
Abstract
Bacillus cereus is capable of producing enterotoxin and emetic toxin, and Bacillus foodborne illnesses occur due to the consumption of food contaminated with endospores. The objectives of this study were to investigate the growth and toxin production of B. cereus in cooked rice and to determine the effect of temperature on toxin destruction. Cooked rice inoculated with B. cereus was stored at 15, 25, 35, and 45°C or treated at 80, 90, and 100°C. The results indicated that emetic toxin was produced faster than enterotoxin (which was not detected below 15°C) at all the storage temperatures (15–45°C) during the first 72 h. Emetic toxin persisted at 100°C for 2 h, although enterotoxin was easily to be destroyed by this treatment within 15 min. In addition, B. cereus in cooked rice stored at a warm temperature for a period was not inactivated due to survival of the thermostable endospores. These data indicate that the contaminated cooked rice with B. cereus might present a potential risk to consumers. Results from this study may help enhance the safety of such food, and provide valuable and reliable information for risk assessment and management, associated with the problem of B. cereus in cooked rice.
Introduction
B
B. cereus diarrheal-type food poisoning was first identified after an investigation of a hospital outbreak in Norway, while B. cereus emetic syndrome was originally identified in the early 1970s as a result of consumption of cooked rice in Chinese restaurants in the United Kingdom (Hauge, 1955; Schoeni and Wong, 2005). Prevalence of foodborne-disease outbreaks due to B. cereus is increasing in industrialized countries. From 1980 to 1996, B. cereus was reported to cause 3.9% of the cases of foodborne illnesses in Australia (Cameron, 1997). About 16.5% of the cases of foodborne illnesses were attributed to B. cereus from 1993 to 1998 in The Netherlands (Tirado and Schmidt, 2001). In 2007, 19 outbreaks resulted in 164 cases of foodborne illnesse were attributed to B. cereus in the United States (CDC, 2010). Also in the United States, about 52.8% of 178 samples of raw rice from retail food stores were found to be contaminated by B. cereus spores (Ankolekar et al., 2009).
Cooked rice is one of the main foods associated with B. cereus outbreaks in many countries (Lake et al., 2004; Little et al., 2002), including China, Japan, and Korea. Approximately 95% of B. cereus food-poisoning cases with emetic symptoms have been associated with fried or cooked rice (Hui and Khachatourians, 1994). In Sri Lanka, 56.0% of 200 fried-rice samples were found to be positive for B. cereus (Perera and Ranasinghe, 2012). In Belgium, 18.5% of 54 rice dish samples in restaurants were found to be contaminated by B. cereus strains (Delbrassinne et al., 2012). In Korea, 25% of 293 cereal samples were positive for B. cereus, and all of the isolates had diarrheal toxin genes (Park et al., 2009). From 2001 to 2008, a total of 27 outbreaks of foodborne illnesses related to the consumption of food contaminated by B. cereus were reported by the Korea Food and Drug Administration, including 13 cases of B. cereus diarrhea, from 2001 to 2007 (Kim et al., 2010a).
Korean traditional foods such as kimbab (made from cooked rice and rolled in dried seaweed) and tteok (a class of Korean traditional rice cakes) are typically stored at room temperature, since refrigeration temperatures would significantly reduce their quality. Many consumers may eat cooked rice that has been stored at or above room temperature for a period in a restaurant, cafeteria, or at home. B. cereus spores can germinate during storage, increasing the risk of foodborne disease (Finlay et al., 2000). It is therefore necessary to investigate growth and toxin production by B. cereus in cooked rice for risk assessment studies of B. cereus foodborne disease. In this study, we stimulated the storage of cooked rice to determine risk factors. In a previous study, Finlay et al. (2002) reported the germination, growth, and emetic toxin production by B. cereus in boiled rice with a 24-h sampling interval. However, research is needed to investigate the effects of heat on the growth and toxin production by B. cereus in cooked rice. In addition, we determined the rate of B. cereus toxin production in shorter intervals (minimum 3 h) during the incubation period. Specifically, the objectives of this study were (1) to investigate the growth and toxin production of B. cereus in cooked-rice in order to identify safe storage temperatures for cooked rice and (2) to determine the thermal stability of B. cereus and its toxins in contaminated cooked rice.
Materials and Methods
Preparation of bacterial strains
Mixed cultures of B. cereus strains, JNHE36 (enterotoxin toxin gene nhe and hbl, emetic toxin gene ces) isolated from feces, ATCC 14579 (enterotoxin toxin gene nhe and hbl), and F4810/72 (enterotoxin toxin gene nhe and emetic toxin gene ces) (Kim, 2010; Kim et al., 2010a), were used in this study. Frozen stock cultures maintained at −70°C in tryptic soy broth (TSB; Becton, Dickinson and Company, Sparks, MD) containing 20% glycerol were thawed at room temperature, and 1 mL of each strain was inoculated separately into 100 mL of TSB in a 250-mL Erlenmeyer flask at 35°C with shaking at 150 rpm for 24 h in order to reach the stationary phase. Each vegetative cell suspension was centrifuged at 3000×g for 10 min at 4°C. The supernatant was removed, and the pellet was washed twice using 0.1% (wt/vol) sterile peptone water (PW; Becton, Dickinson and Company) and resuspended in 0.1% sterile PW. The cell suspensions were mixed and stored at 4°C and used for inoculation within 2 days.
Preparation and inoculation of cooked rice
Raw prewashed rice was purchased from a local supermarket in Chuncheon, Korea. An approximately 500-g rice sample was cooked in a pressure cooker with a 1:1.8 (wt/vol) suspension in distilled water. For growth experiments, the 10-g cooked rice samples were inoculated using 0.1 mL of the mixed culture of B. cereus, resulting in a final concentration of about 3 log colony-forming units (CFU)/g. The rice samples were stored at 15–45°C for 72 h. In order to investigate the heat resistance of B. cereus in freshly cooked rice and the cooked rice during storage, cooked rice samples were inoculated using the method described above, and the B. cereus population was determined following 2 days of incubation at 35°C. Freshly inoculated samples, with approximately 8 log CFU/g vegetative cells, were used as control. The stored and freshly inoculated samples were treated at 80, 90, and 100°C for 2 h. At 15-min intervals, the surviving population and the active toxin concentrations (emetic toxin and enterotoxin) were determined.
Microbiological analysis
At each sampling interval, inoculated cooked rice samples (10 g each) were mixed with 90 mL sterile phosphate-buffered saline (PBS, Sigma, Poole, UK) in a stomacher bag and then stomached for 2 min (Lab-blender 400, Seward, London, UK). From appropriate dilutions made in 0.1% (wt/vol) sterile PW, 100 μL of the sample suspensions were spread onto duplicate mannitol–egg yolk–polymyxin agar plates (Becton, Dickinson and Company). The agar plates were incubated at 35°C for 24 h to enumerate B. cereus, and data were expressed as log CFU per gram. Means of cell populations from each treatment were calculated from three replications of each experiment. Each time interval had two plates per independent replicate.
Detection of B. cereus toxin
At each sampling interval, the inoculated cooked rice samples (10 g each) were mixed with 10 mL sterile PBS (Sigma) in a stomacher bag and then homogenized for 2 min (Lab-blender 400). Samples were then centrifuged at 4500×g for 40 min at 4°C, and the supernatants were used for detection of B. cereus diarrheal toxin. For emetic toxin detection, supernatants were autoclaved for 15 min at 121°C to denature B. cereus diarrheal toxin prior to the toxin assay (Kim et al., 2010b).
The B. cereus enterotoxin was assayed using a Tecra® Bacillus diarrheal enterotoxin immunoassay kit (Bioenterprises Ltd., Roseville, New South Wales, Australia). The assay was performed according to the manufacturer's instruction. The toxin detection limit is 1 ng/mL. B. cereus emetic toxin was determined using a Hep-2 cell vacuolation assay as previous reported (Taylor et al., 2005), with minor modifications. One hundred microliters of each supernatant was added to Hep-2 cells in 96-well microtiter plates. The appearance of vacuolation was monitored after incubation for 8 h at 37°C in 5% CO2.
Results and Discussion
Effect of temperature on the growth and toxin production of B. cereus in cooked rice
B. cereus grew during the storage period for all temperatures except 15°C (Fig. 1). The cell population of B. cereus in cooked rice increased from 3.4 log CFU/g to >7.7 log CFU/g within 12 h at 25, 35, and 45°C. During storage at 15°C, B. cereus grew slowly and reached a cell population of 6.2 log CFU/g within 24 h. There were no significant differences (p>0.05) among the final populations of B. cereus for all temperatures tested (15, 25, 35, and 45°C) after a 72-h storage period. Similarly, a previous study reported that B. cereus in boiled rice could reach a final cell population of about 8 log CFU/g after 33-h storage at 30°C and 37°C, but at 22°C 6.30–6.88 log CFU/g was observed (Gilbert et al., 1974). Finlay et al. (2002) investigated growth and toxin production by B. cereus in boiled rice cultures and showed that B. cereus growth could reach viable counts of around 7.2 log CFU/g at 30°C within 24 h, which is lower than the results of our study.
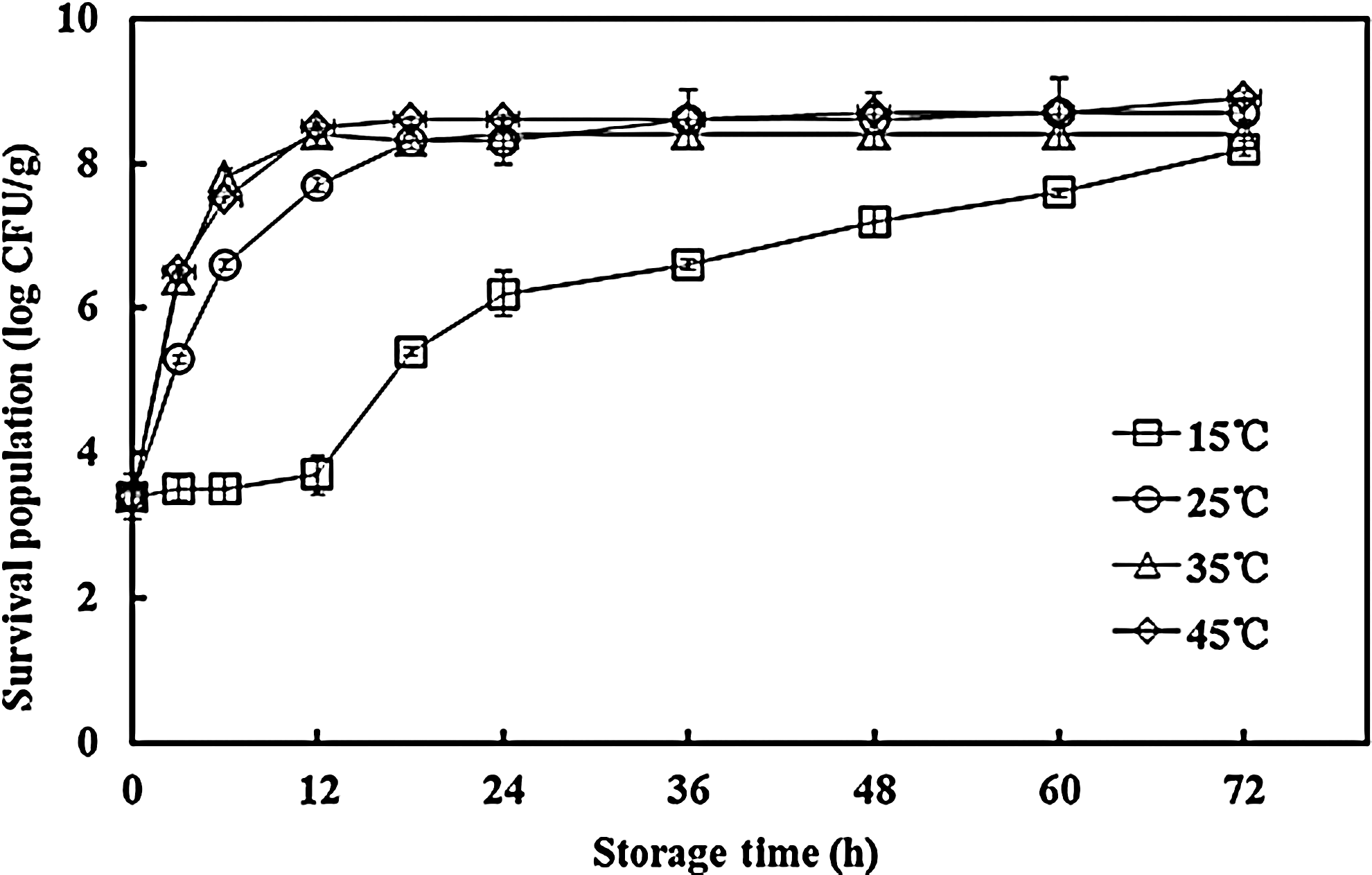
Effect of temperatures on the growth of Bacillus cereus in cooked rice. CFU, colony-forming units.
As shown in Table 1, B. cereus emetic toxin was produced at 15, 25, 35, and 45°C and was first detectable from 18, 12, 6, and 6 h, respectively; the corresponding viable counts of B. cereus were 5.4, 7.7, 7.8, and 7.5 log CFU/g, respectively. The minimum bacterial account of B. cereus needed for emetic toxin production at 15°C was lower than those at 25, 35, and 45°C. Finlay et al. (2002) reported that the times needed for emetic toxin production at 15, 20, and 30°C were 48, 24, and 24 h, with corresponding viable counts of B. cereus of 6.1, 7.2, and 7.2 log CFU/g, respectively. At higher temperatures (>20°C), the results reported by Finlay et al. (2002) were similar to our results, requiring >7 log CFU/g for emetic toxin production. Finlay et al. (2000) reported that B. cereus emetic toxin production was not observed at 15°C unless the viable count reached 6 log CFU/g in skim-milk medium. B. cereus emetic toxin was detected at 12°C with the mean viable counts of 4.40±1.04 log CFU/g, and the toxin titer achieved at 12– to 15°C was higher than 30 to 37°C. A similar result was observed in our study (Fig. 1 and Table 1) (i.e., that emetic toxin could be produced at low temperatures [15°C] with a low cell population: 5.4 log CFU/g in cooked rice). These data may be valuable and helpful for food safety control and improvement of cooked rice products. As shown in Table 1 and Figure 1, B. cereus emetic toxin was formed during the late exponential and/or stationery growth phase at all temperatures, which agreed with other studies (Mikami et al., 1994; Agata et al., 1995; Haggblom et al., 2002). Lücking et al. (2009) reported that ces promoter activity reached a peak at 15 h, which corresponded to the exponential phase of B. cereus growth at 30°C. The results reported by Lücking et al. (2009) could support our findings, since the strain (F4810/72) used in the present study for producing emetic toxin contained the ces gene.
“+” represents toxin production; “−” represents no toxin production.
The results in the present study were different from those of Finlay et al. (2002) concerning the time until toxin production. The probable reason for this apparent discrepancy may be that the bacterial cells were in different physiological states, or there may be strain variation. Another possible reason for the discrepancies was that emetic toxin might be produced at higher temperatures within 24 h. The detection of B. cereus emetic toxin was performed according to the Hep-2 cell vacuolation assay. The toxin from the test strain was observed to cause vacuolation in Hep-2 cells treated with the autoclaved supernatant of the sampling cooked rice samples, compared to no vacuolation with the control.
B. cereus enterotoxin was not detectable at 15°C, but was observed at 25, 35, and 45°C and was first detectable from 48, 18, and 18 h, respectively. This is similar to a previous report that B. cereus enterotoxin was detectable after 48 and 24 h at 21 and 30°C, respectively, and was not detected in cereal incubated less than 12 h at 8, 15, 21, and 30°C (Jaquette and Beuchat, 1998). The viable counts of B. cereus needed to produce enterotoxin were 8.6, 8.3, and 8.6 log CFU/g. The minimum B. cereus cell population needed for production of enterotoxin was >7 log CFU/g at temperatures of 4–37°C, in agreement with previous reports (van Netten et al., 1990; Jaquette and Beuchat, 1998). However, in another report, B. cereus enterotoxin was not detected after cell counts reached 8 log CFU/g at 48 h, 20°C (Ankolekar and Labbe, 2009). In the present study, no detectable enterotoxin was observed at 15°C until 72 h of incubation time, even though the mean viable count reached 8.2 log CFU/g. This is contrary to the report that B. cereus could grow at 8°C in a reconstituted rice-based infant cereal and that enterotoxin production occurred at 15°C after 72 h (Jaquette and Beuchat, 1998). In addition, van Netten et al. (1990) reported that B. cereus in rice meal could produce enterotoxin at 7 and 17°C after 11 days and 2 days when the population reached 6.9 and 7.1 log CFU/g, respectively. One reason for the differences in these studies may be that different B. cereus strains were used (van Netten et al., 1990; Jaquette and Beuchat, 1998).
Heat resistance of B. cereus growth and toxin in cooked rice at different temperatures
In order to investigate the heat resistance of B. cereus in cooked rice, the freshly inoculated and stored samples were treated at 80, 90, and 100°C. The results are shown in Figure 2. In the freshly inoculated rice with B. cereus, cells were killed at all the treated temperatures within 30 min. However, B. cereus in stored rice samples were not destroyed with heat treatment at 80, 90, and 100°C, regardless of the treatment time. The results indicate that thermostable B. cereus spores were produced during the storage period and suggested that cooked rice contaminated with B. cereus might present a potential risk to consumers after storage. In accordance with our findings, a thermal treatment resulted in a 3.47 log reduction of B. cereus in a vegetable substrate after 10-min thermal treatment at 90°C and pH 6.2 (Fernandez et al., 2002). Chung and Sun (1986) reported that a 17.5-min cooking period would yield a 2-log reduction of B. cereus spores isolated from rice, while an 11.5-min cooking period obtained a 1-log reduction. In the present study, an approximate 3-log reduction of B. cereus in the stored cooked rice was observed at all the treated temperatures. The difference between the previous research and our findings might be attributed to the different food matrices used in the studies. Alternatively, results could be biased by spores produced during the storage period, which were difficult to inactivate by heating.

Heat resistance of Bacillus cereus in cooked rice at 80, 90, and 100°C (⋄: freshly inoculated cells without storage at 35°C for 2 days, ◯: cells in cooked rice incubated at 35°C for 2 days). CFU, colony-forming units.
Table 2 shows that B. cereus enterotoxin was easily destroyed by heating for 15 min at all of the treatment temperatures (80, 90, and 100°C), while B. cereus emetic toxin was difficult to inactivate even at 100°C for 2 h. Similar findings showed that emetic toxin could survive 121°C for 90 min and was also resistant to proteolytic degradation and pH extremes (Granum and Lund, 1997). These data indicate that further emetic toxin research is needed.
“+” represents exist toxin; “−” represents no toxin detection.
Conclusions
In conclusion, B. cereus emetic toxin was produced faster than enterotoxin in a rice cooker under the same storage conditions, and emetic toxin was more thermostable than enterotoxin. The results demonstrated that reheating treatment was not helpful to avoid or reduce the risk of disease if cooked rice contaminated by B. cereus has been stored in a warm temperature for a period of time. Therefore, the best way to avoid food poisonings would be to improve the sanitizing conditions and to consume cooked rice as soon after cooking as possible. Results from this study should help enhance the food microbiological safety of cooked rice and related products, and improve the management strategy for these foods, especially in cooked rice contaminated with B. cereus.
Footnotes
Disclosure Statement
This research was in part supported by Bio-industry Technology Development Program (LBI605), Ministry of Agriculture, Food and Rural Affairs.