
Abstract
Pigs are an important reservoir of hepatitis E virus (HEV) in many countries throughout the world. We evaluated the association between farm size and presence of serum antibodies against HEV, as well as other risk factors for infection in pigs raised in Nan Province, Thailand in a cross-sectional study. The sampling frame was a total-population census of all pig herds, stratified into three classes of the farm size according to criteria developed by the Nan provincial livestock health office. One-eighth of all pigs in each farm were sampled randomly. All pig-farm owners were interviewed to elicit information on general characteristics of their farms, biosecurity and hygienic procedures, and farm management. We obtained sera and fecal samples from 879 pigs to test for antibodies to HEV and HEV RNA. Odds ratios (OR) and 95% confidence intervals (CI) for risk factors for HEV seroprevalence were estimated by multivariate logistic regression. The overall prevalence of anti-HEV immunoglobulin G antibodies was 9.9%. Pigs studied from medium-sized farms had a higher HEV seroprevalence than those from larger farms (adjusted OR 4.95, 95% CI: 1.79, 13.70). Factors associated with HEV seropositivity included feeding pigs with agro-industrial byproducts, having veterinarians on farms, and presence of other pig farms within 100 m. Twenty-five (2.9%) of 875 sampled pig stools were positive for HEV RNA. Phylogenetic analysis revealed that all HEV isolates clustered to HEV genotype 3.
Introduction
H
Thailand agriculture is largely rural village–based, with pigs providing an important source of food and income. There is considerable heterogeneity in pig farm sizes across rural Thailand, ranging from smaller, domestic husbandry for consumption within the home or village, to larger commercial swine herds maintained for export.
We conducted a cross-sectional study in Nan Province, Thailand, to evaluate the role of pigs in HEV transmission to humans. Our objective was to determine seroprevalence of HEV antibodies among domestic pigs in different-sized farms, possible risk factors related to HEV infection in pigs, and to further evaluate the presence of active HEV infection through RT-PCR in stools.
Materials and Methods
Study site and subjects
The study population included 800 pigs from randomly selected herds registered at the Nan provincial livestock health office in 2009. Farms were stratified by size into three classes: small farm <30 pigs, medium farm 30–300 pigs, and large farm >300 pigs. Sampling was carried out across six districts with high density of pig population. Two different comparisons were tested: medium-sized farms versus large-sized farms and small-sized farms versus large-sized farms.
We calculated sample size using Epi Info Version 2.3.1 (Centers for Disease Control, Atlanta, GA). The prevalence of anti-HEV immunoglobulin G (IgG) in large-sized farms was estimated at 9.0%. If prevalence ratio in medium-sized farms and small-sized farms is 1.95 times that of large-sized farms, we needed 269 pigs from each size class to be able to reject the null hypothesis of risk ratio equal to 1 (power 0.8, α=0.05).
We invited owners of randomly selected farms to participate in the study. One-eighth of all pigs in each farm were sampled randomly. As an example, 40 of 320 pigs from a large farm, 4 of 32 pigs from a medium farm, or 1 of 8 pigs from a small farm were sampled. Piglets younger than 2 months, pregnant sows, and nursing sows with litters less than 2 months old were excluded.
Data Collection
Veterinarians used a standard questionnaire to gather information on potential risk factors related to pig HEV infection from farmers. Breed and sex were determined by examination, while age was provided by the farmer. We took 10 mL of blood from the anterior vena cava of each pig. Blood was transported on ice to the Nan Provincial Livestock Health Office and allowed to clot at 4°C, after which serum was separated into 1-mL labeled aliquots and stored at −80°C.
Fecal samples were collected from the rectum via digital manipulation (approximately 3 g of stool material), transferred to a 5-mL tube filled with RNA Later Transport Solution (Ambion, Grand Island, NY), then kept in cold storage for approximately 3 days. Stool samples were homogenized in a 20% stool suspension in RNA Later by vortexing for 1 min. Homogenates were centrifuged and the supernatant was transferred to fresh tubes. All serum and stool specimens were placed on ice packs and shipped to U.S. Army Medical Component of the Armed Forces Research Institute of the Medical Sciences in Bangkok, Thailand, and stored at −80°C until subsequent tests.
Laboratory Analysis
Enzyme-linked immunosorbent assay for HEV antibodies
Detection of HEV IgG in pigs was modified from an enzyme-linked immunosorbent assay procedure described by Innis et al. (Innis et al., 2002) and Wang et al. (Wang et al., 2001). Duplicates were averaged. When duplicates difference fell outside the 20% coefficient of variation, the specimen was retested. The positive cut-off (>0.258) was determined by calculating the mean optical density±3 standard deviations of 30 Thai anti-HEV-negative pig serum.
RT-PCR
HEV RNA was extracted from 140 μL of 20% stool suspension using QIAamp viral RNA mini kit (Qiagen, Germany) according to the manufacturer's instruction. The RNA was tested by nested RT-PCR using primers flanking the junction of open reading frame (ORF) 1, 2, and 3 with external primers F2782 (5′- GGDCTBGTTCATAACCTGAT-3′) and R2783 (5′- GGTTGGTTGGATGAATATAGG-3′) and internal primers F2781 (5′- GTTCATAACCTGATWGGYATGCT-3′) and R2784 (5′- GGATTGCGAAGGGCTGAGAATCA-3′) (Tsarev et al., 1993). Briefly, genomic RNA was converted to cDNA using specific primer R2783 with the AMV-RT (Promega, USA) according to the manufacturer's instruction. The first- and second-round PCR were amplified by using AmpliTaq DNA polymerase (Applied Biosystems, USA) with the external and internal primer sets, respectively. Cycling conditions included 35 amplification cycles of 1 min at 94°C, 1 min at 55°C, and 1 min at 72°C. After electrophoresis in a 1.5% agarose gel stained with ethidium bromide, the expected 310-bp bands of the second-round PCR product were visualized on an ultraviolet transilluminator. The PCR-amplified DNA fragments were excised and purified using QIAquick PCR purification kit and the QIAquick gel extraction kit (Qiagen, Germany) according to the manufacturer's instructions.
Sequencing reactions were performed using the DYEnamic ET dye terminator sequencing kit (GE Healthcare, UK) according to the manufacturer's instruction, with sequencing primers. The sequencing products were cleaned by standard precipitation before sequencing on a MegaBACE 500 automated DNA sequencer (GE Healthcare, UK). Overlapping nucleic acid sequences were combined for analysis and edited with the aid of Sequencher software (Gene Code Corp., USA). The genome sequences of 20 Thai isolates were compared with 20 known HEV swine and human sequences obtained from GenBank.
Sequences were aligned using molecular evolutionary genetics analysis (MEGA) software version 4.0 (MEGA4) (Tamura et al., 2007), and a phylogenetic tree was constructed using PhyML v3.0 (Guindon and Gascuel, 2003) under the model GTR+G+I determined using best-fit substitution model (maximum likelihood).
Statistical methods
Statistical analyses were performed using STATA SE10 (Stata Corporation, College Station, TX). We used multiple logistic regressions to investigate associations between risk factors and HEV seropositivity. Variables were included based on p<0.05 significance in univariate analysis. The model was reduced in a backward-elimination procedure. A 10% change in the coefficient was considered evidence of possible confounding. Adjusted OR and 95% CI were calculated for main effects.
Results
Overall prevalence of IgG anti-HEV among pigs was 9.9% (87/879). Twenty-five (7.9%) of 315 pigs from the large farms, 40 (14.8%) of 271 pigs from medium farms, and 22 (7.5%) of 293 pigs from small farms were tested positive for IgG anti-HEV (Table 1).
Of the 94 herds, 29 were positive for HEV antibodies (30.9% herd prevalence). Anti-HEV IgG seroprevalence among farms ranged from 0 to 66.7%. Large farms and small farms had similar prevalence of HEV antibodies, but differed from that observed in medium farms.
Eight hundred fourteen of all pigs were Hampshire crossbreeds (92.6%). General characteristics of farms by size class are shown in Table 2. None of the medium farms treated drinking water before it was delivered to the farms. Few medium farms and some small farms reported never cleaning floors with disinfectant. Most of the small farms discarded manure nearby the premises without utilizing hygienic practices to disinfect the manure or contain it in lagoons. Access to veterinary services was more frequent in the large farms than in the medium and small farms.
CI, confidence interval.
In a multivariate logistic regression analysis comparing the medium and large farms, the adjusted OR was 4.95 (95% CI: 1.79–13.70) for a higher HEV seroprevalence among pigs from the medium-sized farms (Table 3). No difference was found for anti-HEV prevalence between the small versus large and small versus medium farm size. The adjusted OR was 0.91 (95% CI: 0.29–2.87) (Table 4) and 0.53 (95% CI: 0.24–1.15) (Table 5), respectively.
CI, confidence interval.
CI, confidence interval.
In a model comparing seroprevalence among pigs from medium and large-sized farms, the following factors were significantly associated with anti-HEV: feeding pigs with agro-industrial byproducts, having veterinarians on the farm, and presence of other pig farms within 100 m (Table 3). In a model of both small versus large and small versus medium farm size, after adjustment using multivariate logistic regression analysis, having veterinarians on the farm was protective for anti-HEV seroprevalence (Tables 4 and 5). In addition, feeding pigs with agro-industrial byproducts and the presence of other pig farms within 100 m were protective, whereas presence of birds' feces inside the pig house was a risk factor for anti-HEV seroprevalence (OR=2.87, 95% CI: 1.07–7.71) for small versus medium-sized farm (Table 5).
Twenty-five (2.9%) of 875 stool samples were positive for HEV RNA by RT-PCR using primers that span the ORFs 1, 3, and 2 of the viral genome. Eleven (20.0%) of 55 stool samples from 2-month-old pigs, seven (5.1%) from 137 stool samples of 3-month-old pigs, and six (3.3%) from 184 stool samples of 4-month-old pigs were positive for HEV RNA. All pigs 5 months or older were negative for HEV RNA. Female pigs were positive for HEV RNA from 11 (3.2%) of 339 stool samples and male pigs had 13 (2.4%) of 535 HEV RNA positive rectal samples.
The 20 Thai isolates were compared with 20 known HEV swine and human sequences obtained from GenBank as shown in Figure 1. The accession numbers of 20 HEV sequences at the junction between ORF 1, 2, and 3 are JQ359025–JQ359044. Sequence analysis of the 300-bp fragment indicated that all 20 Thai isolates belonged to genotype 3 and were 88.3%–100% identical to each other. When sequences were compared to known human and pig HEV isolates obtained from GenBank, they had 89.7%–95% sequence homology with genotype 3 strains, 84.7%–85.7% homology with Japan genotype 4 HEV (AB481227), 83.0%–84.7% homology with Nepal genotype 1 HEV (AF051830), and 85.3%–87.0% homology with Mexican genotype 2 HEV (M74506). Genotype 3 sequences from both human and pig HEV isolates appeared to cluster indiscriminately within the phylogenetic tree.
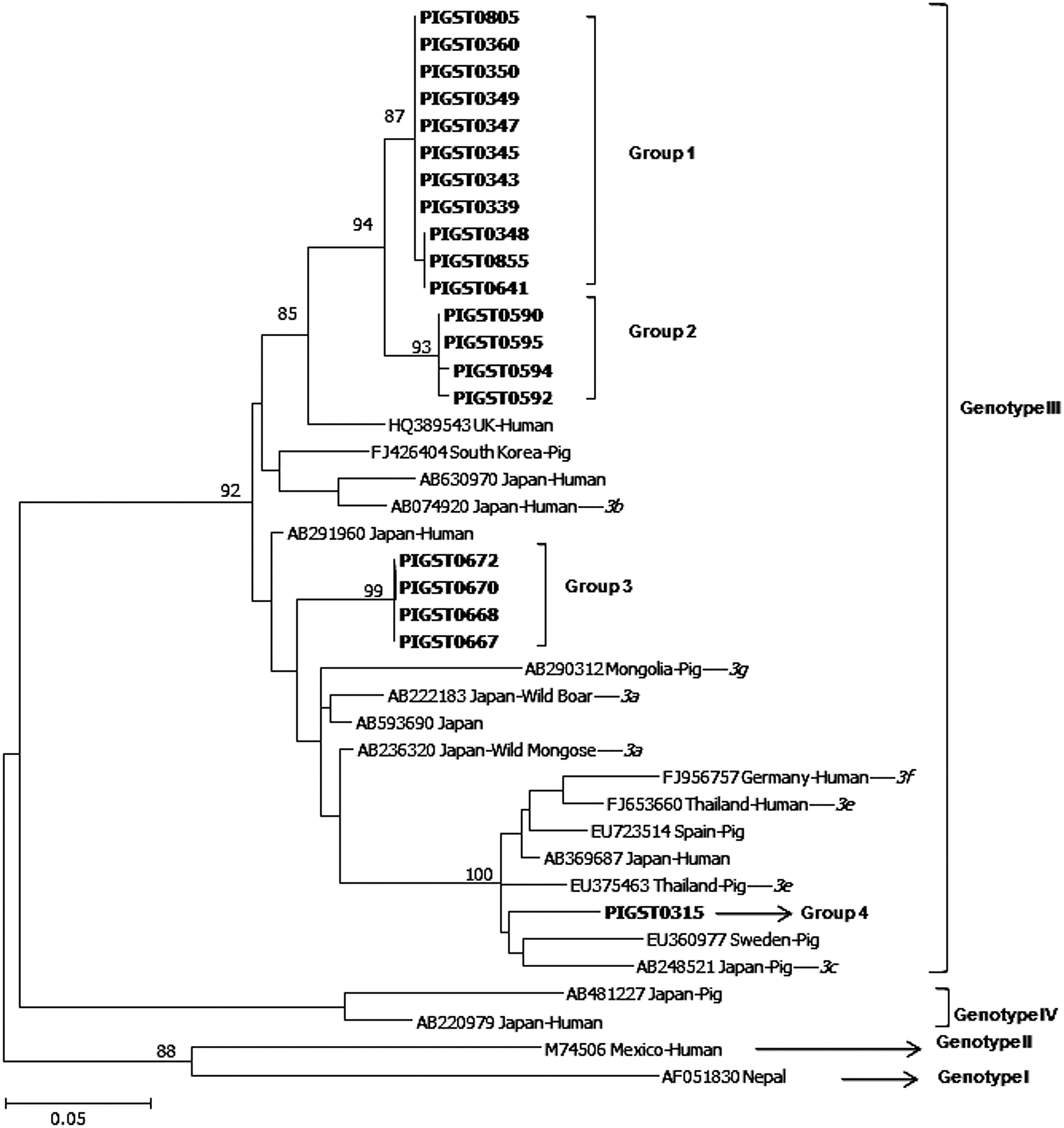
The phylogenetic tree constructed by the maximum-likelihood method (GTR+G+1, model) based on 300 nucleotides and 40 sequences in junction with open reading frame (ORF) 1, 2, and 3 of the hepatitis E virus (HEV). Twenty HEV isolates from Thailand pigs are listed in bold type, compared to human and swine sequences obtained from GenBank. Genotypes and subtypes (3a–g, italics), if known, are denoted on the tree, and genetically distinct Thailand groups are enclosed in brackets.
Genetic variability was observed within the 20 Thai HEV isolates, and we tentatively assigned them into four apparent subgroups. Group 1 could not be classified to the subtype level as there was 94.7%–95.0% sequence homology with HEV isolate from United Kingdom (GenBank accession number: HQ389543) and 93.0%–93.3% sequence homology with subtype 3b human HEV isolate in Sapporo prefecture, Japan (AB074920). Group 2 had 93.0%–93.3% sequence homology with a subtype 3b human HEV isolate from Sapporo prefecture, Japan (AB074920). Group 3 could not be classified to the subtype level as there was 95.7% sequence homology with subtype 3a (AB236320), 93.3% sequence homology with subtype 3b (AB074920), 90.3% sequence homology with subtype 3c (AB248521), 90.7% sequence homology with subtype 3e (EU375463), and 89.3% sequence homology with subtype 3f. Group 4 had 95.0% sequence homology with HEV isolate from Thailand pig subtype 3e (EU375463). Group 1 came from the four farms in four districts. Group 2, group 3, and group 4 came from the different farms and different districts. We also compared our isolates to representative isolates from other genotypes. Our isolates share 83.0%–84.7% nucleotide sequence homology with an HEV isolated from Nepal (genotype 1, accession number AFO51830), 85.3%–87.0% sequence homology to an HEV isolate from Mexico (genotype 2, M74504), and 84.7%–85.7% to an isolate from Japan (genotype 4, AB481227).
Discussion
Studies in several countries have documented pigs as the most important zoonotic reservoir of genotype 3 and 4 hepatitis E virus infections (Clayson et al., 1995; Hsieh et al., 1999; Meng et al., 1999; Labrique et al., 2010). Our study of pigs from a rural province in Northern Thailand found a lower prevalence of anti-HEV IgG and HEV RNA compared to two other studies in pig populations from other areas of Thailand (Meng et al., 1999; Siripanyaphinyo et al., 2009). The pig population from Nan is isolated and infrequently exposed to infected pigs from other provinces.
Our investigation of 94 pig farms revealed that the prevalence of HEV infections varied from 0 to 66.7% within different farms. Pigs from large farms and small farms had similar HEV seroprevalence, but pigs from medium-sized farms had higher seroprevalence. Large pig farms had a higher biosafety and hygienic standard than medium-sized farms. In Nan, there are less than 15 large commercial pig farms. Medium-sized farms and family-scale farming are increasing in frequency and play an important role in the total pork production.
Surprisingly, management, sanitation, and infrastructure in large farms and small farms were generally better than in medium-sized farms. One explanation may be that lower pen density (fewer pigs per pen) on the small farms results in better sanitation than the medium-sized farms with higher densities. Although pigs were often crowded in large farms, owners of large farms were better able to invest in sanitation improvements. This study showed significantly higher anti-HEV prevalence among pigs in medium farms than in large-scale farms. Poor sanitation and housing management of small-scale farms is associated with higher prevalence of hepatitis E (Emerson and Purcell, 2003; Li et al., 2009; Meng, 2010).
Overall, HEV pig seroprevalence in Nan province was lower than other areas in Thailand (Blacksell et al., 2007). The relatively isolated location of Nan province and less frequent contact between pigs on small farms may have reduced transmission between pigs on small farms. However, in medium-sized farms, substandard hygiene may have resulted in more cross-transmission of HEV between pigs. In large farms, improved hygiene may have reduced risk of HEV transmission.
We identified several factors associated with HEV infection. Feeding pigs with industrial and agricultural byproducts, having a veterinarian available, and presence of other pig farms nearby were protective against HEV infection when adjusted for other risk factors. Currently, there is limited understanding of the role of diet in HEV infection. Evidence suggests that some enteric infections can be influenced by dietary factors. Agro-industrial byproducts such as palm kernel meal, brewers dried grain, wheat, or other wastes contain nonstarch polysaccharides, which may protect against transmission of environmental bioagents (Iyayi, 2004; Panda and Kumar, 2006). Adequate feeding of pigs with starches may decrease the frequency of habitual coprophagia by pigs, thereby reducing the rate of fecal–oral HEV transmission.
Our study suggests that availability of veterinarians to pig farms was protective against HEV infection, likely because veterinarians are a key source of education regarding farm management and sanitation. The presence of other pig farms nearby was also protective against HEV infection in pigs. Clustering of pig farms potentially increases access to veterinary services, consequently improving hygienic conditions. This is contrary to recommendations that piggeries should be separated from each other to reduce risk of disease transmission. In rural areas of Thailand such as Nan province, there are a few veterinarians. Therefore, a small pig farm is more likely to receive advice and professional veterinary services when it is located near another pig farm.
Pigs 2–3 months of age had higher shedding peaks, suggesting that these pigs posed the greatest risk of spreading HEV through stools. This is consistent with previous studies in Japan, Korea, and Taiwan (Wu et al., 2002; Choi et al., 2003; Takahashi et al., 2003; Nakai et al., 2006; Lee et al., 2009) that found viremia occurred in pigs approximately 2–4 months of age. In contrast, this study showed that prevalence of anti- HEV increased in pigs older 6 months of age, but the differences were not significant. Previous studies (Clayson et al., 1995; Wu et al., 2000; Wu et al., 2002; Guimaraes et al., 2005; Nakai et al., 2006; Seminati et al., 2008; Lee et al., 2009) have shown that the highest proportion of IgG-positive pigs was among the adult sows or all adult pigs. This phenomenon may be due to most sows having already been infected prior to pregnancy; then passively transmitted maternal antibodies to HEV may provide resistance to their piglets against HEV during the first 1–3 months of life; however, maternal antibodies are believed to wane in the first 1–3 months in young pigs (Clayson et al., 1995). Therefore, if 1–3-month-old pigs were infected with HEV, these young pigs may be the group at greatest risk for HEV shedding, and those of older ages might have already been infected and cleared the virus.
In this cross-sectional study, we could not be certain of the temporal relationship between risk factors and HEV infection. Risk factors from the more remote past might have been missed. It is also important to note limitations of interpreting sequence data from a 300-bp fragment; nevertheless, genetic variability was evident among the Thai HEV isolates. Interestingly, four genetically distinct isolates were detected in Nan province, which could reflect movement of people and animals in the region. These subtype differences may also be associated with viral characteristics that affect human and porcine epidemiology of HEV.
In conclusion, proper management of farm hygiene may effectively control HEV infection among pigs raised in farms. Pigs shed HEV in their stools and can spread infection to other pigs through drinking water or by coprophagia. Although the prevalence of HEV infection was lower among pigs in the farms included in our study in Nan Province, there may be a greater risk of exposure to HEV infection among persons having a higher risk of severe infection, such as pregnant women and immunocompromised persons, in these small farms than in large industrialized farms. Improved awareness about the risk of HEV from contact with pigs is needed for persons living on small rural farms.
Footnotes
Acknowledgments
The authors acknowledge the veterinarians and technicians from Department of Livestock Development, the pig production companies, the farmers for their cooperation, and Dr. Alden Henderson from Bureau of Epidemiology, Ministry of Public Health for his help. The support of the World Health Organization's South-East Asia Regional Office is appreciated. We also thank Punnarai Smithsuwan, Tanawat Likitkererat, Dr. Pravit Choomkasien, and Dr. Sopon Iamsirithaworn for advice during the data entry and data analysis. Finally, we thank the staff from the USAMC-AFRIMS in Bangkok for help in the diagnostic testing.
Disclosure Statement
No competing financial interests exist.