
Abstract
Treatment of Escherichia coli O157:H7 by certain antimicrobial agents often exacerbates the patient's condition by increasing either the release of preformed Shiga toxins (Stx) upon cell lysis or their production through the SOS response-triggered induction of Stx-producing prophages. Recommended subinhibitory concentrations (sub-MICs) of azithromycin (AZI), gentamicin (GEN), imipenem (IMI), and rifampicin (RIF) were evaluated in comparison to norfloxacin (NOR), an SOS-inducer, to assess the role of the SOS response in Stx release. Relative expression of recA (SOS-inducer), Q (late antitermination gene of Stx-producing prophage), stx1, and stx2 genes was assessed at two sub-MICs of the antimicrobials for two different strains of E. coli O157:H7 using reverse transcription–real-time polymerase chain reaction. Both strains at the two sub-MICs were also subjected to Western blotting for LexA protein expression and to reverse passive latex agglutination for Stx detection. For both strains at both sub-MICs, NOR and AZI caused SOS-induced Stx production (high recA, Q, and stx2 gene expression and high Stx2 production), so they should be avoided in E. coli O157:H7 treatment; however, sub-MICs of RIF and IMI induced Stx2 production in an SOS-independent manner except for one strain at the first twofold dilution below MIC of RIF where Stx2 production decreased. Moreover, GEN caused somewhat increased Stx2 production due to its mode of action rather than any effect on gene expression. The choice of antimicrobial therapy should rely on the antimicrobial mode of action, its concentration, and on the nature of the strain.
Introduction
The main inducers of Shiga toxin–producing prophages are DNA-damaging agents that activate the bacterial SOS response, a DNA repair system (Loś et al., 2009). The inducer RecA and the repressor LexA are the two main key regulators of this SOS response (Foti et al., 2010). Stressors that cause a direct DNA damage or a halt in DNA replication resulting in the exposure of ssDNA will activate RecA protein (Foti et al., 2010). Activated RecA then catalyzes LexA autocleavage to alleviate the repression of SOS response gene expression (Foti et al., 2010). RecA also mediates the self-cleavage of repressors of lambda and lambdoid bacteriophages such as the cI repressor of Stx-producing prophages (Galkin et al., 2009). Once the cI repressor is cleaved, a regulatory cascade of gene expression occurs to induce the Stx-producing phages and express the Stx genes. First, the immediate early genes are expressed such as N antitermination factor that permits RNA polymerase to proceed transcribing the delayed early genes that include replication, recombination, integration, and regulator genes such as Q antitermination protein (Foti et al., 2010). Finally, this Q protein allows the transcription of the late genes, which are the Stx and lysis genes (Yarnell and Roberts, 1992).
As for treatment, a regimen that avoids the risk of serious complications, such as the development of HUS, is notably absent (Panos et al., 2006). Many reports have shown that increased Stx release and disease exacerbation occur upon exposure to most antimicrobials (Panos et al., 2006; Wong et al., 2000). This led to a controversy surrounding antimicrobial therapy of this bacterial infection. This increase in Stx release by antimicrobial agents was attributed to either the release of stored Stx upon cell lysis (Shimizu et al., 2009) or to the induction of Stx-producing prophages activated by the SOS response (Kimmitt et al., 2000).
Most of the recent studies have recommended azithromycin (AZI) (Ohara et al., 2002; Zhang et al., 2009; McGannon et al., 2010), imipenem (IMI) (Kimmitt et al., 2000; Lee and Stein, 2009), rifampicin (RIF) (Rahal et al., 2011a), and gentamicin (GEN) (Kanbar et al., 2003; Mohsin et al., 2010) as potential treatments for E. coli O157:H7 infection, but warned against the use of norfloxacin (NOR), especially at its subinhibitory concentration, since it activates the SOS response, induces the Stx-producing phage, and eventually increases Stx production (Herold et al., 2005; Matsushiro et al., 1999).
Since the minimum inhibitory concentrations (MICs) and supra-MICs of antimicrobial agents eventually reach a subinhibitory concentration (sub-MIC) in vivo, we are interested in this concentration of the recommended antimicrobials. Thus, to assess the effect of sub-MICs and the role of SOS response in Stx production, an evaluation of those recommended antimicrobials at sub-MICs was done in this study in comparison to NOR, an SOS inducer.
Materials and Methods
Bacterial strains
Two strains of E. coli O157:H7 (CDC 07-98 and CDC 05-98) were obtained from the Centers for Disease Control and Prevention (CDC) that isolated them during an outbreak of HUS in the United States.
MIC determination
According to the Clinical and Laboratory Standards Institute (CLSI) guidelines (2009), the MIC of CDC 07-98 and CDC 05-98 for five antimicrobial agents was determined using the serial dilution method in Mueller Hinton II broth (Becton, Dickinson & Co., Sparks, MD). Stock solutions of the five antimicrobial agents were prepared from AZI (Sigma Chemical Co., St. Louis, MO) in 95% ethanol, GEN (Sigma Chemical Co.) in distilled water, RIF (Sigma Chemical Co.) in methanol, NOR (Fluka, USA) in distilled water, and imipenem (MSD, USA) in 0.01 mol/L phosphate buffer. Two ATCC control strains were used, which are Staphylococcus aureus 29213 and E. coli 25922. Serial broth dilution was done in triplicate for each strain, and the effect of the solvents without antimicrobial agents was also assessed.
DNA extraction and polymerase chain reaction (PCR)
DNA was extracted from the two strains using illustra bacteria genomicPrep Mini Spin Kit (GE Healthcare, UK) according to the protocol for purification of genomic DNA from Gram-negative bacteria. PCR was performed to detect the Shiga toxin genes (stx1 and stx2), lexA, recA, Q933 and Q21 genes using the primers in Table 1.
Ta, annealing temperature; qPCR, real-time PCR.
RNA extraction and cDNA preparation
The two bacterial strains were grown overnight in Mueller Hinton II broth (Becton, Dickinson & Co.). Bacterial suspensions adjusted to 107 colony-forming units (CFU)/mL of both strains were prepared with and without one of the five antimicrobial agents at two concentrations, which are twofold (sub-MIC 1) and fourfold (sub-MIC 2) dilutions of MIC. Since SOS response occurs until 60 min after its induction and then ceases (Janion, 2008), samples with and without antimicrobial agents were incubated at 37°C for 2 h with vigorous shaking and then removed to be adjusted to 1.5×108 CFU/mL. Total RNA extraction was performed on these samples using Illustra RNAspin Mini RNA Isolation kit (GE Healthcare, UK) according to the manufacturer's specifications for bacterial cells. Afterwards, cDNA was synthesized using the QuantiTect® Reverse Transcription Kit (Qiagen, Germany) according to the manufacturer's instructions from RNA with an adjusted concentration of 10 μg/mL.
Reverse transcription–real-time polymerase chain reaction (RT-qPCR)
In order to study the relative gene expression of recA, Q, stx1, and stx2 genes, RT-qPCR was performed using the QuantiFast™ SYBR® green PCR kit (Qiagen, Germany) and the BioRad CFX96 Real Time System, C1000 Thermal Cycler (Germany). Reactions using QuantiFast SYBR Green PCR master mix (Qiagen, Germany), 10 μg/mL cDNA, and 5 μM of each primer (Thermo Scientific Inc., USA), were done in duplicates for each gene in addition to the rpoB housekeeping gene. The cycling conditions were 95°C for 15 min, 95°C for 10 s, annealing temperature (Ta) for 30 s, and 72°C for 20 s, where Ta and specificity for each primer were determined using a temperature gradient and melting curve analysis (Table 1). Using the ΔΔCT equation (Livak and Schmittgen, 2001), the relative expression of the tested genes was determined in the samples with and without antimicrobial agents using the rpoB reference gene as a normalizer.
ΔΔCT= ΔCT(with antimicrobial agent) – ΔCT(without antimicrobial agent).
ΔCT(with antimicrobial agent)=CT(tested gene, with antimicrobial agent) – CT(rpoB gene, with antimicrobial agent) ΔCT(without antimicrobial agent)=CT(tested gene, without antimicrobial agent) – CT(rpoB gene, without antimicrobial agent).
Western blotting
Protein extraction was performed using bacterial lysis buffer (50 mM Tris-Cl pH=8.0, 10% glycerol, 0.1% Triton X-100, 2 mM MgCl2, lysozyme (C=100 μg/mL), DNase 3.6 U/L, and protease inhibitor) on a 0.6 optical density 600 bacterial suspension from antibiotic-free, sub-MIC 1 and sub-MIC 2 (for RIF, IMI, GEN, and NOR) samples of CDC 05-98 and CDC 07-98 that were incubated with shaking for 2 h. Protein concentrations were measured using the Bradford assay, and 20 μg were suspended in 5x Laemmli buffer. The samples were boiled for 3 min to be run on a sodium dodecyl sulfate–polyacrylamide gel electrophoresis for 1 h and then transferred to a polyvinylidene difluoride (PVDF) membrane (GE Healthcare, UK). The membrane was blocked for 1 h in 5% nonfat dry milk. After that, the membrane was incubated overnight at 4°C with the primary antibody mouse monoclonal anti-LexA, sc-7544 (Santa Cruz, USA) (diluted 1:1000 in 1% nonfat dry milk) followed by the secondary anti-mouse immunoglobulin G–horseradish peroxidase (IgG–HRP), RPN4201 (GE Healthcare, UK) diluted 1:5000. As a control, anti-L9 ribosomal protein (Gift from Dr. Isabella Moll, Max F. Perutz Laboratories, Vienna) was utilized with secondary anti-goat IgG–HRP, sc-2768 (Santa Cruz, USA) diluted 1:5000. Detection was made using ECL Western blotting detection reagents and analysis system (GE Healthcare, UK). The protein bands were visualized by autoradiography and analyzed using Image J software.
Reverse passive latex agglutination assay (RPLA) for Stx1 and Stx2
This test was utilized to detect the production of Stx1 and Stx2 upon incubation of CDC 05-98 and CDC 07-98 with and without one of the five antimicrobial agents. Supernatants of the samples, prepared similarly to those used in RNA extraction, were lyophilized to get a more concentrated solution using a freeze dryer (LABCONCO). Lyophilized supernatant was reconstituted with distilled water, and the toxin titers were determined using a VTEC-RPLA ‘SEIKEN’ Kit (Denka Seiken Co. Ltd., Tokyo, Japan) according to the manufacturer's instructions.
Statistical analysis
All statistical analyses were performed using an unpaired t-test using the GraphPad t-test calculator. Values were considered significant as compared to antibiotic-free control if p-value is <0.05.
Results
Control experiments that were done on the solvents (95% ethanol, methanol, and phosphate buffer) of the antimicrobial agents in their relative concentration showed that these diluents had no influence on the MIC. The MIC of ATCC controls were in the acceptable range according to CLSI guidelines. Results showed that both CDC 07-98 and CDC 05-98 had an MIC of 8 μg/mL for RIF, 4 μg/mL for AZI, 0.25 μg/mL for GEN, 0.125 μg/mL for IMI and 0.125 μg/mL for NOR.
PCR results showed that both CDC 05-98 and CDC 07-98 had recA, lexA, and Q933 genes while only strain CDC 05-98 had the Q21 gene. As for the Stx genes, their detection was performed at the beginning and at the end of this study to ensure that no gene loss occurred upon subcultivation. CDC 07-98 had both stx1 and stx2 genes, while CDC 05-98 only had the stx2 gene. According to Ahmad and Zurek (2006), strains with Q933 gene alone like CDC 07-98 produce more Stx than strains with Q933 and Q21 genes like CDC 05-98. RPLA results also showed that antibiotic-free CDC 07-98 produced twofold more Stx2 than antibiotic-free CDC 05-98 (data not shown). All this indicates that the strains have different characteristics.
In order to assess whether the SOS response plays a role in inducing the Stx-producing prophage in the presence of the five antimicrobial agents (AZI, RIF, IMI, GEN, and NOR), relative gene expression of recA (SOS response inducer), Q (late antitermination gene of Stx-producing prophage), stx1, and stx2 genes was performed for the CDC 07-98 and CDC 05-98 strains with and without the antimicrobial agents, and the results were normalized to a housekeeping rpoB gene as shown in Figures 1 and 2. For CDC 07-98 strain in Figure 1, sub-MIC 1 and sub-MIC 2 of NOR and AZI had higher recA, Q, and stx2 gene expression than the antibiotic-free control, but only AZI concentrations had an increase in stx1 gene. As for IMI and RIF, both concentrations showed low level of recA gene compared to the antibiotic-free control. Sub-MIC 1 of IMI along the sub-MIC 2 of IMI and RIF showed high or equal levels of stx1, Q, and stx2 genes except for sub-MIC 1 of RIF that showed low levels of those genes. As for GEN, all the genes had a lower expression in comparison to antibiotic-free control. As for the strain in Figure 2, results similar to those for CDC 07-98 were obtained except for sub-MIC 1 of RIF, which showed an increase of stx2 gene instead.
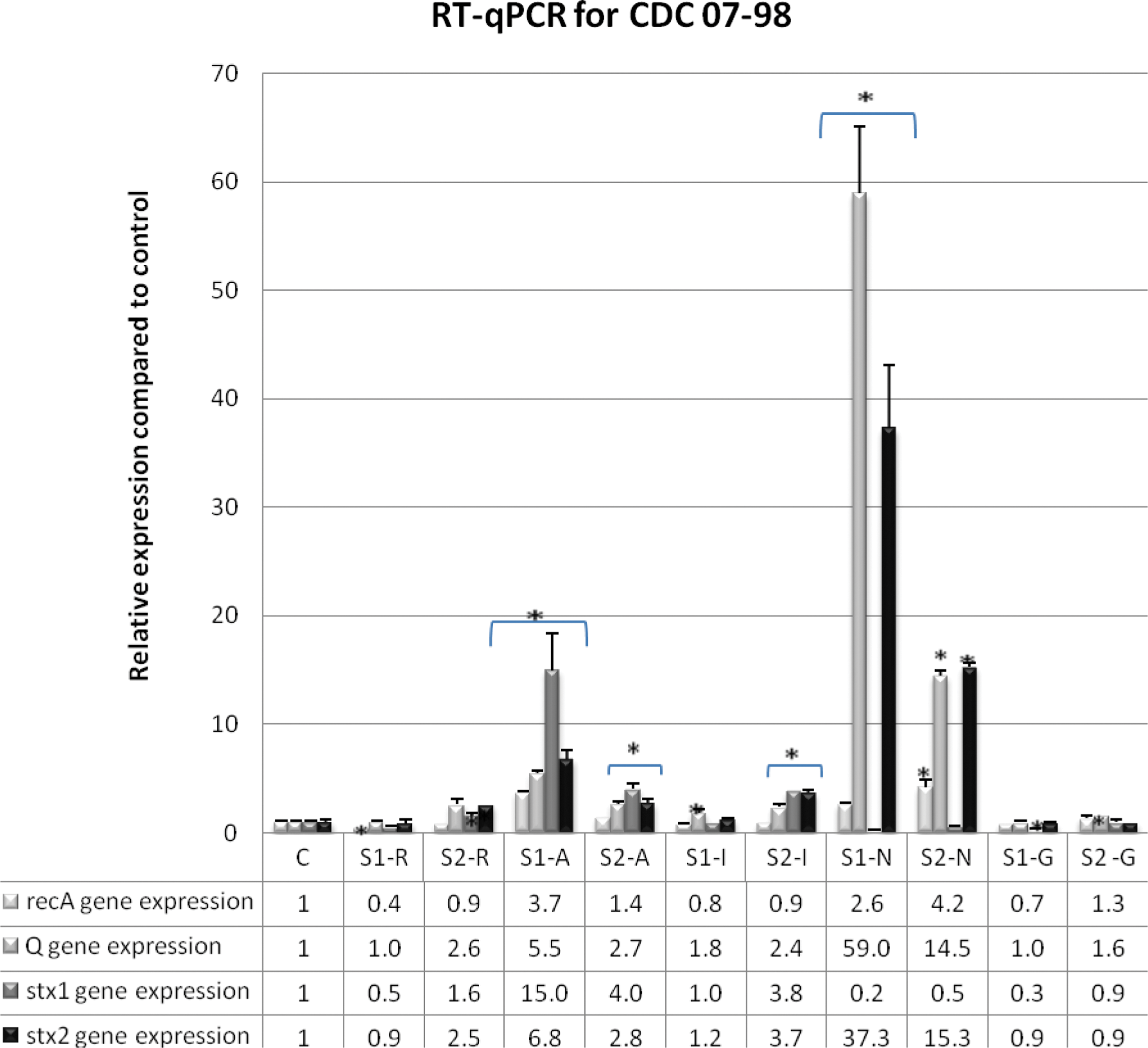
Relative expression of recA, Q, stx1, and stx2 genes in strain CDC 07-98 treated with subinhibitory concentration (sub-MIC) 1 (S1) and sub-MIC 2 (S2) of rifampicin (R), azithromycin (A), imipenem (I), norfloxacin (N), and gentamicin (G). C: Antibiotic-free sample of CDC 07-98. The relative expression of the tested genes was determined in the samples with and without antimicrobial agents and normalized to that of the rpoB reference gene. *p<0.05.
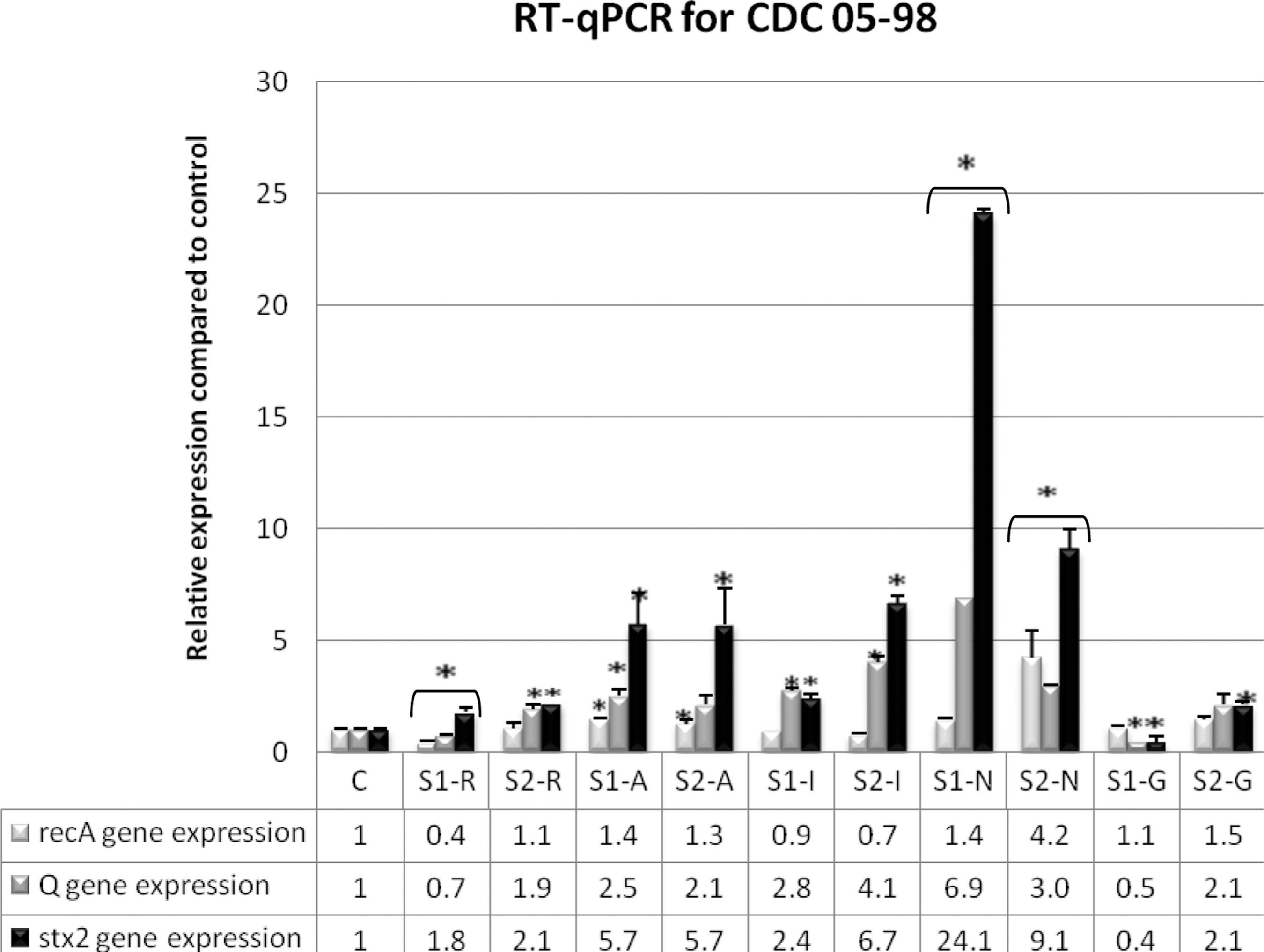
Relative expression of recA, Q, stx1, and stx2 genes in strain CDC 05-98 treated with subinhibitory concentration (sub-MIC) 1 (S1) and sub-MIC 2 (S2) of rifampicin (R), azithromycin (A), imipenem (I), norfloxacin (N), and gentamicin (G). C, Antibiotic-free sample of CDC 05-98. The relative expression of the tested genes was determined in the samples with and without antimicrobial agents and normalized to that of the rpoB reference gene. *p<0.05.
The protein expression of LexA, the SOS response repressor, was tested on CDC 07-98 and CDC 05-98 strains subjected to the antimicrobial agents to determine whether or not the SOS response is activated (Fig. 3). As compared to antibiotic-free control, LexA expression was greatly expressed only with IMI, RIF, and GEN antimicrobial agents, indicating that SOS response is not activated in contrast to NOR. Furthermore, Stx production by CDC 07-98 and CDC 05-98 strains was detected by RPLA in the presence and absence of the antimicrobial agents (Fig. 4). High Stx2 production occurred with NOR, AZI, GEN, and IMI for both strains as compared to the control. As for RIF, Stx2 production was equal to or twofold higher than the control except for sub-MIC 1 of CDC 07-98, which showed a decrease. As for Stx1, its production by CDC 07-98 decreased with sub-MIC 2 of RIF, AZI, IMI, and the two concentrations of GEN.

Western blotting results for strains CDC 07-98 referred to as “a” and CDC 05-98 referred to as “b” treated with subinhibitory concentration (sub-MIC) 1 (S1) and sub-MIC 2 (S2) of rifampicin (R), imipenem (I), norfloxacin (N), and gentamicin (G). LexA was detected in each sample using specific monoclonal antibodies for LexA (anti-LexA), and a loading control ribosomal L9 was detected using specific antibodies for L9 (anti-L9). Ca, antibiotic-free sample of CDC 07-98; Cb, antibiotic-free sample of CDC 05-98.
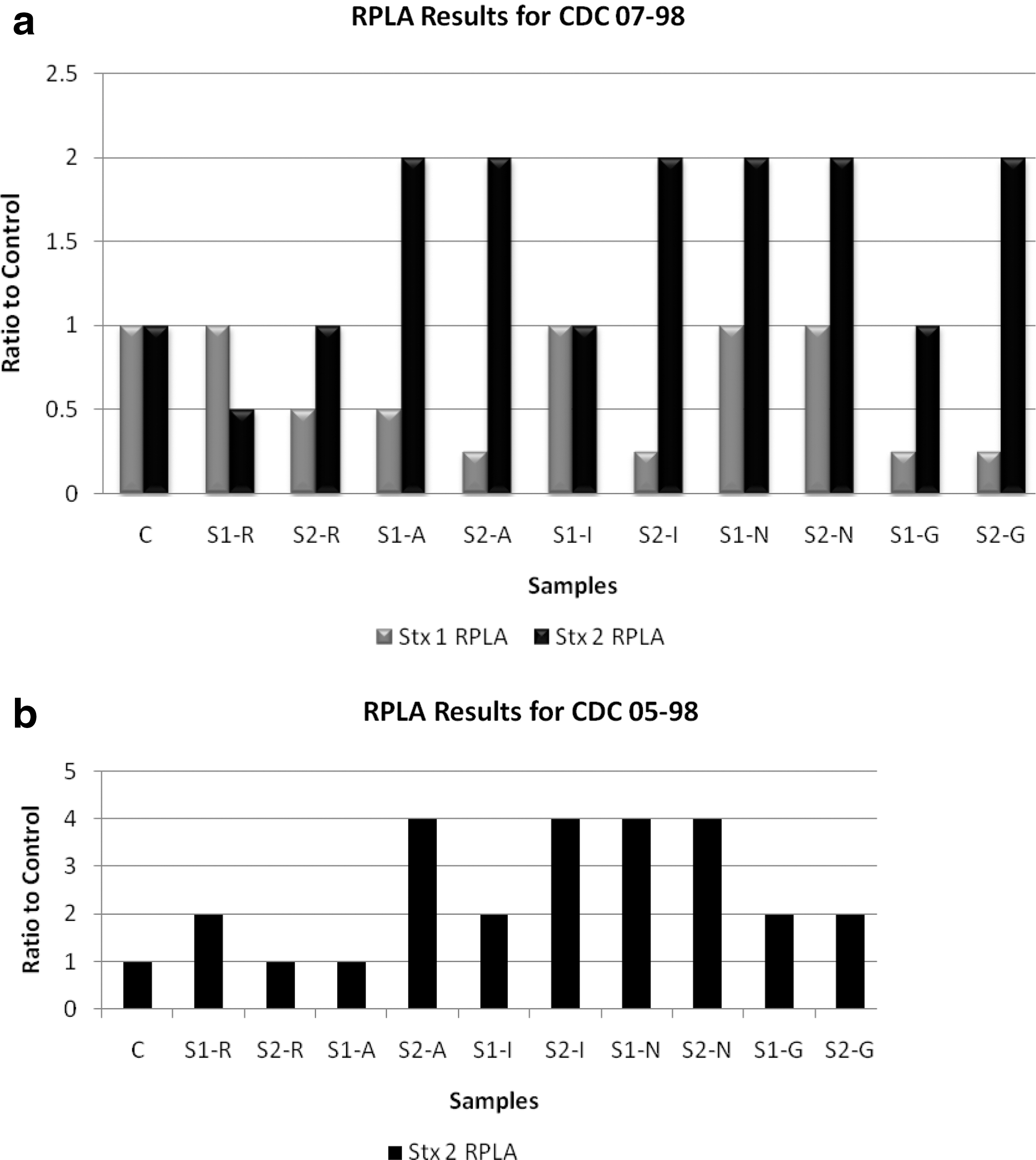
Reverse passive latex agglutination assay (RPLA) for Stx1 and Stx2 in supernatants from Escherichia coli O157:H7 cultures treated with subinhibitory concentration (sub-MIC) 1 (S1) and sub-MIC 2 (S2) of rifampicin (R), azithromycin (A), imipenem (I), norfloxacin (N), and gentamicin (G).
Discussion
NOR and AZI are SOS inducers
For CDC 07-98 and CDC 05-98, all sub-MICs of NOR and AZI caused an increase in recA, Q, and stx2 gene expression compared to the antibiotic-free samples (Figs. 1 and 2). As for LexA, Western blotting showed a decrease in its expression at both sub-MICs of NOR for both strains (Fig. 3). Moreover, compared to the antibiotic-free samples, RPLA results revealed an increase in Stx2 release for sub-MICs of NOR and AZI for both strains (Fig. 4). Hence, NOR and AZI activated the SOS response and increased Stx2 production. Consequently, these antimicrobials should be avoided in the treatment of E. coli O157:H7.
Our results were in accordance with what is reported to be NOR's mode of action. NOR is a fluoroquinolone that acts as a DNA-damaging agent by inhibiting the A subunit of DNA gyrase and eventually inhibiting DNA synthesis. In a sequential manner, NOR can activate the SOS response, induce Stx2 prophages, increase stx2 gene expression, and subsequently increase Stx2 production (Matsushiro et al., 1999). Other studies have shown that at the transcriptional level, the subinhibitory concentration of NOR causes upregulated expression of prophage genes in addition to recombination and stress genes. On the other hand, it leads to the downregulation of bacterial primary metabolism genes such as those involved in energy production, cell division, and amino acid biosynthesis (Herold et al., 2005). As for AZI, this antimicrobial agent acts by reversibly binding to the 50S ribosomal subunit and inhibiting the RNA-dependent protein synthesis. Even though AZI has been observed to decrease Stx2 in certain reports (Zhang et al., 2009; McGannon et al., 2010), our results showed increased Stx2 production similar to that reported by Grif et al. (1998) but not as much as NOR. This could be dependent on the Stx2 toxin subtype, since Pedersen et al. (2008) demonstrated variability in the suppression of Stx2 release upon exposure of different Stx2 subtype producing E. coli O157:H7 to sub-MICs of AZI.
RIF and IMI produce Stx2 by a non-SOS-dependent mechanism
RIF and IMI did not activate the SOS response as implied by the low or equal recA gene expression (Figs. 1 and 2) and the moderate-to-high LexA expression as compared to the controls at both sub-MICs for both strains (Fig. 3). However, as compared to the control, in general both antimicrobials managed to increase Q and stx2 gene expression (Figs. 1 and 2) and to have an equivalent or increase in Stx2 release (Fig. 4), which denotes the potential Stx2 prophage induction to produce Stx2. This occurred in all sub-MICs of RIF and IMI for both strains except with sub-MIC 1 of RIF for CDC 07-98, where low Q gene expression occurred (Fig. 1) along with decreased Stx2 release as compared to the control (Fig. 4a). Moreover, the stx2 gene expression and its release of CDC 05-98 in response to IMI concentrations was more virulent (higher fold) than CDC 07-98, but the stx2 gene expression and its release of both bacteria in response to RIF was less virulent (equal to low fold) as compared to control and to all the other antimicrobial agents used, denoting that RIF could be safely used in the treatment of this bacterium (Figs. 1, 2, and 4).
Even though RIF and IMI did not activate the SOS response, Stx2 production could be due to a mechanism other than the SOS activation of Stx2 prophage. Rokney et al. (2008) found that the lambda phage, which is similar to the Shiga toxin–producing prophage, can be induced by the activation of the heat-shock stress response in a pathway that requires the Q protein but is quite different from that of the SOS response. Even though RIF inhibits RNA transcription, it has been reported that this inhibition occurs mainly at σ70 (primary σ)-dependent promoters more than σ32 (responsible for heat-shock genes)-dependent promoters (Wegrzyn et al., 1998), thus allowing the transcription of heat-shock genes. Knowing that some antimicrobial agents were reported to activate the heat-stress response (Foster, 2005), RIF could also activate this response, which could eventually induce Stx-producing prophages and produce Stx2.
Moreover, an inhibition of Stx2 release occurred on a specific strain at the higher concentration of RIF, which is closer to MIC, which suggests that the inhibitory effect of RIF relies on the nature of the strain and the concentration of the agent. The differential response of E. coli O157:H7 strains to subinhibitory concentrations of antimicrobial agents was also shown by Grif et al. (1998), where Stx2-producing strains showed different results than Stx1- and Stx2-producing strains upon exposure to 13 antimicrobial agents. A recent study that assessed the use of RIF rifampicin in treating E. coli O157:H7 in vitro and in vivo showed that treatment with the MIC of rifampicin followed by its minimum bactericidal concentration managed to cause an effective decrease in Stx toxins in vitro and to enhance the survival of infected Balb/c mice in vivo (Rahal et al., 2011b).
Gentamicin releases Stx2 in spite of SOS response inactivation
At both sub-MICs with both strains, GEN failed to activate the SOS response as shown by the lowered recA gene expression (Figs. 1 and 2) and high LexA expression (Fig. 3). Even though GEN showed low Q and stx2 gene expression for both strains (Figs. 1 and 2), RPLA demonstrated a double or equal Stx2 production as compared to the control (Fig. 4). This could be due to the fact that the cationic GEN at its sub-MIC concentration not only can inhibit protein translation but can also perturb the packing order of lipids, thereby destabilizing bilayered membranes and liberating preformed Stx2 found in membrane vesicles (Kolling and Matthews, 1999). This has been observed with Pseudomonas aeruginosa, where GEN destabilized the membrane and increased the release of membrane vesicles three- to fivefold (Kadurugamuwa and Beveridge, 1997).
Stx1 showed different results compared to Stx2 with all antimicrobials
For CDC 07-98, even though Stx1 and Stx2 are phage encoded, the expression level of stx2 and stx1 genes is not the same, especially with NOR that causes a high stx2 gene expression but low stx1 gene expression. Moreover, even when an increase in stx1 gene expression occurs, in contrast to Stx2, Stx1 release into the culture was not observed. For instance, high stx1 gene expression occurred with sub-MICs of AZI and sub-MIC 2 of IMI (Fig. 1), but RPLA results showed that Stx1 release from all sub-MIC samples was equal to or lower than the antibiotic-free control (Fig. 4). The difference in the results of Stx1 and Stx2 release could be due to their different translocations across the outer membrane. Stx1 is almost completely cell associated, whereas Stx2 is found in the extracellular fraction (Shimizu et al., 2009). Another possible explanation could be that if the transcription of the stx1 gene initiates at the Stx1 promoter, upstream of the phage lysis genes, these lysis genes would not be sufficiently expressed to allow the release of the toxin (Shimizu et al., 2009).
Conclusions
In conclusion, Stx2 production by two distinct strains of E. coli O157:H7 can occur in an SOS-dependent and -independent manner at the sub-MICs of NOR, GEN, AZI, IMI, and RIF except for sub-MIC 1 of RIF for CDC 07-98. Hence, the choice of antimicrobial therapy for E. coli O157:H7 should avoid the release of Stx by taking into consideration the antimicrobial mode of action, its concentration, and the nature of the strain. Further studies should be done on different strains to assess the effect of RIF on Shiga toxin release, since this antimicrobial agent may constitute a potential agent for treatment.
Footnotes
Disclosure Statement
No competing financial interests exist.