
Abstract
There is a growing concern of a public health risk associated with non-O157 Shiga toxin–producing Escherichia coli (STEC) since E. coli serogroups O26, O45, O103, O111, O121, and O145 are frequently implicated in outbreaks of human illness worldwide. Recently, the Food Safety and Inspection Service of the U.S. Department of Agriculture declared these six STEC O groups to be adulterants in beef. We describe here a rapid, sensitive, and highly specific enzyme-linked immunosorbent assay (ELISA) for the detection of these top six non-O157 STEC O groups. The assays were tested against 174 reference E. coli O groups, with 60 clinical isolates belonging to the target O groups and 10 non-E coli strains belonging to the family Enterobacteriaceae. Assays for serogroups O103, O111, and O121 exhibited 100% specificity, while assays for serogroups O26 and O45 had 98.2% specificity, and O145 had 99.1% specificity. ELISA conducted using artificially inoculated ground beef samples displayed 100% accuracy. The sensitivity of the assay was 5×105 colony-forming unit (CFU)/mL, with limits of detection in the range of 1–10 CFU/25 g of ground beef sample following enrichment. The findings of the study suggest that the assay described is simple and rapid, and can be employed to detect target STEC O groups in beef and other food samples. In addition, the assay provides a conceptual framework that can be adapted for the development of similar tests for the rapid detection of other serogroups of E. coli.
Introduction
F
O antigens, a part of the lipopolysaccharide (LPS) complex on the outer membrane of bacteria, are composed of repeat units of oligosaccharides. The sugar residues in the O antigen vary considerably and differ in their arrangement and linkage between and within the oligosaccharide units, making the O-group the most antigenically variable region of the cell. Conventional serotyping for O group identification is based on agglutination reactions and still remains one of the most comprehensive and simple methods for identifying O groups (Orskov et al., 1977; Orskov and Orskov, 1984). However, agglutination reaction cannot be performed for detecting the target O groups in beef samples as mixed cultures cannot be used for serotyping. It is, therefore, well recognized that rapid, sensitive, specific, convenient, and economical diagnostic tests are urgently required for detecting major serogroups in meat and other food samples (Mathusa et al., 2010; NCBA, 2008).
The objective of the present study was to develop a rapid enzyme-linked immunosorbent assay (ELISA) for detecting the six major non-O157 STEC O groups that can be easily adopted by diagnostic laboratories, food industries, regulatory agencies, and researchers. There are currently no simple ELISA-based tests available commercially for the detection of the top six STEC O groups. The concept behind this sandwich ELISA method is easily amenable for further improvement and development of similar detection methods for other bacteria. Therefore, the work presented will be useful for improving the diagnostic capability for foodborne pathogens.
Materials and Methods
Bacteria
All bacterial strains used in this study are from the E. coli Reference Center (ECRC) at the Pennsylvania State University. E. coli strains H311b (O26), K42 (O61), H515b (O103), Stoke W (O111), 39w (O121), E1385 (O145), and a total of 174 other standard reference strains (from the World Health Organization) belonging to serogroups O1 through O181, except O31, O47, O72, O93, O94, and O122, which are not designated (Orskov et al., 1977). Ten clinical isolates that belonged to each of the six non-O157 STEC O groups (n=60 isolates) were also used in the study. Other bacterial species used for determining the specificity of ELISA were Citrobacter freundii, Enterobacter cloacae, Hafnia alvei, Klebsiella pneumoniae, Proteus vulgaris, Salmonella enterica serovars Enteritidis and Typhi, Serratia marcescens, Shigella boydii, and Shigella flexneri.
Antibodies
Purified polyclonal rabbit anti–E. coli antibodies generated against reference strains belonging to serogroups O26, O45, O103, O111, O121, and O145 were obtained from SDIX (Newark, DE). The antibodies were prepared and purified by SDIX using proprietary methods. Purified antibodies against the top six STEC O groups from other sources were also found to be effective for ELISA (data not presented).
ELISA plate preparation
Polystyrene microtiter plates were coated with 100 μL of mouse monoclonal antibody to lipid A (1 μg/mL; Abcam, Cambridge, MA) in 0.05M carbonate buffer (pH 9.0) and incubated overnight at 4°C. The coating solution was removed, and wells were washed twice with 200 μL of phosphate-buffered saline (PBS). They were filled with 200 μL of blocking solution (5% dry milk in 0.01 M PBS with 0.05% Tween 20) and incubated at 4°C overnight. The wells were then washed in distilled water three times and stored at 4°C until use.
Antigen preparation
Reference standard E. coli strains as well as clinical isolates belonging to serogroups O26, O45, O103, O111, O121, and O145 were grown in 5 mL of Veal Infusion Broth (VIB; Difco, Sparks, MD) overnight at 37°C. The cell density of the culture was adjusted to 1 OD600 (optical density of a sample measured at a wavelength of 600 nm) with VIB, and 1 mL of culture was harvested at 6,000 × g for 10 min and resuspended in PBS (1 mL). The cultures were serially diluted in PBS for enumeration. Antigens were derived from bacteria (at 107, 106, 105, and 104 colony-forming unit [CFU]/mL concentrations) by boiling for 1 h at 100°C, and 100 μL of the heat-inactivated antigen was utilized for the ELISA. The same procedure was employed for generating antigens from non–E. coli bacterial species.
ELISA
Antigens (100 μL) of each of the O groups were dispensed in triplicate into microtiter plates coated with anti–lipid A antibodies and placed on a shaker incubator at room temperature (RT) for 1 h. The solution in each well was removed, and the wells were washed four times with 200 μL of distilled water. The washing step was followed after incubation with primary and secondary antibodies as well. Antibodies (1 μg) against each of the O groups were diluted 1000-fold in 5% dry milk in 0.01 M PBS containing 0.05% Tween 20. Respective diluted antibodies (0.1 μg in 100 μL) were added to each well, and plates were incubated for 1 h at RT. Goat anti–rabbit IgG peroxidase conjugate (100 μL at 1:3000 dilution; Sigma-Aldrich, St. Louis, MO) prepared in 5% dry milk in 0.01 M PBS containing 0.05% Tween 20 was added to each well, and the plates were incubated for 1 h at RT. To develop color, 100 μL of 1-Step Ultra TMB (3,3',5,5' tetramethylbenzidine; Fisher Scientific, Pittsburgh, PA) was added to each well, and the plates were incubated for 15 min at room temperature followed by the addition of an equal volume of stopping solution (2 M H2SO4). The OD was examined at 450 nm using a plate reader (Biotek Instruments, Winooski, VT). Wells containing all components of the assay but without antigen served as negative controls for assessing the background noise. The background OD450 for all ELISA was in the range of 0.071–0.113, except for O26 which was in the rage of 0.165–0.201. All background readings were subtracted, and the averages of triplicate readings were resolved. An OD450 of 0.2 above the background was considered positive. The assays were repeated three times. Since binding of antibodies to polystyrene plates may vary, reproducibility of the assays was tested in triplicate using polystyrene plates obtained from different manufacturers.
Detection of O groups in artificially inoculated ground beef samples
Ground beef (25 g) samples purchased from a local store were individually spiked with 1–10 CFU of six strains belonging to each of the six STEC O groups (n=36) and propagated in 250 mL of Tryptic Soy Broth (TSB) medium. All samples were pre-enriched by incubating static for 6 h at 37°C in the presence of vancomycin (16 mg/L). Following pre-enrichment, the cultures were supplemented with bile salts (1.5 g/mL), rifampicin (2 mg/L), and potassium tellurite (1 mg/L), and incubation continued for 18 h at 42°C on a shaker (Valadez et al., 2011; Fratamico et al., 2011; Possé et al., 2008). An un-inoculated ground beef sample that helps to determine the signals from background flora was enriched similarly. Subsequently, the samples were passed through qualitative filter paper (grade 1, 11 μm; Whatman, Maidstone, Kent, UK) to remove debris, the bacteria (1 ml) were harvested by centrifugation at 6000×g for 10 min
Competitive assay to determine antigen binding
Antigens (100 μL) generated from strains belonging to each STEC O group at 106 CFU/mL were mixed individually with 5 μg of anti–lipid A monoclonal antibody and incubated for 1 h at room temperature. The antigen–antibody complex (100 μL) was dispensed in triplicate wells of microtiter plates coated with anti–lipid A antibodies as described. Respective antigens at similar concentration but without incubating with anti–lipid A antibody served as control for each serogroup. ELISA assays were conducted as described. Averages of the absolute value of the three readings are presented.
Sensitivity and specificity of the assays
The sensitivity of the ELISA was determined by detecting signals generated for each reference strain belonging to the target serogroup at different concentrations (107, 106, 105, and 104 CFU/mL). Antigens were prepared from cultures at each dilution, and assays were performed as described. The limit of detection is defined as the minimum CFU required to produce a signal (OD450) of 0.2 above the background. The limit of detection for O groups in ground beef was resolved by artificial inoculation of the target O serogroup at 1–10 CFU/25 g of ground beef samples followed by enrichment and ELISA as described.
These six antibodies were tested for cross reaction against the antigens from standard reference strains belonging to all other serogroups except the target serogroup (n=173). Antibody against each serogroup was tested against 50 clinical isolates (10 isolates for each of the five non-target serogroups) collected from diverse sources and 10 bacterial species belonging to the family Enterobacteriaceae. The specificities (%) of the six antibodies presented are based on the correct activity of the antibody against strains of heterologous O types to the total number of strains tested.
Statistical analysis
Data are presented as means±standard error (SE). Differences between means were tested for statistical significance by using t-test probabilities. Differences were considered significant at p<0.05.
Results and Discussion
A sandwich ELISA was developed that utilizes a capture antibody targeting the highly conserved lipid A moiety of the LPS complex of the Gram-negative bacteria. The heat-stable LPS complex consists of three distinct covalently bound moieties—the innermost and most conserved lipid A; the core oligosaccharide in the middle; and the outermost “O antigen” polysaccharide, which is highly variable and provides the basis for serogroup designation. When the bacterial cells are heat inactivated, conserved lipid A region of the LPS complex is available for capture using anti–lipid A antibody. Further serotype-specific detection is possible using detection antibody that targets the highly variable polysaccharide O-antigenic region. A schematic representation of the ELISA is presented (Fig. 1).
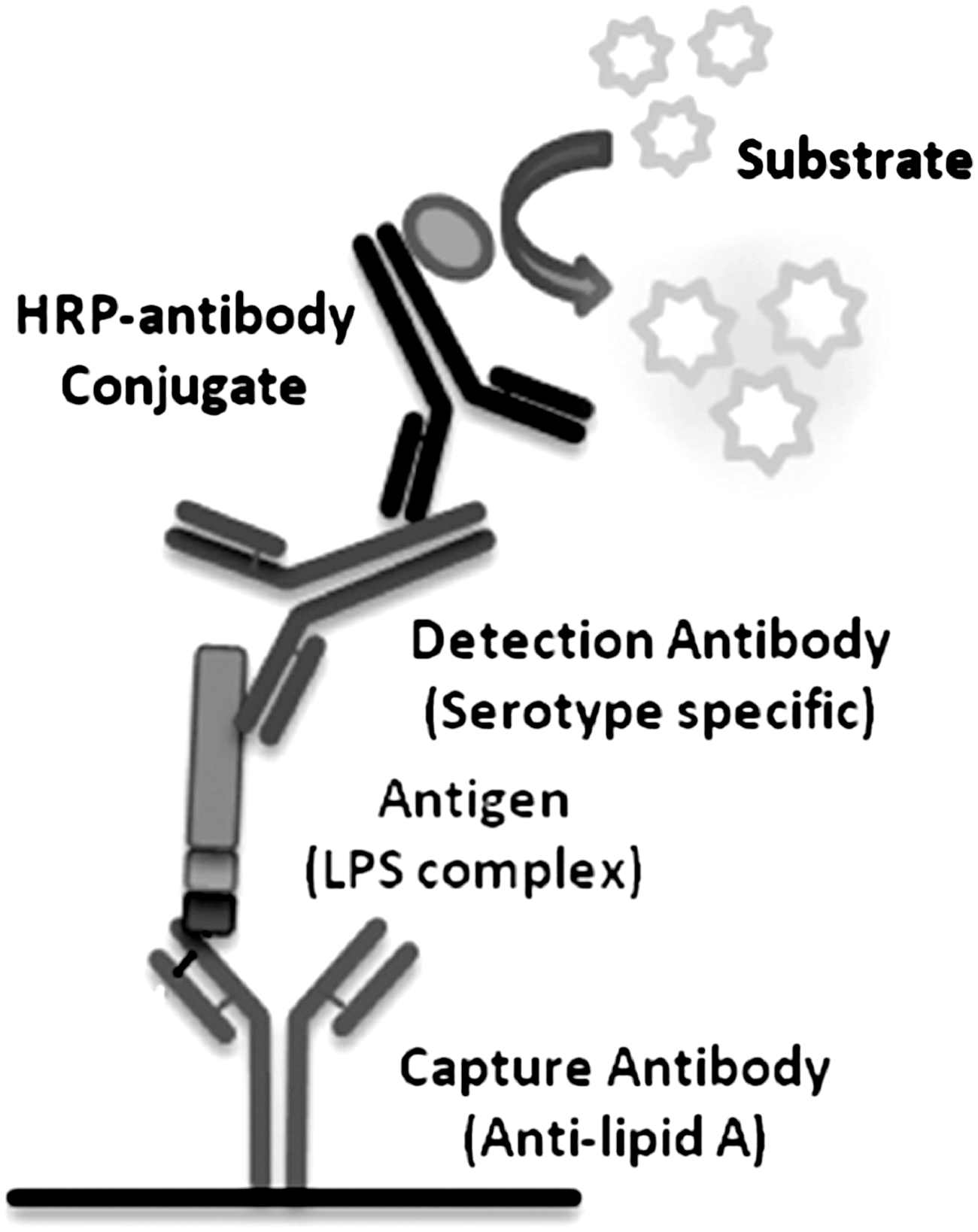
Diagrammatic representation of the enzyme-linked immunosorbent assay (ELISA). Anti–lipid A antibody was used to capture conserved lipid A region of the lipopolysaccharide (LPS) complex that was further detected by antibody targeting the highly variable polysaccharide O-antigenic region.
Sensitivity of the ELISA
The ELISA assays were tested for antigen concentrations derived from 104–107 CFU/mL, and 5×105 CFU/mL concentration was found to be resolving all the O serogroups unequivocally (Fig. 2). The optimum concentration of antibodies, following titration, was found to be 0.1 μg for detecting antigens prepared from 104–107 CFU/mL. Antibodies to serogroup O103 exhibited the highest signal followed by O145, O111, O26, O45, and O121. The differences in the reactivity of the antibodies, as reflected by the signal, could be due to antibody titer, which was found to be greater in O103 than O121. Clinical isolates belonging to each of the top six STEC O serogroups were individually spiked at 1–10 CFU in 25 g of ground beef and enriched for 18 h. ELISA was conducted using antigens derived from 1 mL of the enriched sample to detect the target serogroup (Fig. 3). Assays detected respective serogroups with differences in the signal intensities. It has been reported that novobiocin in the enrichment media can affect the growth of O111 and other non-O157 STEC strains (Fratamico et al. 2010; Vimont et al. 2007). We have recently reported differences in cell counts among the six O groups in artificially inoculated ground beef samples, following 8-h enrichment (Hegde et al., 2012). Therefore, the differences in ELISA signal among the O groups in ground beef samples may be due to differences in cell numbers and/or differential antibody titer.

Detection of top six non-O157 Shiga toxin–producing Escherichia coli (STEC) O groups by enzyme-linked immunosorbent assay (ELISA). The colorimetric signal (OD450 values; y-axis) generated in ELISA for different antigen concentrations prepared from bacteria belonging to the top six non-O157 STEC serogroups (x-axis). The mean values of three independent assays are presented (p<0.001).
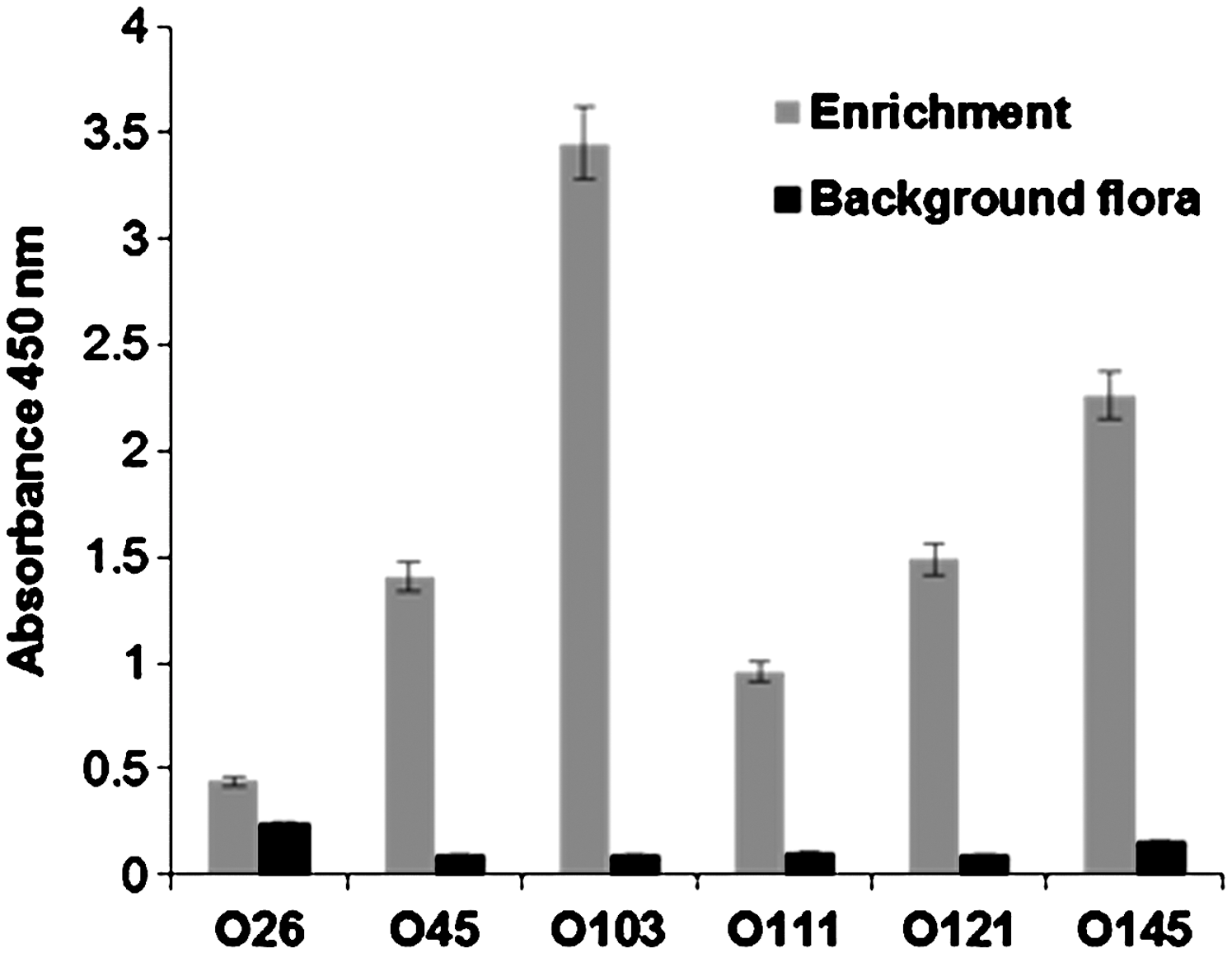
Sensitivity of the enzyme-linked immunosorbent assay (ELISA). Clinical isolates belonging to the top six Shiga toxin–producing Escherichia coli (STEC) serogroup were individually spiked at 1–10 colony-forming unit (CFU) in 25 g of ground beef and enriched for 18 h. Antigens (100 μL) prepared from cells collected from 1 mL of the enriched media were employed in the assay. Gray bars, signal generated for specific serogroups; black bars, control un-inoculated ground beef samples. Absolute mean test values are depicted (p<0.005).
Specificity of the ELISA
Specificities of the six antibodies were tested against strains belonging to all serogroups other than the targeted one, and against 10 clinical isolates belonging to each of the five nontargeted serogroups. As depicted in Table 1, antibodies against O103, O111, and O121 did not cross react with any other E. coli serogroups. However, antibodies against O26 were found to cross react with strains belonging to serogroups O146, O156, O167, and O168; antibodies against O45 were found to cross react with serogroups O146, O153, O156, and O168; and antibodies against O145 were found to cross react with serogroups O150, O151. The six antibodies did not show any cross reaction with 10 of the non–E. coli strains tested belonging to the family Enterobacteriaceae (Table 1). Efforts directed toward generating antibodies highly specific to variable region of the O polysaccharide would help to improve the specificities of this ELISA further.
Cross reactivity of antibodies to top six serogroups tested against reference strains, non–Escherichia coli strains and clinical isolates.
Positive: OD450>0.2 above background; negative: OD450<0.2 above background. The background OD450 for all ELISA was in the range of 0.071–0.113, except for O26 which was in the rage of 0.165–0.201. All assays were conducted using antigens prepared from 106–107 CFU/mL concentration.
STEC, Shiga toxin–producing Escherichia coli (STEC); OD, optical density; ELISA, enzyme-linked immunosorbent assay; CFU, colony-forming unit.
A competitive ELISA was performed to verify whether the lipid A component of LPS complex was truly responsible for binding to the antibodies coated on the plate. To allow blocking of lipid A binding sites, antigens prepared from the six STEC O groups were mixed with an excess amount of anti–lipid A antibodies (50× concentration used for ELISA) and incubated for 1 h before dispensing into the wells coated with anti–lipid A antibodies. Antigen incubated with anti–lipid A antibody resulted in a 75% reduction in signal for all serogroups with the exception of serogroup O121 which exhibited 60% reduction (Fig. 4). This experiment suggested that the lipid A component of the LPS complex was responsible for binding to the anti–lipid A antibody coated on the wells of the microtiter plate. The availability of intact LPS complex for capture of antigens is important for ELISA.
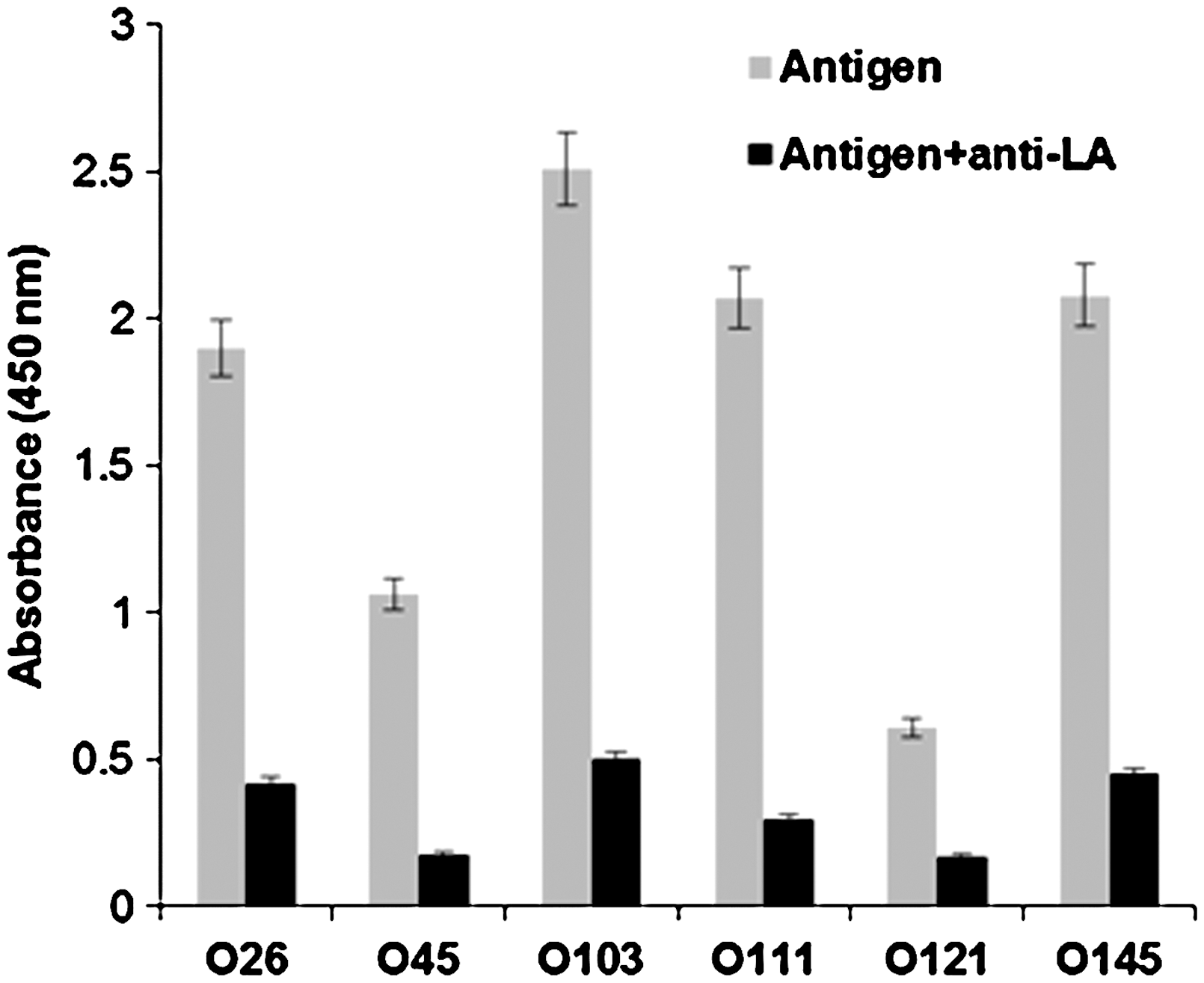
Competitive assay for lipid A binding. The antigens (100 μL) prepared from 106 colony-forming unit (CFU)/mL with and without incubating with anti–lipid A antibodies were tested. Gray bars, absorbance generated with antigens of target bacteria; black bars, reduction in signal when antigens were incubated with anti–lipid A antibody. Antigens incubated with anti–lipid A antibodies showed ∼75% reduction in the signals. Averages of the absorbance from triplicate experiments are presented (p<0.005).
Conclusion
A novel ELISA method is described here for the detection of six non-O157 STEC O groups, which can be used for detecting target O groups in mixed culture in conjunction with PCR assays for detecting Shiga toxin genes (stx-1 and stx-2) and attaching and effacing gene (eae) in beef.
Footnotes
Acknowledgments
We acknowledge the generosity of Herbert Lotman, who has kindly provided funding to Penn State College of Agricultural Sciences for food safety research.
Disclosure Statement
Narasimha V. Hegde, Chitrita DebRoy, and Vivek Kapur have provisional patent for the method described in the manuscript.