
Abstract
Staphylococcus aureus is a pathogenic bacterium capable of developing biofilms, leading to nosocomial infection and cross-contamination of foods. The current study was focused on the detection of adhesin genes, staphylococcal nuclease and hemolytic activities, and biofilm formation among the isolates of S. aureus from different sources. Fifteen adhesin genes (bap, bbp, clfA, clfB, cna, ebpS, fib, fnbA, fnbB, eno, icaAD, icaBC, sasG, sasC, pls) involved in S. aureus cell aggregation and biofilm accumulation were detected by polymerase chain reaction using specific primer. The activities of staphylococcal nuclease and hemolysis were analyzed by using toluidine blue–DNA agar and sheep blood agar for each strain. The ability of biofilm formation among different S. aureus strains was tested by using the glass tube method and microtiter-plate method. Our results showed the diversity of biofilm formation from different sources. Some isolates were strong biofilm producers; some were weak biofilm producers; and some were nonbiofilm producers. Staphylococcal nuclease and hemolysis seem to play a certain inhibitory role in biofilm formation. The adhesin genes varied among different S. aureus strains. The bap gene was not present in any strains. The bbp gene was only detected in one strain. The detection rates of other adhesin genes were as follows: clfB and sasG (100%); cna, eno, fib, and ebpS (93.75%); fnbA, icaAD, and icaBC (87.50%); fnbB (68.75%); sasC (31.25%); clfA (25%); and pls (12.50%), respectively. The variation between phenotypic and genotypic characterization may be due to the heterogeneity in the genetic origins. There was no direct correlation in distribution of adhesin genes and biofilm formation, which indicates that a single gene or subset of genes cannot be utilized as a biofilm indicator for morphology. Our results also indicated that biofilm formation might be affected by many factors, which brings new challenges to the prevention of this serious pathogen due to biofilm-related infection and contamination.
Introduction
Molecular studies have shown that during late phases of adherence, the organisms first adhere to each other and then elaborate a biofilm. This ability depends on the production of the extracellular matrix of S. aureus, mainly composed of poly-b(1,6)-N-acetyl-
Some S. aureus strains have the ability to produce nucleases to break down extracellular DNA, hence inhibiting biofilm formation, which has been verified in some specific laboratory strains (Mann et al., 2009; Tang et al., 2011c).
With regard to the complexity and variability of the biofilm phenotype, genetic studies of biofilm formation by S. aureus have involved the search for biofilm-defective mutants. Such an approach has identified a number of genetic loci required for biofilm formation, which include many adhesin genes (Shanks et al., 2008). However, these data still call into question the importance of these genes. It is unclear whether all the adhesin genes play important role in biofilm formation of clinical S. aureus isolates. The present study aimed to investigate biofilm-forming properties, the activities of staphylococcal nuclease and hemolysis, and the different adhesin genes in S. aureus isolates from different sources.
Materials and Methods
Bacterial strains and growth conditions
Fifty-seven S. aureus isolates were investigated. Among them were 30 strains (S-C1, S-C2, to S-C30) isolated from chicken (Tang et al., 2011a); eight strains (S-F1, S-F2, to S-F8) isolated from samples associated with food poisoning outbreaks (Tang et al., 2011b); and 19 strains (S-G1, S-G2, to S-G19) isolated from different goats slaughtered to send to food markets in four slaughterhouses (Tang et al., 2012). All isolates were analyzed for the presence of staphylococci on Baird-Parker agar and also identified as S. aureus by specific genetic tests (16s rDNA). Stock cultures were maintained in 15% glycerol at −80°C. All strains were thawed and subcultured in tryptic soy broth (TSB) for 18 h at 37°C, 180 rpm prior to use.
Glass tube biofilm assay
Glass tube biofilm-formation ability was tested using a method previously described by Tang et al (2011c).
Quantitative biofilm formation on polystyrene
Biofilm-forming ability was measured by determining the adhesion to polystyrene microtiter plates (Tang et al., 2011c).
Staphylococcal nuclease assay
To assay staphylococcal nuclease activity, bacteria were grown in TSB at 37°C for 18 h and then heated in boiling water for 15 min. Aliquots (10 μL) of each boiled culture supernatant were tested for staphylococcal nuclease activity using toluidine blue–DNA agar according to the method described by Lachica et al. (1971).
Hemolytic activity observation
Hemolytic activity was studied on agar containing 5% sheep blood. An overnight subculture of each isolate in TSB was streaked and inoculated on sheep blood agar media, which were subsequently incubated at 37°C for 48 h. The sheep blood agars were then incubated at 4°C for a further 4 h (because the β-hemolysin is “hot–cold” hemolysis). Then the hemolytic activities were investigated with the presence of clear zones around the colonies due to the erythrocyte lysing (Wang et al., 2011).
Extraction of genomic DNA
Genomic DNA used for polymerase chain reaction (PCR) analysis was isolated from the isolates by using the extraction method described by Tang et al. (2006).
Specific primers
The adhesin gene primer sets were designed according to the S. aureus icaAD, icaBC, clfB, clumping factor A, fnbA, fnbB, ebpS, eno, fib, sasG, sasC, bbp, bap, cna, and pls gene sequence by using the Primer Premier 5.0 software (PREMIER Biosoft, Palo Alto, CA). All primer pairs were also aligned by using the Basic Local Alignment Search Tool (
Detection of adhesin genes
Amplifications of 15 adhesion genes (icaAD, icaBC, clfB, clfA, fnbA, fnbB, ebpS, eno, fib, sasG, sasC, bbp, bap, cna and pls) were performed by PCR. Reactions were carried out in a volume of 20 μL with a 10×PCR buffer; 2 μL of 2.0 mM MgCl2; 200 mM of each of the following nucleotides: dATP, dTTP, dGTP, and dCTP; 1 U of Taq polymerase (from TaKaRa Biotechnology Co., Ltd., Dalian, China); 1 μL of purified sample DNA; and 0.5 mM of each primer (using primers for each gene in separate reaction mixes). The mixtures were submitted to a program performed on a thermocycler (Mycycler, Bio-Rad, Hercules, CA) with an initial denaturation step at 95°C for 5 min, 35 amplification cycles each with 40 s at 95°C; 50 s at different temperatures for different genes (Table 1); and 50 s at 72°C, followed by an additional extension step of 10 min at 72°C.
Statistical analysis
Data for optical density (OD) values were collected from three independent experiments, and each was performed in triplicate. Statistical analyses were performed by using SPSS version 17.00.
Results
Biofilm formation analysis
The biofilm formation of all 57 strains on glass tubes exhibited considerable variations. Then, we selected 16 strains to further analyze the ability of strains to form biofilm both on glass tubes and on polystyrene (Fig. 1A and B). According to the previous research, the cut-off value (ODc) for determining a biofilm producer was defined as twice the negative control value (Tang et al., 2011c). Based on the OD values of each strain, S. aureus isolates S-C4, S-C6, and S-C8 were defined as strong biofilm producers (2×ODc≤OD); isolates S-C3, S-C9, S-F2, S-F8, S-G1, S-G4, and S-G8 were defined as weak biofilm producers (ODc≤OD≤2×ODc); and isolates S-C10, S-F4, S-F5, S-F6, S-G10, and S-G15 were defined as nonbiofilm producers (OD≤ODc) (Fig. 1B). However, the ability of strains to form biofilm on polystyrene was observed to be inconsistent with that on glass tubes (i.e., isolates S-C4, S-C9, S-C10, S-G4, S-G15, S-F4, and S-F8) (Fig. 1A).
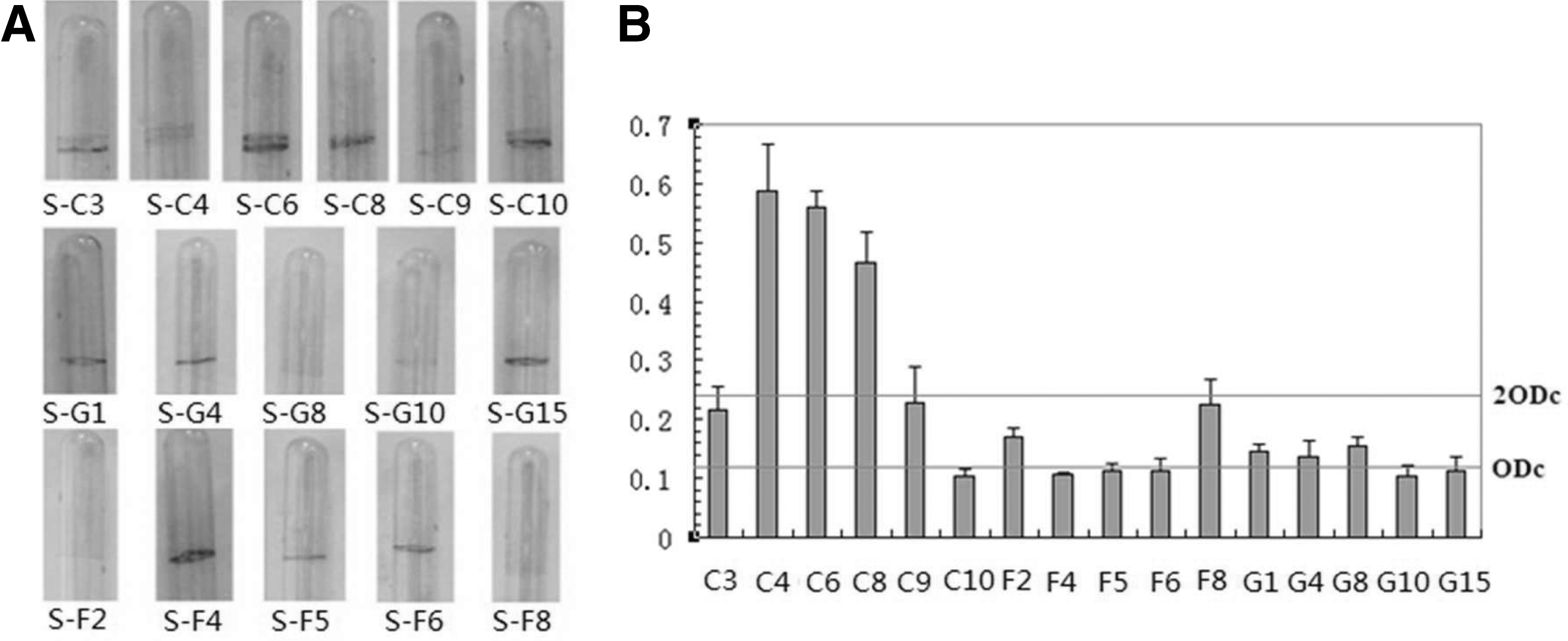
Analysis of biofilm formation.
Staphylococcal nuclease and hemolytic activities observation
The thermostable nuclease activities of 16 isolates are shown in Figure 2A. For S. aureus isolates S-C3, S-C4, S-C6, S-C8 and S-C10, hardly any thermostable nuclease activities were present; S-G1, S-G10, S-F4, S-F5 and S-C9 could produce very weak nuclease activities; while S-G4, S-G8, S-G15, S-F2, S-F6, and S-F8 produced comparatively stronger nuclease activities than other isolates due to the diameter sizes and color depth of different pink halo on toluidine blue–DNA agar. For the hemolytic activity observation, S-G1, S-G4, S-G8, S-G10, S-G15, and S-F5 exhibited strong hemolytic activities because of a large zonal radius of complete hemolysis on sheep blood agar; S-F8 isolate represented a weak hemolytic activity compared to isolates S-F2, S-F4, S-F6, S-C3, S-C4, S-C6, S-C8, S-C9, and S-C10, which had no changes on sheep blood agar (Fig. 2B). These isolates exhibited different characters due to their sources. All the tested isolates from goats had strong hemolytic activities, while all the tested isolates from chicken did not present any hemolytic activities. Among the isolates from food-poisoning samples, only S-F5 had a strong hemolytic activity and S-F8 had a weak hemolytic activity.
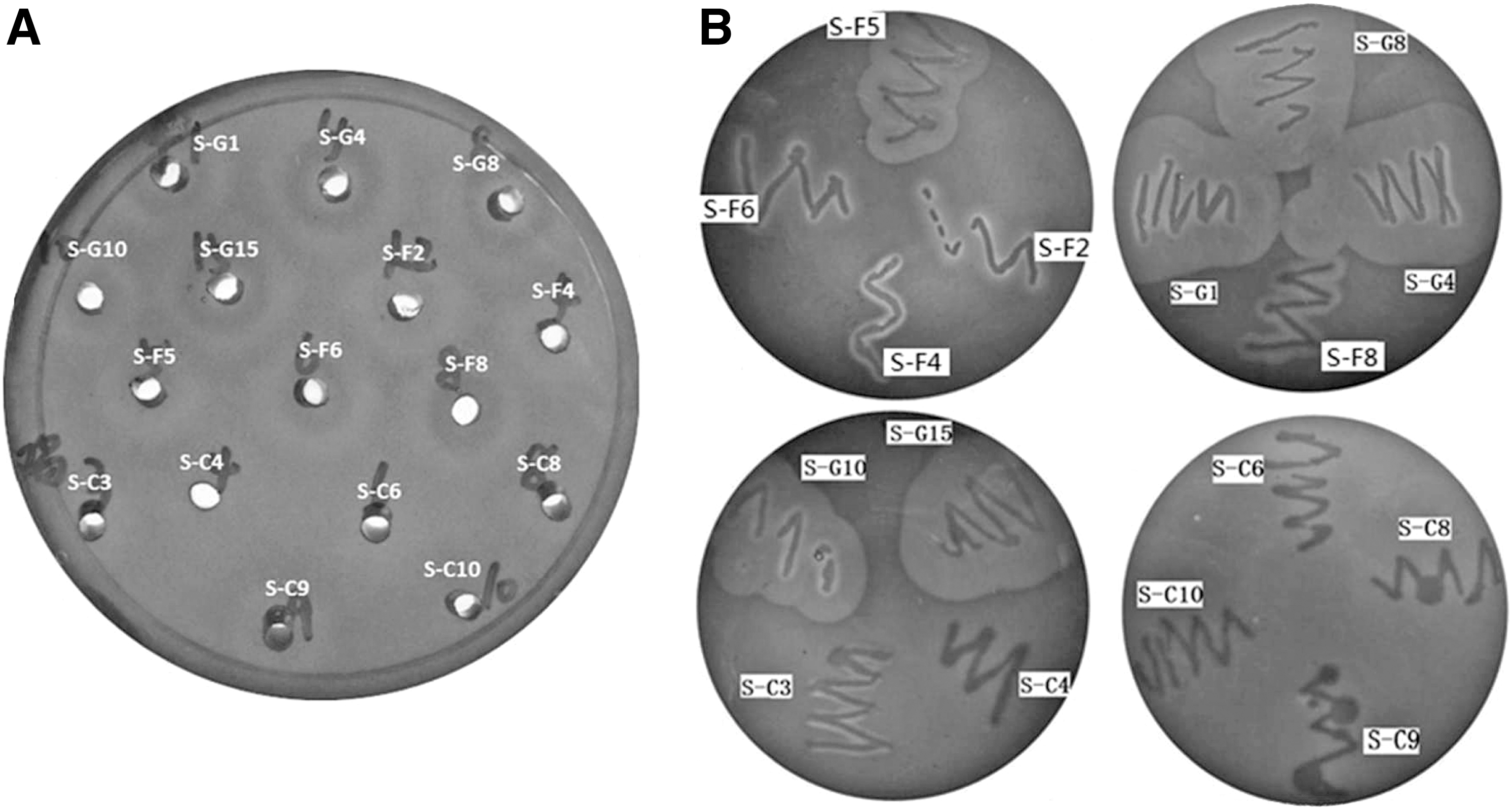
Staphylococcal nuclease and hemolytic activities observation.
Diversity of adhesin genes
We detected 15 adhesin genes (bap, bbp, clfA, clfB, cna, ebpS, fib, fnbA, fnbB, eno, icaAD, icaBC, sasG, sasC, pls) involved in S. aureus cell aggregation and biofilm accumulation. The results showed that the adhesin genes varied among different S. aureus strains. In Table 2, it can be seen that the bap gene did not appear in any detected strains and bbp gene was only detected in one strain. The detection rates of clfB and sasG genes were 100% (16/16); can, ebpS, eno, fnbA and fib genes were 93.75% (15/16); icaAD and icaBC genes were 87.50% (14/16); fnbB was 68.75% (11/16); sasC was 31.25% (5/16); clfA was 25% (4/16); and pls was 12.50% (2/16), respectively. It seemed that C10 and G10 were different from the other strains in genetic pattern, in which the icaAD and icaBC genes were not detected.
Discussion
Previous research showed that biofilm formation depends on the characteristics of the surface, the bacterial cell, the growth medium and other environmental conditions (Donlan and Costerton, 2002). In this study, the biofilm formation observed on polystyrene and on glass tubes was not completely consistent. This kind of phenomenon was always observed in our lab. Similar observations were also reported by other authors (Vázquez-Sánchez et al., 2013). The possible reason could be the different hydrophobic and hydrophilic characteristic between polystyrene and glass surfaces. Vázquez-Sánchez et al. (2013) showed that S. aureus strains expressed moderate hydrophobicity, indicating a lower initial adhesion to hydrophobic polystyrene compared to hydrophilic surfaces such as glass. Mafu et al. (2011) reported a moderate hydrophobicity and a low tendency to attach to polystyrene in S. aureus, but a single strain was used. Our study tested 16 different isolates, which indeed showed a certain difference in biofilm formation on these two different materials. These indicated that different materials had a big impact on biofilm formation and adherence.
Furthermore, a number of soluble extracellular proteins also can affect biofilm formation. The relationship between staphylococcal nuclease activities and biofilm formation was reported by several articles. Mann et al. (2009) explored the possibility that staphylococcal nuclease might play a role in biofilm development. In their study, biofilms produced by the nuc mutant (UAMS-1471) were thicker compared to the parental and complemented strains. Our previous study (Tang et al., 2011c) also demonstrated the role of staphylococcal nuclease on preventing biofilm formation in S. aureus RN4220 strain. Kiedrowski et al. (2011) further examined the regulation and role of staphylococcal nuclease in biofilm maturation. All these findings demonstrated that staphylococcal nuclease had an inhibitory impact on biofilm formation in S. aureus. However, these studies focused on each specific laboratory strain. In this study, we focused on investigating different clinical isolates. The biofilm formation abilities of S-C3, S-C4, S-C6, S-C8, and S-C10 strains isolated from chicken, which did not exhibit any staphylococcal nuclease activity phenotype, were indeed stronger than others. These findings were basically consistent with the above description, which further demonstrated that non-nuclease-producing S. aureus strains could form comparatively stronger biofilm. However, the strong nuclease-producing isolates (S-F2, S-F6, S-F8, S-G4, S-G8, and S-G15) and the weak nuclease-producing isolates (S-G1, S-G10, S-F4, S-F5, and S-C9) in biofilm formation had no obvious distinction. We presumed that there could be other factors that influence the biofilm phenotype in these isolates. Considering that S. aureus can produce a lot of extracellular proteins, we suggested further analyzing the relationship between the hemolytic activity and biofilm formation. A researcher reported that inactivation of the hla gene encoding α-hemolysin resulted in a strain capable of initial attachment but incapable of the cell-to-cell interactions required for biofilm accumulation (Cue et al., 2012). In our study, the isolates S-C3, S-C4, S-C6, S-C8, and S-C9, which could form comparably stronger biofilm than other strains, did not exhibit any hemolytic activity on sheep blood agars. It seemed that the hemolysin activity did not have a commensurate increase in biofilm formation. Actually, isolates S-G1, S-G4, S-G8, S-G10, S-G15 and S-F5, which exhibited strong hemolytic activities, had no significant difference with the isolates S-F2, S-F4, S-F6, S-F8, and S-C3, which presented very little or no hemolytic activities. The abovementioned conclusion did not agree with our results. Linked with our previous research, isolates from chicken were less virulent than the other isolates from goats and from humans (Tang et al., 2011a; 2011b; 2012). We speculated that the more virulent the strain is, the less the biofilm forms. That is why so many S. epidermidis strains could cause biofilm-associated infections (Otto, 2012). S. epidermidis is much less virulent than S. aureus and the capacity to form biofilms is considered the most important trait of S. epidermidis. It is reported that the staphylococci are the leading isolates of infections related to intravenous catheter use, and S. epidermidis is more commonly isolated, while S. aureus is more associated with morbidity and mortality (Cue et al., 2012). Another reason we presumed might be a certain interaction between staphylococcal nuclease and hemolysin, which could co-influence the biofilm formation phenotype. Further investigations should be done in the future to deepen this study.
Furthermore, the involvement of biofilms in clinical infections has received increasing interest due to characterization of genes involved in biofilm formation (Atshan et al., 2012). Different strains isolated from different clinical sources can make a difference in the adhesion and/or ability to form biofilm. In this study, the different isolates from different sources exhibit diversity. The variation between phenotypic and genotypic characterization may be due to the heterogeneity in the genetic origins, and not because of the presence or absence of genes required for the biofilm. Therefore, there was no statistical difference in the distribution of genes in S. aureus strains between high and low biofilm formation.
Apart from the abovementioned features, we also observed that isolates S-C10 and S-G10 were different from the other strains in terms of genetic pattern. The icaAD and icaBC genes were not detected in isolates S-C10 and S-G10. Heilmann (2011) reviewed that S. aureus harbors a variety of proteinaceous and nonproteinaceous adhesions that mediate attachment to a multitude of host factors and is essential for biofilm accumulation. The MSCRAMM family belongs to proteinaceous adhesins and the PNAG/PIA belongs to nonproteinaceous adhesins. According to this, the biofilm formation of S-C10 and S-G10 might belong to the proteinaceous adhesins pattern, in which secreted proteins and extracellular DNA appear to substitute for PIA/PNAG.
Moreover, FnbA and FnbB are large adhesive proteins that are important for virulence (Shanks et al., 2008). Most S. aureus strains carry both fnb genes, but there seems to be no difference in Fn-binding activity between the strains carrying one or two fnb genes. However, a study analyzing a larger collection of isolates from infected patients indicated that S. aureus strains associated with invasive disease were more likely to encode both fnb genes (Heilmann, 2011). In this study, isolates S-C3, S-C4, S-C6, S-C8, S-C9, S-F2, S-F4, S-F5, S-F6, and S-F8 carried both fnbA and fnbB genes. S-G1, S-G4, S-G8, S-G10, and S-G15 carried only the fnbA gene; and S-C10 only carried fnbB. It seems that these genes are related to the original sources through a general comparison. clumping factor A and ClfB are fibrinogen-binding proteins, which are encoded by clfA and clfB, respectively. clfA mutants failed to form clumps and adhered poorly (Heilmann, 2011). ClfB may be an important factor in S. aureus nasal colonization (O'Brien et al., 2002). In our study, the clfB gene was present in all the tested strains, while the clfA gene was only detected in four isolates (S-C3, S-C4, S-C6, and S-G10). The biofilm formations of S-C3, S-C4, and S-C6 were indeed much stronger; however, any single gene could not have a decisive influence on biofilm formation, which should be a harmonious regulation by multifactor. Additionally, the bbp gene was only detected in S-F4. The report shows Bbp is immunogenic and expressed during infection, suggesting playing a specific role in the pathogenesis of osteomyelitis, an infection of the bone, which may be caused by hematogenous spread (Vazquez et al., 2011). The strain S-F4 was isolated from a food-poisoning outbreak, which may pose a high threat to people's health. Future studies regarding the role of the MSCRAMM in disease-specific models may aid in elucidating the contribution of Bbp to S. aureus pathogenesis. The bap gene did not appear in any strains detected. Similarly, Vázquez-Sánchez et al. (2013) found that none of the S. aureus isolates from seafood carried the bap gene. In fact, the bap gene has only been identified in a small proportion of S. aureus strains originating from bovine mastitis, even though Bap was the first protein reported to be involved in S. aureus biofilm formation. To date, the clinical significance of Bap is not clear, because it is apparently present only in bovine mastitis (5% of the 350 S. aureus isolates tested) and absent from all human clinical S. aureus isolates tested so far (Cucarella et al., 2001). The pls gene was only present in S-C4 and S-C6. Pls, a surface protein of certain methicillin-resistant S. aureus strains, is a virulence factor for septic arthritis and sepsis, which implies the potential risks to humans.
Above all, considerable variations in biofilm-forming ability were observed between the different strains tested. The ratios and contributions of extracellular substances also vary greatly within strains. There was no correlation in distribution of adhesin genes and biofilm formation with morphology. Biofilm formation might be affected by many factors, which brings new challenges to the prevention of this serious pathogen due to biofilm-related infection and contamination.
Footnotes
Acknowledgments
This work was jointly supported by the National Natural Science Foundation of China (No. 31071515), the New Century Excellent Talents in University (NCET-11-0847), the National Science and Technology Major Project of the Ministry of Science and Technology of China (201203009), and the Fundamental Research Funds for the Central Universities, Southwest University for Nationalities (No. 11NZYTH08).
Disclosure Statement
No competing financial interests exist.