
Abstract
The aim of this study was to investigate the prevalence of plasmid-mediated quinolone resistance (PMQR) determinants in Escherichia coli isolated from food-producing animals and to characterize the PMQR-positive isolates. A total of 365 E. coli isolates which were either nalidixic acid resistant and ciprofloxacin susceptible (NALR-CIPS; n=185), or nalidixic acid and ciprofloxacin resistant (NALR-CIPR; n=180) were assessed for the presence of PMQR determinants by polymerase chain reaction. PMQR-positive isolates were further characterized by mutation analysis within the quinolone resistance-determining region (QRDR) of gyrA, gyrB, parC, and parE, phylogenetic group analysis, and pulsed-field gel electrophoresis (PFGE). Fourteen NALR-CIPS (n=8) and NALR-CIPR (n=6) E. coli isolates were positive for PMQR genes. Among them, qnrB4, qnrS1, and aac(6’)-Ib-cr genes were detected in two (0.5%), eight (2.2%), and four (1.1%) isolates, respectively. None of the isolates harbored qnrA, qnrC, qnrD, and qepA genes. All but one PMQR-positive isolates harbored one or more point mutations in the QRDR of gyrA, and five of these isolates had additional mutations in the parC gene. Furthermore, one isolate each had additional substitutions in gyrB and parE genes, respectively. The most prevalent mutation was Ser83-Leu within the QRDR of gyrA. Phylogenetic analysis identified three major phylogenetic lineages, with phylogroups A (n=7) and D (n=4) being the most common phylogroups. None of the isolates belonged to virulent phylogroup B2. PFGE demonstrated that a combination of clonal and horizontal gene transmission is disseminating PMQR genes among the veterinary E. coli isolates in Korea. To our knowledge, this is the first report of occurrence of qnrB, qnrS, and aac(6’)-Ib-cr genes in E. coli isolated from food-producing animals in Korea. Isolation of PMQR genes from food animals is a matter of concern since they could be transmitted to humans via food animals.
Introduction
Besides conferring a low-level resistance to (fluoro)quinolones, PMQR genes may also facilitate selection of additional chromosomal resistance mechanisms, leading to the emergence of bacterial strains with a high level of (fluoro)quinolone resistance (Strahilevitz et al., 2009; Poirel et al., 2008). Moreover, reduced susceptibility to fluoroquinolones in Enterobacteriaceae is an important cause of clinical treatment failures in humans which cause significant therapeutic problems worldwide (Aarestrup et al., 2003; Molbak et al., 1999; Strahilevitz et al., 2009). Furthermore, PMQR genes are usually carried on mobile genetic elements and the transferability of resistant bacteria or mobile resistance genes between animals and humans via the food chain or direct contact has been well documented (Guardabassi et al., 2004; Simjee et al., 2002; Molbak et al., 1999). Thus, resistance to (fluoro)quinolones due to PMQR determinants is a matter of concern both in human and veterinary medicine.
During the past decade, the PMQR gene alone or in association with plasmid-mediated extended-spectrum β-lactamase (ESBL) or AmpC β-lactamase have been increasingly reported among the clinical isolates of Enterobacteriaceae from humans in Korea (Tamang et al., 2008; Kang et al., 2009; Kim et al., 2009). However, data on the occurrence of PMQR determinants among bacteria from animals in Korea is very rare (Tamang et al., 2011). Therefore, the objective of the present study was to determine the prevalence of PMQR genes in E. coli strains isolated from food-producing animals during 2003–2011 and to characterize the PMQR determinant-positive E. coli isolates.
Materials and methods
Bacterial strains
A total of 365 nalidixic acid resistant and ciprofloxacin susceptible (NALR-CIPS; n=185) and nalidixic acid and ciprofloxacin resistant (NALR-CIPR; n=180) Escherichia coli isolated from food-producing animals in Korea were investigated. Among them, 187 isolates were obtained from diseased cattle (n=67) and diseased pigs (n=120). These E. coli from sick animals were isolated from clinical samples at Diagnostic Laboratory of Animal, Plant, and Fisheries Quarantine and Inspection Agency (QIA), Anyang, Korea during 2003–2009 or received from 16 local veterinary service centers during 2010–2011. Remaining 178 isolates examined were obtained from healthy cattle (n=42) and healthy pigs (n=136). These commensal E. coli were isolated from fecal samples collected from various farms located throughout Korea. The collection of samples, culture, and isolation of E. coli isolates were done as described previously (Lim et al., 2007). During strain selection, 10 E. coli isolates (one E. coli per fecal sample) per farm were randomly selected for initial screening of quinolone resistance and one E. coli isolate per farm was randomly selected for further analysis. Identification of E. coli isolates was done by standard biochemical tests and confirmed by the Vitek system (BioMerieux, Hazelwood, MO).
Detection of PMQR genes
The qnrA, qnrB, qnrC, qnrD, qnrS, aac(6’)-Ib, and qepA genes were detected by two sets of multiplex polymerase chain reactions (PCRs) using primers (Table 1) and conditions as described previously (Jacoby et al., 2009; Kim et al., 2009; Park et al., 2006; Tamang et al., 2011). All aac(6’)-Ib positive PCR amplicons were further subjected to digestion with BtsCI (New England Biolabs, Ipswich, MA) restriction enzyme to identify aac(6’)-Ib-cr, which does not have the BtsCI restriction site present in the wild-type gene as described previously (Park et al., 2006). For amplification of the entire qnr genes, qnrB- or qnrS-positive isolates were reamplified and sequenced using qnrB and qnrS full-length primers as described previously (Tamang et al., 2008; Hopkins et al., 2007).
Antimicrobial susceptibility testing
Antimicrobial susceptibility test was performed by standard disc diffusion method according to the guidelines of Clinical Laboratory Standards Institute (CLSI, 2009) using Mueller Hinton agar (Becton-Dickinson, Sparks, MD). The antimicrobial drugs tested were ampicillin, amoxicillin-clavulanic acid, apramicin, cephalothin, ceftiofur, cefoxitin, colistin, chloramphenicol, florfenicol, gentamicin, neomycin, streptomycin, tetracycline, and trimethoprim-sulfamethoxazole (Becton-Dickinson). Minimum inhibitory concentrations (MICs) of nalidixic acid, oxolinic acid, ciprofloxacin, enrofloxacin, norfloxacin, ofloxacin, and danofloxacin (Sigma Chemical Co., St. Louis, MO) were determined for all the PMQR-positive isolates following the CLSI guidelines (CLSI, 2009). Escherichia coli ATCC 25922 was used as the quality control strain.
Sequencing of gyrA, gyrB, parC, and parE
Quinolone resistance-determining regions (QRDRs) of gyrA, gyrB, and parC genes in PMQR gene-positive isolates were amplified using previously described primers and protocols (Giraud et al., 1999; Everett et al., 1996). The primer set for amplification of QRDR of parE was designed using the parE nucleotide sequence reported in the GenBank. Purified PCR products were sequenced with specific primers using an automated ABI Prism 3700 Analyzer (Applied Biosystems, Foster City, CA). The QRDR DNA sequences of gyrA, gyrB, parC, and parE for each of the isolates examined were compared with those of E. coli strain K-12. Analysis and comparison were performed with BLAST program at the National Center for Biotechnology Information (
Phylogenetic grouping
The major phylogenetic group of the E. coli strains was determined by multiplex PCR based on the presence of chuA, yjaA, and TSpE4, as described previously (Clermont et al., 2000).
Pulsed-field gel electrophoresis (PFGE)
PFGE of XbaI (Takara Bio Inc., Shiga, Japan) digested genomic DNA was carried out according to the Pulse Net Standardized Laboratory Protocol as described previously (Gautom, 1997) using a CHEF MAPPER apparatus (Bio-Rad, Laboratories, Hercules, CA). The similarities of the restriction fragment length polymorphisms were analyzed using Bionumerics software, version 4.0 (Applied Maths, Sint-Martens-Latem, Belgium) to produce a dendogram. Clustering was carried out by the unweighted pair-group method with arithmetic averages (UPGMA), based on the Dice similarity index.
Results
Prevalence of PMQR determinants
A total of 14 E. coli isolates representing 3.8% of the 365 E. coli isolates investigated were positive for PMQR genes. Among them, 10 were from samples from sick animals submitted for diagnostic investigation, and four were commensal isolates from healthy animals. Over all, qnrB, qnrS, and aac(6’)-Ib genes were detected in two (0.5%), eight (2.2%), and 10 (2.7%) isolates, respectively. Of the 10 aac(6’)-Ib genes detected, four (1.1%) were aac(6’)-Ib-cr. None of the isolates harbored qnrA, qnrC, qnrD, and qepA genes. The PMQR determinants were detected as early as 2005. All qnrB and qnrS determinants belonged to the qnrB4 and qnrS1 alleles, respectively, on subsequent sequencing. The prevalence of PMQR genes was slightly higher in NALR-CIPS isolates (4.3%) compared to that in NALR-CIPR isolates (3.3%), whereas prevalence of PMQR genes was higher in cattle (7.3%) than in pigs (2.3%). The occurence of various PMQR genes in NALR-CIPS and NALR-CIPR swine and cattle E. coli isolates is compared in Table 2.
PMQR, plasmid-mediated quinolone resistance; NALR, nalidixic acid resistant; CIPS, ciprofloxacin susceptible; and CIPR, ciprofloxacin resistant.
Antimicrobial resistance phenotype
The MICs for nalidixic acid, oxolinic acid, ciprofloxacin, enrofloxacin, norfloxacin, ofloxacin, and danofloxacin against PMQR-positive E. coli isolates are shown in Table 3. Of them, 2.7% (10/365) and 0.8% (3/365) of the isolates exhibited high level resistance to nalidixic acid (MIC≥256 mg/L) and ciprofloxacin (MIC≥64 mg/L), respectively. All PMQR-positive isolates were co-resistant to at least four additional classes of antimicrobial agents other than quinolones and fluroquinolones. The resistance patterns of antimicrobials other than quinolones are shown in Figure 1.
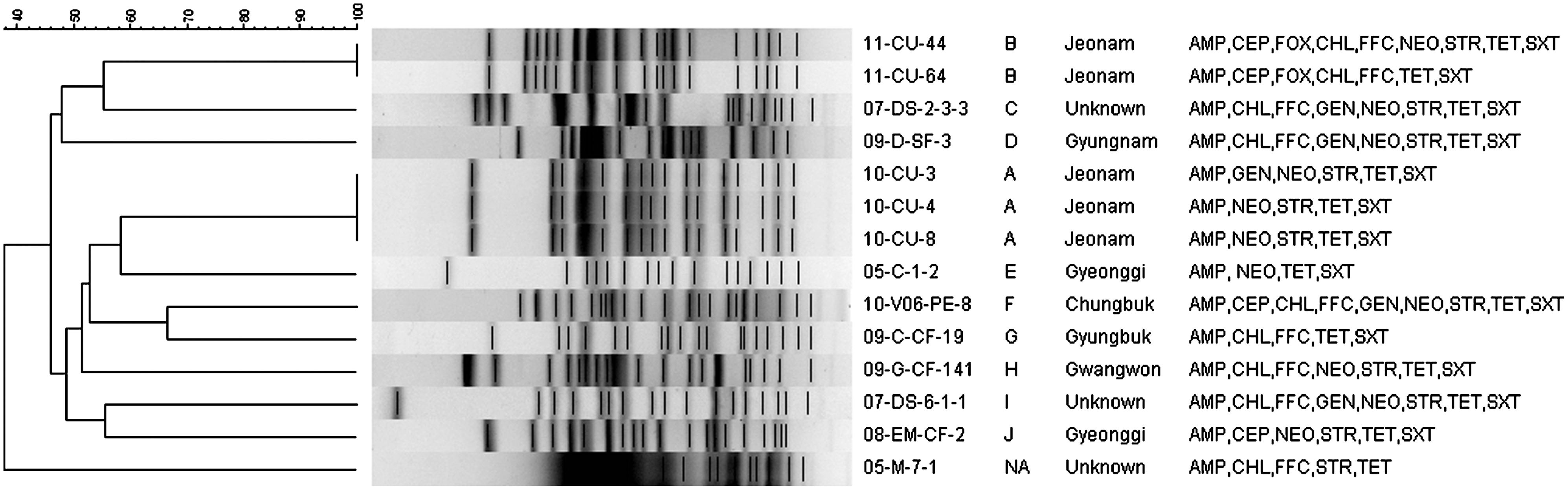
Dendogram generated by Bionumerics software showing the cluster analysis of XbaI- PFGE patterns of PMQR-positive Escherichia coli strains isolated from food-producing animals. Similarity analysis was performed by using the Dice coefficient, and clustering was done by the unweighted-pair group method with arithmetic averages (UGPMA). Details given include the strain, pulso-type of each strain, geographical origin of host animal, and resistance pattern of antimicrobials other than quinolones shown by each strain. XbaI- macrorestriction yielded a few DNA banding patterns in an E. coli strain 05-M-7-1 due to the autodegradation of the genomic DNA during agarose plug preparation and hence a cluster formed by this strain is ignored. NA, not applicable; AMP, ampicillin; CEP, cephalothin; FOX, cefoxitin; CHL, chloramfenicol; FFC, florfenicol; NEO, neomycin; GEN, gentamicin; STR, streptomycin; TET, tetracycline; SXT, trimethoprim-sulfamethoxazole.
PMQR, plasmid-mediated quinolone resistance; NAL, nalidixic acid; OXA, oxolinic acid; CIP, ciprofloxacin, ENR, enrofloxacin; NOR, norfloxacin; OFX, ofloxacin; DAN, danofloxacin; WT, wild-type gene; QRDR, quinolone resistance-determining region.
Mutations in the topoisomerase genes
All but one PMQR-positive isolates harbored one or more point mutations in the QRDR of topoisomerase genes (Table 3). Within the QRDR of gyrA, they showed single amino acid substitutions at codon Ser83 to Leu (n=5) or Ala (I=2) or at codon Asp87 to Tyr (n=1) as well as double mutations at codons Ser83 to Leu and Asp87 to Asn (n=3). Besides mutations within gyrA, five isolates revealed additional mutations in the parC: Ser80 to Arg (n=2) or Ile (n=1); Ser80 to Ile and Glu84 to Gly (n=1); and Ala56 to Thr and Ser80 to Ile (n=1). In addition, one isolate each had additional substitution in parE (Ser458-Ala) and gyrB (Ser492-Asn) genes, respectively. There were no strains with a gyrB, parC, or parE QRDR mutation alone. Overall, the most prevalent mutation in the QRDR was Ser83-Leu.
Phylogenetic grouping of PMQR-positive isolates
Three major phylogenetic lineages (A, B1, and D) were found among the isolates harboring PMQR genes. Phlygroups A (n=7) and D (n=4) were the most common (Table 3). None of the isolates belonged to virulent phylogroup B2.
PFGE analysis
In order to compare genetic relatedness of PMQR-positive E. coli strains, molecular typing was done by PFGE of XbaI-digested genomic DNA. Among them, at least 10 arbitrary pulso-types (designated as A through J) were observed (Fig. 1). While Pulso-type A (3 strains) and Pulso-type B (2 strains) consisted of identical clones, the eight strains belonging to the remaining Pulso-types C–J demonstrated<70% similarity and were unlikely to be derived from a single clone of E. coli. It is to be noted that only a few banding patterns were obtained from an E. coli strain (05-M-7-1) due to constant autodigestion of genomic DNA during agrose plug preparation, and thus a cluster formed by this strain is ignored throughout this article.
Discussion
In the present study, the prevalence of PMQR determinants in E. coli isolated from food-producing animals was investigated and the PMQR-positive isolates were characterized using molecular methods. This study provides the first report of plasmid mediated quinolone resistance in E. coli isolates from animals in Korea. Our study yielded a prevalence of 3.8% for PMQR genes among 365 nalidixic resistant and ciprofloxacin susceptible or resistant E. coli isolated from pigs and cattle during the 2003–2011 period. Since only a subset of isolates from the total E. coli isolated during this period was examined, this does not represent the true prevalence of PMQR determinants among the E. coli strains in 2003–2007. Nevertheless, our results are similar to those of previous studies on prevalence of PMQR genes in E. coli isolated from human feces (Jeong et al., 2011) and other clinical specimens (Kim et al., 2009) in Korea. In contrast, a previous study from China reported that 17.2% of the 64 E. coli isolates from food-producing animals contained one or more PMQR determinants (Ma et al., 2009). Similarly, our results demonstrated lower (3.8%, 14/365) prevalence of PMQR genes compared to that of a recent international collaborative study in E. coli isolated from animals, humans, food, and the environment in 13 European countries in which PMQR genes were identified in 15% of the E. coli isolates showing reduced susceptibility to fluoroquinolones and nalidixic acid (Veldman et al., 2011). The lower prevalence of PMQR genes in our study may be related to study design of this work where nalidixic acid susceptible isolates were excluded from PMQR screening.
In Korea, qnr genes were first detected in 0.8% (2/260) of human clinical E. coli isolates collected during 2001–2003 period (Jeong et al., 2005). We identified qnr genes in 2.7% of E. coli isolates and among them qnrS1 was the most predominant qnr subtype which was detected in eight (2.2%) of 365 E. coli isolates investigated. This is in contrast to previous findings from human clinical E. coli isolates (Tamang et al., 2008; Kim et al., 2009) and human fecal E. coli isolates (Jeong et al., 2011) in Korea in which the most predominant qnr subtype was qnrB4. However, like in our study, qnrS1 was the most predominant qnr subtype identified in E. coli isolates in recent reports from China (Huang et al., 2009) and Europe (Veldman et al., 2011). Moreover, qnrS1 has also been reported in E. coli from healthy piglets from Hungary (Szmolka et al., 2011) and in poultry E. coli from Italy (Cerquetti et al., 2009). As shown in other similar studies (Ma et al., 2009; Cerquetti et al., 2009), qnrA was not detected in this study. Furthermore, none of the studied isolates were positive for qnrC or qnrD gene. Thus, our data and those of others support the suggestion that that the most prevalent qnr determinant in isolates of animal origin may be qnrS rather than other PMQR determinant (Strahilevitz et al., 2009; Poirel et al., 2012). Although, the qnrS gene by itself confers low-level resistance to (fluoro)quinolones, it may facilitate the selection of mutants under selective pressure of antimicrobials (Strahilevitz et al., 2009).
In this study, aac(6’)-Ib-cr gene was detected in 1.1% (4/365) of the E. coli isolates examined. In a previous study in Korea, the aac(6’)-Ib-cr gene was detected in 2% of the total clinical Enterobacteriaceae studied, most of which were E. coli (Kim et al., 2009). Recently, we identified this gene in four (0.7%) nalidixic acid resistant non-typhoid Salmonella from livestock in Korea (Tamang et al., 2011), but to our knowledge, it has not been reported in E. coli from animals in Korea. Nevertheless, this gene is geographically widespread and has been reported from several countries of Asia, North America, and Europe with high frequency among both human and veterinary E. coli isolates (Strahilevitz et al., 2009; Poirel et al., 2012). In contrast, in a recent study aac(6’)-Ib-cr was rarely found in Salmonella and E. coli isolates from 13 European countries (Veldman et al., 2011) which might be explained by the inclusion criteria (reduced susceptibility to fluoroquinolones and nalidixic acid) defined for this study. Moreover, there was no isolate that contained multiple PMQR genes which suggests that PMQR determinants in animals in Korea circulate independently. Furthermore, despite previous reports of qepA in human clinical isolates from Korea (Kang et al., 2009; Kim et al., 2009) and Japan (Yamane et al., 2008) or in animal isolates from China (Ma et al., 2009; Huang et al., 2009), none of our studied isolates was qepA positive. The reason for this difference is unclear. Nevertheless, our findings indicate that the prevalence of PMQR determinants in nalidixic acid resistant E. coli from animals from Korea is relatively low and less diverse.
PMQR genes are known to facilitate selection of additional chromosomal resistance mutations in the presence of quinolones (Strahilevitz et al., 2009). We determined mutations in the QRDR of gyrA, gyrB, parC, and parE in all PMQR-positive isolates. Except one wild-type gyrA, all NALR-CIPS isolates harbored single point mutation in the QRDR of gyrA either at codon Ser83 or Asp87, whereas all but one NALR-CIPR isolates contained double point mutations in gyrA and additional single or double mutations in parC in majority of them which support the fact that single mutation in gyrA of E. coli may be sufficient to cause resistance to nalidixic acid but additional mutations in gyrA and/or the topoisomerase IV genes are required for fluoroquinolone resistance. The various mutation types observed in gyrA, gyrB, parC, or parE in this study had been described previously and most of them are associated with increase in MICs of nalidixic acid and/or fluoroquinolones (Hopkins et al., 2005). However, we could not support the role of PMQR determinants in promoting mutations in topoisomerase genes as has been previously described (Kim et al., 2009), since the PMQR-negative isolates were not analyzed in this study.
Phylogenetic analysis showed that a majority of PMQR-positive isolates were of avirulent phylogroups A (n=7) and B1 (n=3) which have been reported as animal and human commensal E. coli strains previously (Jakobsen et al., 2010). The PFGE analysis of the PMQR-positive E. coli revealed great genomic diversity in strains from pigs but the PMQR-positive strains from cattle were relatively homogenous. Among them, three of the eight qnrS1-positive strains or two strains carrying qnrB4 genes isolated from cattle were derived from a particular E. coli clone, respectively, whereas the remaining strains from cattle or pig carrying qnrS1 or aac(6’)-Ib-cr genes were clonally unrelated which indicate that the dissemination of PMQR genes in E. coli from pigs may be attributed to horizontal transmission and in E. coli from cattle was owing to combination of both horizontal and clonal spread.
In this study, a majority of PMQR-positive isolates were recovered during the 2009–2011 period. This can be explained by the fact that more clinical samples were submitted to the diagnostic laboratory of QIA, and also more E. coli isolates were received from the local veterinary service centers for diagnostic investigation during this period. Furthermore, a majority (10/14, 71.4%) of the isolates with PMQR genes were derived from sick animals compared to healthy animals (4/14, 28.6%). These findings suggest that PMQR genes are more frequently present in isolates from sick animals compared to commensal E. coli from the healthy food-producing animals. Nevertheless, this constitutes a potential public health concern since the recovered sick animals or the healthy animals enter the food chain.
There are a few limitations of this study. Since the basis of PMQR gene screening in this study was resistance to nalidixic acid and resistance or susceptibility to ciprofloxacin, this study was limited to investigation of PMQR determinants among the nalidixic acid resistant isolates. In addition, the prevalence of the PMQR determinants among E. coli strains during 2003–2011 reported in this study should be considered a minimum estimation since some of the new qnrB variants would not be detected by the PCR methods used in this study. Furthermore, we neither investigated the transferability of plasmids carrying PMQR genes nor examined the incompatibility group of these plasmids. Since E. coli isolates with PMQR genes demonstrated different pulso-types, further studies are needed to determine transferability and incompatibility of these plasmids because of the possibility of horizontal transfer of these resistance genes to humans via food chain.
In conclusion, this is the first report of occurrence of qnrB, qnrS, and aac(6’)-Ib-cr genes in E. coli isolates from animals in Korea. Isolation of PMQR genes in E. coli from food animals is a matter of concern, since they could be transmitted to humans via food animals. Thus, it is essential to carry out continuous surveillance and monitoring of spread of PMQR genes among bacteria from food animals and food-animal products.
Footnotes
Acknowledgments
We are grateful to Prof. Jungmin Kim of the Department of Microbiology, Kyungpook National University School of Medicine, for kindly providing positive control strains for various PMQR determinants. This work was supported by a grant from the Animal, Plant, and Fisheries Quarantine and Inspection Agency, Ministry of Food, Agriculture, Forestry, and Fisheries, Republic of Korea.
Disclosure Statement
No competing financial interests exist.