
Abstract
The aim of this study was to determine the influence of acid-adaptation on the survival as well as adhesion and invasion of human intestinal cells by nine Campylobacter jejuni strains after exposure to different stress conditions. Acid-adapted and nonadapted C. jejuni were exposed to different secondary stress conditions such as acid (pH 4.5), starvation (phosphate-buffered saline, pH 7.2), or salt (3% wt/vol NaCl). After exposure to the secondary stress, the adhesion and invasion abilities of the strains were evaluated in vitro in tissue culture using the human intestinal cell line INT 407. The survival rates of acid-adapted cells of some strains of C. jejuni exposed to different secondary stresses were found to be significantly higher than the non-acid-adapted cells. Similarly, some strains also showed an increase in adhesion and invasion (p<0.05) when acid-adapted C. jejuni were exposed to stresses such as acid, starvation, or salt as compared to non-acid-adapted C. jejuni. We found that adaptation to acid stress can enhance the survival of C. jejuni when exposed to secondary stresses and, thus, result in increased adhesion and invasion of human intestinal cells in vitro. However, the survival rates as well as the degree of adhesion and invasion were found to vary with the strain of C. jejuni, the time of adaptation to acid, the type of the secondary stress and exposure time to the secondary stress. These results show that adaptation to stresses could influence virulence of C. jejuni. Understanding the conditions by which C. jejuni adapts to stresses will provide information concerning how this organism is able to survive inside and outside the host. This, in turn, could offer methods to reduce or eliminate C. jejuni in the environment.
Introduction
Many virulence factors have been proposed in the survival of C. jejuni and establishment of the disease in humans. Interaction of C. jejuni and the host cells is a complex process involving cell surface structures of the bacteria and the host cell receptors (Rubinchik et al., 2012). C. jejuni may cause disease either by invasion of the intestinal epithelial cells and subsequent changes to host cells, production of toxins by the ingested and colonized C. jejuni, or damage to extra-intestinal sites by C. jejuni passing through the lymphatic vessels (Zheng et al., 2006). Depending on the status of the host and the strain involved in infection, one or more of the above-described mechanisms may be involved. Invasion of intestinal epithelial cells was found to be reduced when C. jejuni was exposed to heat shock and starvation, whereas exposure to atmospheric oxygen increased the invasion ability (Mihaljevic et al., 2007). Exposure to oxygen was also found to increase the invasion of murine macrophages by C. jejuni as well as increase their intracellular survival when compared to other stresses such as starvation and heat shock (Pogacar et al., 2009). To the best of our knowledge, the effect of stress-adaptation on the adherence and invasion of C. jejuni when exposed to further secondary stresses has not been studied. So the objectives of this study were as follows: (1) to determine whether acid-adaptation in C. jejuni increased the survivability when the bacteria are further exposed to secondary stresses such as acid (pH 4.5), starvation (phosphate-buffered saline [PBS], pH 7.2), or salt (3% wt/vol NaCl); and (2) to compare any changes in adhesion and invasion of INT 407 cells by C. jejuni, with and without acid-adaptation, after exposure to secondary stresses.
Materials and Methods
Bacterial strains
A total of nine C. jejuni strains, isolated from humans and poultry, were used for this study based on their distinct pulsed-field gel electrophoresis profiles (data not shown). The six human isolates included 81-176, NCTC 11168, and four isolates 70 H, 71 H, 87 H, and 1023 H obtained from humans diagnosed with campylobacteriosis. The three poultry isolates used were obtained from different stages of processing including prechilled (PRCC 3), postchilled (POCC 13), and retail (RECC 3) chicken carcass isolates. Frozen stock cultures were passed twice on Campylobacter enrichment (CE) blood agar (Acumedia®) plates and then inoculated into CE broth and incubated in a micro-aerobic atmosphere consisting of 5% O2, 10% CO2, and 85% N2 at 42°C for 18 h to obtain the early stationary-phase cultures for the experiments.
Survivability of acid-adapted C. jejuni exposed to different secondary stresses
Early stationary phase (18 h) C. jejuni cultures in CE broth were divided into two portions and centrifuged at 8000×g for 5 min and subsequently resuspended in either acid broth (pH 5.5) to obtain acid-adapted cells or in CE broth (pH 7.2) to obtain cells not acid stressed. Acid broth was prepared by adding hydrochloric acid (HCl) directly to the CE broth and pH measured using a pH meter. After an adaptation time of 2 h at 42°C under microaerophilic conditions, both acid-adapted cells and non-acid-adapted cells were centrifuged and exposed to the following stresses for a period of 2 h: (1) acid stress (4.5) by resuspending bacterial pellets from both the treatment groups in CE broth with a pH of 4.5; (2) starvation stress by resuspending acid-adapted and non-acid-adapted bacterial cell pellets in PBS containing no nutrients and with a pH of 7.2; (3) salt stress by resuspending bacterial pellets from both treatment groups in CE broth to which sodium chloride was added to give a final concentration of 3% NaCl (wt/vol). All the treatment groups were incubated for 2 h at 42°C under microaerophilic conditions. Postexposure to the secondary stresses surviving culturable C. jejuni was evaluated by standard plating methods. The survival percentages were calculated by dividing the colony-forming units (CFU)/mL after acid adaptation and exposure to secondary stresses by the CFU/mL at time zero and multiplying by 100. All the experiments were repeated three times. We also examined the effects of a 3 h acid-adaptation on the survivability of four C. jejuni isolates when further exposed to acid stress (pH 4.5).
Cell cultures
The INT 407 cells (human embryonic intestinal cell line ATCC CCl-6) were grown in Basal Medium Eagle (BME) (Sigma Chemical Co., St. Louis, MO) with 10% fetal bovine serum (Sigma) and 2 mM
Adhesion and invasion assays
Adhesion and invasion assays were performed as previously described using INT 407 cells (Gilbert and Slavik, 2005). Briefly, INT 407 cells grown at the rate of 2×105 cells per well on 24-well tissue culture plates for 24 h were washed three times with sterile PBS. Acid-adapted and non-acid-adapted C. jejuni cells were exposed to further secondary stresses as described in objective 1 and subsequently washed twice with PBS, centrifuged, and resuspended in Basal Medium Eagle without antibiotics before challenging the intestinal cells. The multiplicity of infection (MOI) of the stress-adapted or non-stress-adapted bacterial cultures exposed to secondary stresses was assessed by serial dilution and plating and not by spectrophotometry as this may include the dead cells also. Plates then were incubated for 2 h to allow adherence and invasion of tissue culture cells. After 2 h, all plates were washed three times with PBS to remove any nonadhering bacteria. One set of plates was treated with PBS plus 0.1% Triton X-100 and gently swirled for 30 min at room temperature (25°C–27°C) to lyse the intestinal cells, and the total number of bacteria associated with the intestinal cells (invading and adhering) were then determined by serial dilution and plating on CE blood agar plates. To find out the number of invading bacteria, the second set of plates after the incubation period of 2 h and washing with PBS three times was treated with BME media containing 100 μg/mL of gentamicin and incubated for an additional 2 h. Following this step, the intestinal cells were lysed using the same method as described above, and the number of released internalized bacteria were determined by serial dilution and plating on CE blood agar plates. The total number of adherent bacteria was determined by using the formula: number of bacteria in non-gentamicin-treated culture (adherent and invasive) – number of bacteria in gentamicin-treated culture (invasive)=number of adherent bacteria. Results of adhesion/invasion are expressed in percentages as the MOI were found to vary (100 for the secondary stresses of acid and starvation and 10 for the salt stress). The percentage of adhesion/invasion was calculated by dividing the number of adherent/invading bacteria by the total number of bacteria used for challenging the intestinal cells and multiplying by 100.
Statistical analysis
The experiments were conducted in three independent replicates and were analyzed by a one-way analysis of variance and Student t-test using the JMP statistical software package 9.0.2. The results were considered statistically significant with p-values reported at p≤0.05.
Results
Survivability of acid-adapted C. jejuni exposed to different secondary stresses
Survivability of C. jejuni strains were assessed in this study by determining the survival percentages of viable and culturable C. jejuni cells after acid-adaptation and exposure to the secondary stresses of acid, starvation, or salt. Non-acid-adapted cells exposed to the secondary stresses served as the control group. Acid-adapted cells exposed to the secondary stress of acid for 2 h were found to exhibit a significantly higher survival percentage (p<0.05) than their respective controls for the isolates 81-176, NCTC 11168, 87 H, PRCC 3, and RECC 3 (Table 1). However, the isolates 70 H, 71 H, and 1023 H showed no significant differences in survival percentages between treatment and control groups. Increasing the acid adaptation time to 3 h and exposing to the secondary stress of acid was found to increase the survival percentages of acid-adapted cells for only two of the isolates, POCC 13 and RECC 3 (Table 2).
Letters indicate significant differences (p≤0.05) between groups (acid-adapted or non-acid-adapted).
PBS, phosphate-buffered saline.
Letters indicate significant differences (p≤0.05) between groups (acid-adapted or non-acid-adapted).
Similarly, after acid adaptation when C. jejuni cells were exposed to a secondary stress of starvation for 2 h, five of the nine isolates showed higher survival percentages for the acid-adapted cells than their respective controls. These isolates were 81-176, NCTC 11168, 70 H, PRCC 3, and POCC 13 (Table 1). A secondary stress of salt was, however, found to increase the survival percentages of acid-adapted cells for only two of the isolates, POCC 13 and RECC 3 (Table 1). These results show that an acid-adaptation helps the early stationary phase cells of C. jejuni to enhance their survivability after exposure to secondary stresses that varied among the isolates used in our study.
Effects of acid adaptation of C. jejuni on adhesion and invasion of INT 407 cells
A plate-count assay was used to determine the adhesion to and invasion of INT 407 cells by C. jejuni after acid-adaptation and exposure to secondary stresses including acid, starvation, or salt. Non-acid-adapted cells exposed to further stresses served as the control group. Some acid-adapted C. jejuni isolates were found to have increased adhesion and invasiveness, but this varied with the strains, the time of adaptation of acid, and the time of exposure to the secondary stress. With a 2-h acid-adaptation, the isolates PRCC 3, NCTC 11168, and 87 H showed increased adhesion as compared to their controls with no acid-adaptation when exposed to the secondary stress of acid. However, for the human isolate, 71 H, non-acid-adapted controls exhibited higher adhesion rates (Fig. 1). When the acid-adaptation time was increased to 3 h, two isolates—POCC 13 and RECC 3—were found to show significant increases in adhesion compared to their control groups after exposure to further acid stress (Table 2).
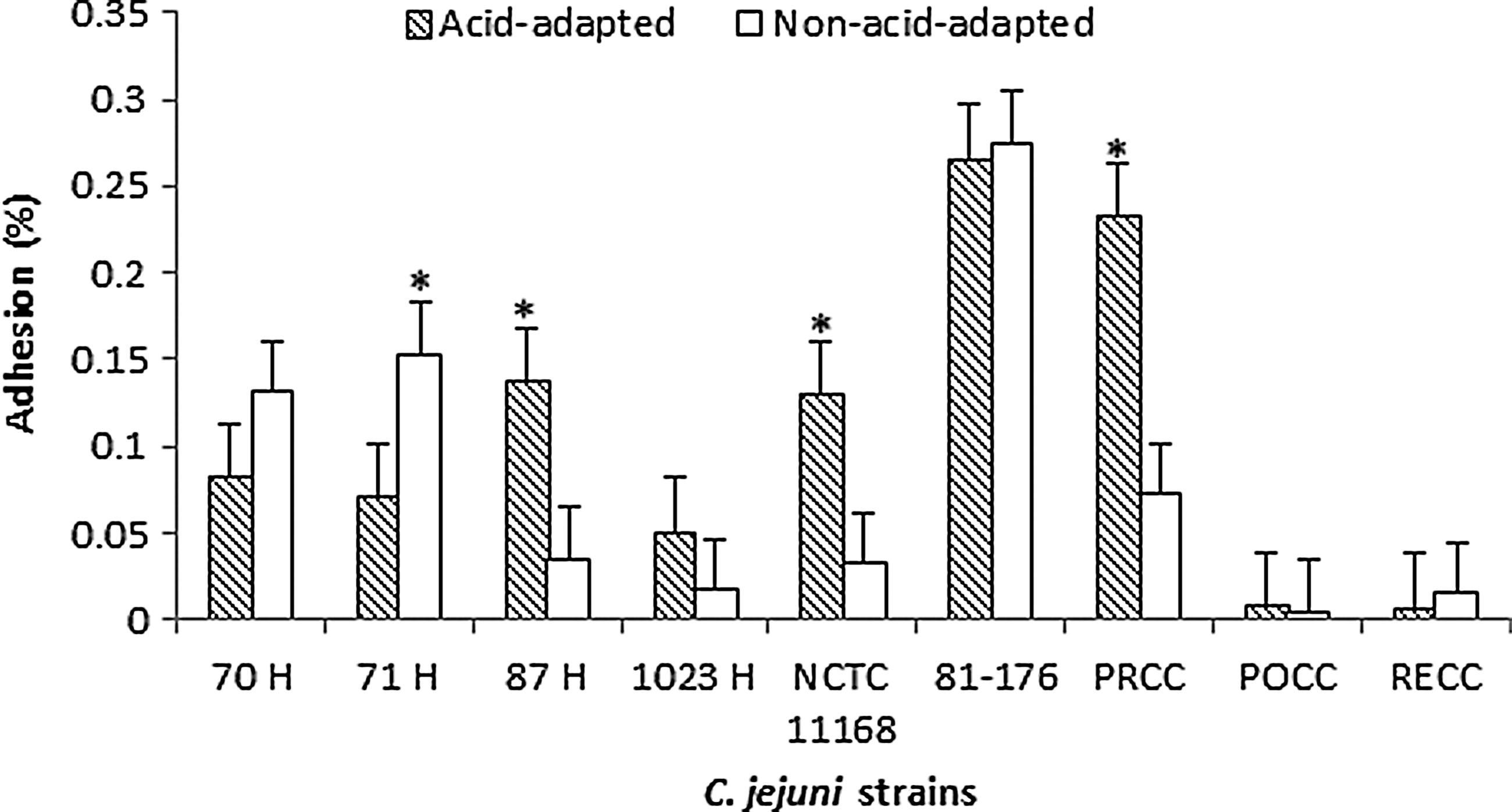
Adhesion of acid-adapted (CE broth, pH 5.5) and non-stressed (CE broth, pH 7.2) Campylobacter jejuni strains with a 2-h adaptation time when exposed to secondary stress of acid pH of 4.5 for a period of 2 h. The results are presented as percentages±standard deviation (*, p≤0.05).
Acid-adapted cells of C. jejuni after exposure to a secondary stress of starvation also showed an increase in adhesion for the isolates PRCC 3, POCC 13, 81176, and NCTC 11168, whereas the non-acid-adapted cells of the human isolate 87 H were found to have significantly higher adhesion and invasion (Fig 2). Similarly, when acid-adapted C. jejuni were exposed to a secondary stress of salt, only two of the isolates (POCC 13 and RECC 3) were found to have significantly higher rates of adhesion than the controls. On the contrary, however, the isolates 81-176 and PRCC 3 were found to exhibit higher adhesion rates for the control group (Fig. 3). The rest of the isolates did not show any difference between the two treatment groups. After exposure to the secondary stresses, we found similar observations in the invasion rates of the strains (Table 3). Invasiveness of the strains was dependent on their adhesion, with higher invasion percentages seen for those strains with higher adhesion rates.
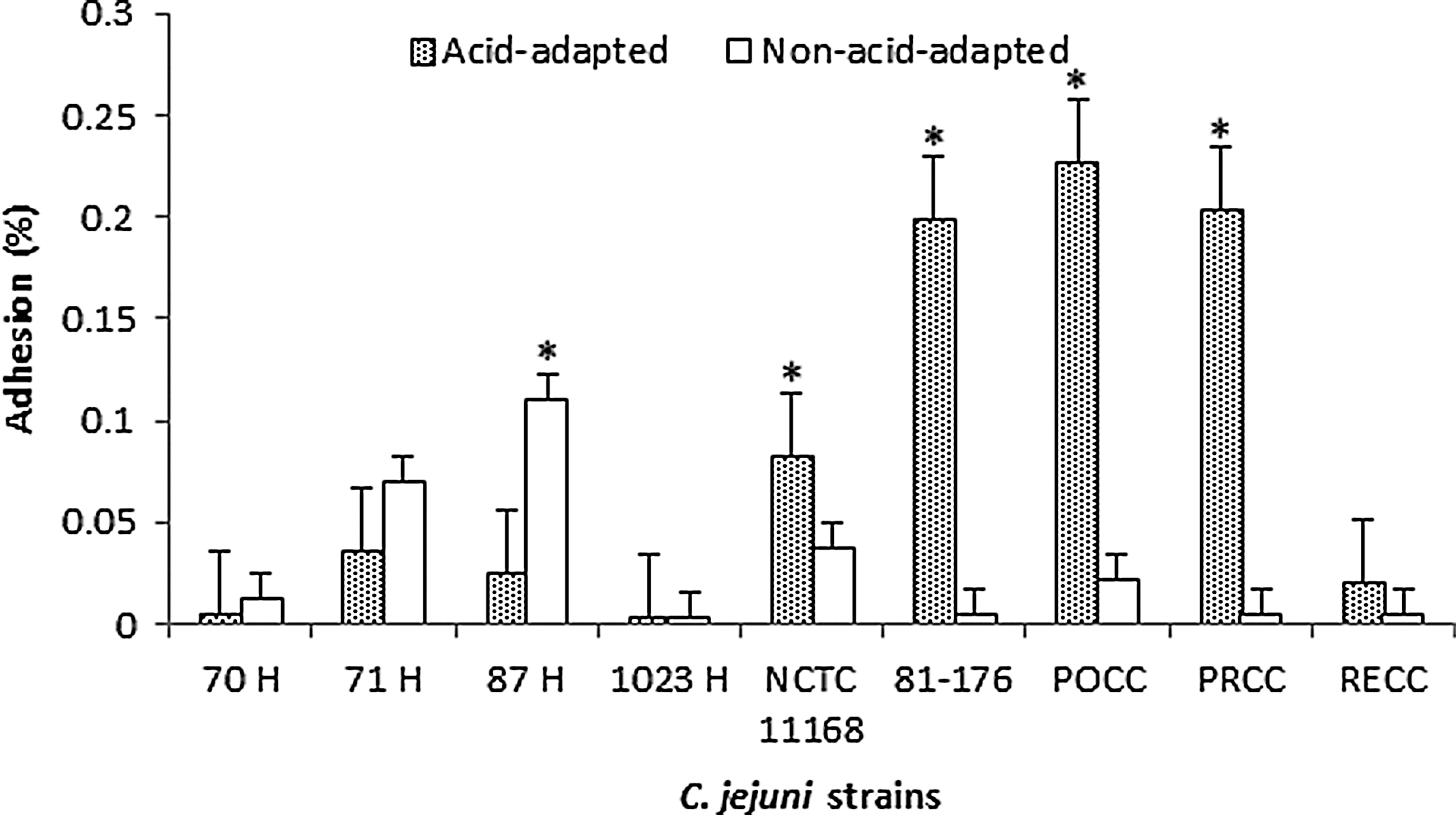
Adhesion of acid-adapted (CE broth, pH 5.5) and non-stressed (CE broth, pH 7.2) Campylobacter jejuni strains with a 2-h adaptation time when exposed to secondary stress of starvation (phosphate-buffered saline, pH 7.2) for a period of 2 h. The results are presented as percentages±standard deviation (*, p≤0.05).
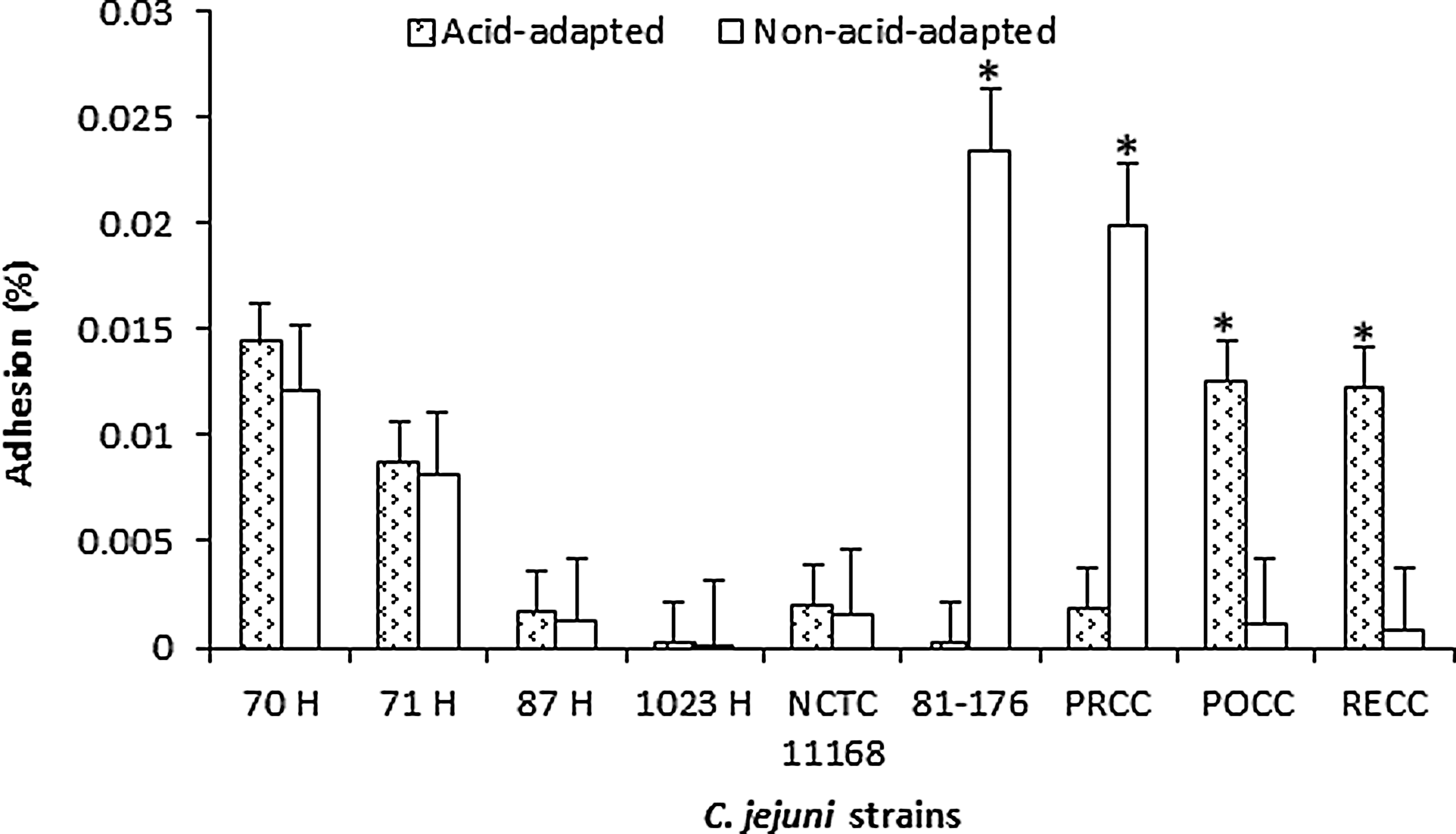
Adhesion of acid-adapted (CE broth, pH 5.5) and non-stressed (CE broth, pH 7.2) Campylobacter jejuni strains with an adaptation time of 2 h when exposed to a secondary stress of 3 % (wt/vol) NaCl for a period of 2 h. The results are presented as percentages±standard deviation (*, p≤0.05).
Letters indicate significant differences (p≤0.05) between groups (acid-adapted or non-acid-adapted).
Discussion
Adaptation to stress is among the survival methods for pathogenic organisms including foodborne bacteria (Ray and Bhunia, 2008). Recent studies have shown the effects of various stress conditions without any stress adaptation on the survival of C. jejuni in vitro in various tissue culture cell lines (Pogacar et al., 2009 and 2010). Research also shows that C. jejuni has the capacity to sense and respond to acid stress very well. Studies by Le and coworkers (2012) showed that the exposure of C. jejuni strain NCTC 11168 to an acid pH of 5.0 without any prior stress adaptation was found to increase its invasion of mouse intestinal crypt cells. They found that on exposure to acid, C. jejuni was down-regulating genes involved in cell division and replication and up-regulating flagellar and stress response genes. However, to our knowledge, there is no previous record on the adhesion and invasion abilities of C. jejuni after acid-adaptation. The results of our study indicate that C. jejuni is capable of inducing an ATR upon exposure to an acidic pH of 5.5, which may increase the survival rates when exposed to secondary stresses including acid (CE broth pH 4.5), starvation (PBS, pH 7.2), or salt (3% wt/vol NaCl). It was also found that the adhesion/invasion of C. jejuni to human intestinal cells (INT 407) were dependent on the survival percentages. However, we found variations among the strains used in our study, indicating that only some strains have the ability to induce an ATR under the conditions used in our study. The induction of an ATR, as well as adhesion and invasion abilities, were found to vary not only with the strain of C. jejuni but also with the time of exposure to a sublethal pH for adaptation, time of exposure to the secondary stress, as well as the type of secondary stress. The adhesion and invasion rates also might be dependent on the survival percentages of C. jejuni surviving further secondary stresses. Studies have shown variations among the strains of C. jejuni in the survival ability to survive further stresses after adaptation to a sublethal stress (Murphy et al., 2003; Shaheen et al., 2007). The isolates selected for this study, however, had similar adhesion and invasion rates under normal growth conditions as shown by previous studies in our laboratory (Gilbert and Slavik, 2005). However, we found differences in acid-adaptation between the poultry isolates and isolates from patients diagnosed with campylobacteriosis, which might be due to a previous acid-adaptation of the human isolates in acidic conditions of the stomach.
In our research, we found that an acid-adaptation helps C. jejuni to increase survivability as well as adhesion and invasion of intestinal epithelial cells when exposed to secondary stresses such as acid, starvation, and salt. Our results are consistent with the results of previous research conducted with other foodborne bacterial pathogens. Salmonella Typhimurium was shown to induce an ATR following acid-adaptation at pH 5.5, which provided protection to the bacteria when subsequently they were exposed to salt stress (Greenacre and Brocklehurst, 2006). Virulence, as determined by invasion of acid-adapted Salmonella Typhimurium in INT-407 cells, was found to increase on further exposure to an acidic pH of 3.8 (Wilmes-Riesenberg et al., 1996). An adaptation to acid or salt and subsequent exposure to acid or salt stress was also found to increase the virulence potential of Escherichia coli strains as measured by adhesion to various tissue culture cells (House et al., 2009; Olesen and Jespersen, 2010). Multiple stress response genes were found to be involved in adaptation and regulation of virulence factors of C. jejuni (Reid et al., 2008). Recent studies show that the transcription of virulence and stress response genes in C. jejuni was induced by the temperature shifts associated with food storage and was found to vary with the strains (Poli et al., 2012).
The induction of an ATR could possibly help C. jejuni to survive stresses in the food and poultry processing environment. C. jejuni also has to survive the acidic conditions in the human stomach to cause the disease. A very low infectious dose of 500 cells or less for C. jejuni (Black et al., 1988) suggests that this organism is capable of survival in an acidic environment. The virulence of pathogenic bacteria depends on their adaptation to and survival in the stressful conditions encountered within the host (House et al., 2009). C. jejuni has been shown to have evolved specific adaptations to survive in the host cells. One such adaptation is the survival of C. jejuni in intestinal epithelial cells in vacuoles that were found to deviate from the endocytic pathway and, thus, avoid delivery into lysosomes (Watson and Galán, 2008). Furthermore, it has been shown that a brief exposure to atmospheric oxygen for 5 h without prior adaptation to any type of stress can increase the survival of C. jejuni in murine macrophages and pig intestinal epithelial cells (Pogacar et al., 2009 and 2010). Adaptation to acid and sodium chloride stress and further exposure to stresses was found to increase the invasion ability of Listeria monocytogenes in Caco-2 cells (Olesen et al., 2009). The results from this study also indicate that adaptation to sublethal stresses may increase the virulence of C. jejuni by increasing the adhesion and invasion abilities. Other factors, such as production of stress proteins, as well as upregulation of virulence genes, might also be involved in Campylobacter pathogenesis and will warrant further study.
Conclusions
Acid-adaptation and exposure to secondary stresses such as acid, starvation, and salt was found to affect the survival as well as adhesion and invasion abilities of C. jejuni. Because these studies have demonstrated a variation among C. jejuni isolates in response to stress, caution should be taken when applying the results of this study to other C. jejuni strains.
Footnotes
Acknowledgments
The authors would like to thank the Arkansas Bioscience Institute (ABI) for funding this project. We thank Drs. Donald Cave, Mike Johnson, and Qijing Zhang for providing the Campylobacter jejuni strains used in this study. We also thank Dr. Jody Lingbeck for the cell lines and assistance with the tissue culture portion of this study.
Disclosure Statement
No competing financial interests exist.