myoxocephalus octodecemspinosus] and shorthorn sculpin [Myoxocephalus scorpius]) were combined and modeled together.
All multivariable models (logistic and negative binomial) were initially constructed using all variables that were unconditionally associated with the outcome at p≤0.20 in univariable modeling. The final multivariable models were determined by using a manual backwards elimination procedure, with p-values of ≤0.05 as the cut-off for inclusion. The potential clustering effect of location was addressed by including it as a fixed effect in the models where necessary (Dohoo et al., 2003). Collinearity was assessed between all variables using the variance inflation factor (VIF); if two variables were highly correlated (VIF>10), then the variable deemed less biologically relevant was dropped (Dohoo et al., 2003). The effect of potential confounding variables was evaluated by individually removing them from the model and determining whether there was at least a 30% change in the coefficients of the remaining significant predictor variables. Linearity of continuous predictors was assessed using a Lowess curve of the continuous variable versus the natural logarithm of the predicted rate. Two-way interactions among all predictors in the final model were tested for significance (p≤0.05) using the likelihood ratio test. Standard residual analyses were performed to assess model fit (Dohoo et al., 2003). All statistical analyses were conducted using STATA/IC 10.1 (StataCorp LP, 2008).
Results
All analyzed Anisakis were found to be A. simplex, and all Pseudoterranova were P. decipiens. Approximately 31.5% of anisakids (1536/4871) could not be speciated due to DNA degradation, which was likely due to freezing and thawing, preservation in saline solution during microscopic examination, and storage in 70% ethanol. Based on the polymerase chain reaction findings, the fact that typable parasites came from all the species sampled (with the exception of Atlantic whitefish, among which only one parasite was found), and the current known geographic distribution of Anisakis and Pseudoterranova species (Kuhn et al., 2011), the speciation results were extrapolated to all isolated anisakids, so that all Anisakis and Pseudoterranova collected were assumed to be A. simplex and P. decipiens, respectively.
Prevalence, mean abundance, and burden range of A. simplex and P. decipiens in tested fish and marine mammals are shown in Tables 2 and 3, respectively. Variables tested in all models and their univariable associations with anisakid abundance are listed in Table 4.
CI, confidence interval; SD, standard deviation; NA, not available.
CI, confidence interval; SD, standard deviation; NA, not available.
Age data were not collected for Atlantic tomcod due to logistical problems and therefore not included in the analysis.
Weight was dropped from further analyses due to high correlation with length.
Significant univariable associations (p≤0.05) are indicated in bold.
In fish, the odds of being infected with A. simplex and/or P. decipiens was 68.6 (95% confidence interval [CI], 11.6–627.7) times higher in marine fish (Atlantic tomcod, polar cod, and sculpins) than in anadromous fish (Arctic charr, Atlantic salmon, Atlantic whitefish, and brook trout), after adjusting for length of fish (odds ratio [OR], 2.6; 95% CI, 1.7–4.2).
Due to the absence of anisakids in brook trout, harbor seals, and walruses, the small sample size for bearded seals, and the low prevalence of anisakids in Atlantic salmon and whitefish, no negative binomial models were created for these species. Results from the final negative binomial models for parasite risk factor analyses in the remaining fish species are shown in Table 5. Interaction terms are illustrated in Figures 1 and 2.
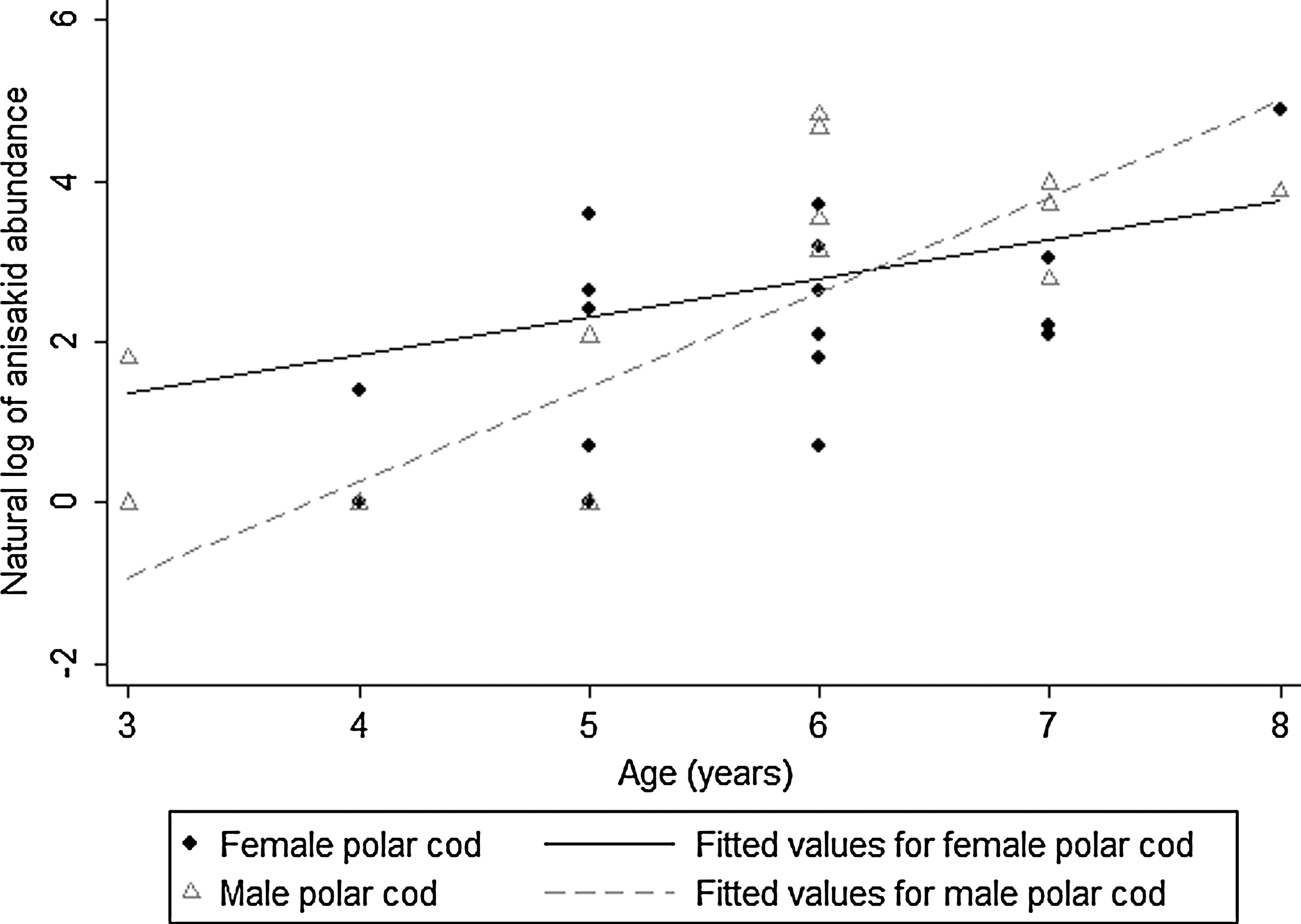
Effect of interaction between age and sex in polar cod on the natural log of anisakid abundance, as derived from a negative binomial model. A 1-year increment in age increased the abundance of anisakid larvae by 1.61 times in female fish, whereas in male fish, it increased larval abundance by 3.27 times.

Effect of interaction between sculpin species and length on the natural log of anisakid abundance, as derived from a negative binomial model. In longhorn sculpin, the abundance of anisakid larvae increased by 1.05 times for every centimeter increase in length, while in shorthorn sculpin, larval abundance increased by 1.46 times with each centimeter increase in length.
Included in final model to account for clustering in data.
CI, confidence interval.
Beluga whales had the highest anisakid prevalence (78.9%, 15/19) of all animals tested in this study. In beluga whales of the same age, the only factor found to be significantly associated with parasite abundance was year of capture. Ten beluga whales were caught in 2008 compared to nine in 2009; the same sampling protocol was used in both years. A whale caught in 2009 was 3.28 times (95% CI, 1.10–9.76) more heavily parasitized than a whale caught in 2008. In ringed seals, no P. decipiens was detected, and the only risk factor significantly associated with A. simplex abundance after model construction was length; for every centimeter increase in seal length, A. simplex abundance increased by 1.08 times (95% CI, 1.05–1.11).
Discussion
From an evaluation of published literature, this appears to be the first comprehensive study of zoonotic anisakid nematodes in Inuit-harvested foods. Of the eight fish species tested, seven had at least one fish positive for A. simplex or P. decipiens. Although all but one fish species were infected with anisakids, there were marked differences in prevalence after adjusting for location. Anadromous fish had significantly lower odds of anisakid infection (p<0.01) than other non-anadromous fish, likely due to their more limited time spent in the marine environment; anisakids are found in saline and brackish waters (Køie et al., 1995).
Few studies have examined the prevalence of anisakids in the fish species discussed here, instead they have focused on more commercially important species such as Atlantic cod (Brattey and Bishop, 1992; Aspholm et al., 1995; Perdiguero-Alonso et al., 2008). However, Hicks and Threlfall (1973) examined anisakids in trout and Arctic charr from Labrador, Canada. The prevalence of A. simplex and P. decipiens in brook trout (0%) and Arctic charr (18.2%) observed here is similar to prevalence values observed in 1969 off the Labrador coast, where 4% (5/124) of brook trout and 8.6% (4/35) of Arctic charr contained Anisakis spp. (Hicks and Threlfall, 1973).
We investigated the occurrence of anisakid nematodes in marine mammals because high anisakid prevalence in definitive hosts can potentially facilitate increased prevalence in fish. Due to the fact that only one harbor seal was captured for this study, it is not possible to comment on their role in anisakid infection in north-eastern Canada. Similarly, because so few beluga whales were tested in this study, direct comparisons to earlier work are not possible. However, two other studies examined beluga whales for anisakids in Canada and found anisakids in 74% (38/51) of beluga whales from the St. Lawrence Estuary (1984–1993) and in 20% (2/10) of beluga whales from the Northwest Territories (1984) (Measures et al., 1995; Wazura et al., 1986).
Seals from the current study had relatively high prevalence and abundance values, particularly when compared to previous studies. In ringed seals, the prevalence and abundance of A. simplex (18.2% and 0.3±1.4, respectively) were higher than 1985–1993 values (7.1% and 0.18±0.11, respectively) in ringed seals from Newfoundland and Labrador (Brattey and Stenson, 1993). Comparing the same studies, bearded seals in the present study also had higher prevalence and mean abundance values (75% and 189.5±126.4, respectively), than those reported by Brattey and Stenson (1993) (55.8% and 41.6±12.5, respectively). The strength of the modeling in the current work was impacted by the small sample sizes for several animal species. This limitation was defined by what was caught by hunters for food within the 2-year sampling period of the study. Despite the small sample sizes, the presence of adult anisakids embedded in the stomachs of seals and beluga whales in this study provide evidence to suggest that these marine mammals act as definitive hosts in Canadian waters.
Documented anisakid infection in humans is rare in the North American Arctic and subarctic, with only nine confirmed clinical cases reported since 1982 (Food Safety Network, 2009). The true extent of human infection is, however, likely to be underreported due to non-specific symptoms, frequently mild clinical presentation, misdiagnosis of severe cases as appendicitis, Crohn's disease, or other gastric ailments (Sakanari and McKerrow, 1989), poor access to medical facilities in many northern regions, and an aversion among Inuit to visiting a physician or hospital (McDermott, 2002; McDonald and Trenholm, 2010). Anisakidosis cases that have been reported in the Arctic are typically associated with the consumption of raw fish (Couture et al., 2003; Food Safety Network, 2009; Bhat and Cleland, 2010). Despite the low frequency of reporting, anisakidosis is recognized as an important Arctic zoonosis by the European Food Safety Authority (EFSA BIOHAZ, 2010) and is directly related to the consumption of traditional country foods (Lymbery and Cheah, 2008). It is therefore of great importance that the monitoring of traditional Inuit food sources for anisakids continues and that the knowledge arising from this work be shared with appropriate public health and community groups in order to allow Inuit to make informed choices regarding their food consumption.
Conclusion
The present study isolated A. simplex and P. decipiens from seven of eight fish species harvested by Inuit in Nunavik and Nunatsiavut. Anisakids were also found in beluga whales and bearded and ringed seals, but not walruses. The presence of A. simplex in seals, and A. simplex and P. decipiens in beluga whales from Inuit hunting grounds suggests that they may act as definitive hosts for these parasites. With respect to zoonotic disease risk associated with Inuit country foods, among the examined fish species, anadromous fish had significantly lower odds of infection with anisakids than marine fish. Overall, Atlantic tomcod, polar cod, and sculpins represented the greatest odds of foodborne disease from A. simplex and P. decipiens.
Footnotes
Acknowledgments
We wish to thank the community collaborators who helped determine the sample list; hunters who collected samples; the Hunter Fisher Trapper Association in Nunavik; laboratory technicians Nancy Sillit (Nain), Sandy Suppa, and Jeannie Gordon (Kuujjuaq); Tara Bortoluzzi, Peter May, and Becky Sjare for seal sub-sampling; John Babaluk for fish aging; William Sears for statistical assistance; and Matt Lehoux for technological assistance. Research funds were provided by the International Polar Year (IPY). Funding for E.P. was provided by the Ontario Veterinary College M.Sc. Fellowship/New Faculty Advisor Award and IPY.
Disclosure Statement
No competing financial interests exist.