
Abstract
Nontyphoidal Salmonella infections caused by antimicrobial-resistant (AMR) strains are of great public health concern. We compared the phenotypic and genotypic relationships among temporally and spatially related AMR Salmonella isolates (n=1058) representing several predominant serovars, including Salmonella Typhimurium, Salmonella Typhimurium var. 5-, Salmonella Derby, Salmonella Heidelberg, Salmonella Muenchen, Salmonella Schwarzengrund, and Salmonella Rissen of human clinical cases (n=572), pig (n=212), and farm environment (n=274) origin in North Carolina. Antimicrobial susceptibility testing was performed using the broth microdilution method, and genotypic resistance determinants, including class I and II integrons, were identified. Overall, Salmonella isolates exhibited the highest frequency of resistance to tetracycline (50%), followed by sulfisoxazole (36%) and streptomycin (27%). We identified 16 different antimicrobial resistance genes, including extended spectrum and AmpC β-lactamases-producing genes (bla TEM, bla PSE, and bla CMY-2), in all the β-lactam- and cephalosporin-resistant Salmonella isolates from humans, pigs, and the environment. Class I integrons of 1-kb and 1.2-kb size were identified from all the three sources (humans, 66%; pigs, 85%; environment, 58%), while Class II integrons of 2-kb size were identified only in pig (10%) and environmental (19%) isolates. We detected genotypic similarity between Salmonella Typhimurium isolated from humans, pigs, and the environment while serovars Derby, Heidelberg, and Muenchen exhibited genotypic diversity. Detection of AMR Salmonella isolates from humans, pigs, and the environment is a concern for clinicians and veterinarians to mitigate the dissemination of AMR Salmonella strains.
Introduction
A
North Carolina ranks second in pork production in the United States, contributing 14.4% of the national inventory (USDA, 2009). Several Salmonella outbreaks associated with pork in humans have been reported in the United States and worldwide in the past decade (CDC, 2007; Wojcik et al., 2012; Scavia et al., 2013). To the authors' knowledge, no studies have been conducted to compare temporally and spatially related NTS isolates from human clinical cases with Salmonella isolates arising from pigs and the environment as part of a longitudinal study in the United States. Therefore, the objective of this study was to characterize and compare Salmonella isolates from humans, pigs, and the environment by antimicrobial susceptibility testing, identification of AMR genes, and strain genotyping to determine whether temporally and spatially related Salmonella isolates from multiple sources in North Carolina were phenotypically and genotypically similar or diverse.
Materials and Methods
Salmonella isolate sources
In the present study, a total of 1058 temporally and spatially related NTS isolates from human clinical cases (n=572), pigs (n=212) and the farm environment (n=274) originating from multiple counties in North Carolina were characterized at the phenotypic and genotypic levels. The human NTS strains were clinical isolates received from the North Carolina State Public Health Laboratory. All samples were collected from October 2008 to December 2011. The geographical distribution of NTS isolates is represented in Table 1. The human clinical isolates (n=572) originated from 72 counties in North Carolina. The majority of isolates (n=183; 32%) came from Mecklenburg, Wake, Cumberland, and Cabarrus counties. Of 572 human clinical isolates, 173 (30%) originated from the primary pig-producing counties, including Duplin, Johnston, Sampson, and Cumberland counties. All of the pig (n=212) and the environmental (n=274) isolates originated from the major pig-producing counties mentioned above (Table 1). The Salmonella isolates from pigs and their environment were collected as part of longitudinal study conducted from October 2008 to December 2011 on 30 conventional farms at different stages of production from farm to slaughter in North Carolina. The environmental sampling from conventional swine farms consisted of water, feed, soil, and barn floor swabs. In addition, lagoon (repository of waste water draining from the barns), barn floor, and interfarm truck floor swab samples were also collected at conventional farms. The details of the study design, sampling methods, estimates of Salmonella prevalence in pigs and their environment, antimicrobial susceptibility profiles, serovars distribution, and their phenotypic and genotypic characterizations have been reported elsewhere (Keelara et al., 2013).
N=total no. of serovars.
Other represents any county with less than three isolates.
H, human, P, pig; E, environment; N/A, not applicable.
Antimicrobial susceptibility testing (AST)
AST was performed for all Salmonella isolates against a panel of 15 antimicrobials via the broth microdilution method in a 96-well Sensititre™ plate (CMV1AGNF, the “NARMS panel”; Trek Diagnostics, Inc., Cleveland, OH). The classes of antimicrobials tested included the following: aminoglycosides (amikacin, AMI; gentamicin, GEN; kanamycin, KAN; streptomycin, STR), β-lactam/β-lactamase inhibitor combinations (amoxicillin/clavulanic acid, AUG), penicillins (ampicillin, AMP), cephems (ceftriaxone, AXO; cefoxitin, FOX; ceftiofur, TIO), quinolones (ciprofloxacin, CIP; nalidixic acid, NAL), folate pathway inhibitors (sulfisoxazole, FIS; trimethoprim/sulfamethoxazole, SXT), phenicols (chloramphenicol, CHL), and tetracyclines (tetracycline, TET). The testing procedure was carried out as described in the previous study (Keelara et al., 2013). The minimum inhibitory concentrations (MICs) were recorded and breakpoints were determined based on Clinical and Laboratory Standards Institute (CLSI) recommendations (CLSI, 2010). The isolates exhibiting resistance to three or more classes of antimicrobials were classified as MDR.
Detection of resistance genes and integrons
All the AMR Salmonella isolates from humans, pigs, and the environment were screened for the presence of all known corresponding resistance genes and class 1 and 2 integrons. Polymerase chain reaction (PCR) was performed for detection of different AMR genes using the following primers, including: the extended spectrum β-lactamases bla TEM, bla PSE-1 (Carlson et al., 1999) and bla CMY-2 genes (Zhao et al., 2001), streptomycin resistance coding aadA1/A2 and strA/B genes (Madsen et al., 2000), kanamycin resistance coding aphAI, Kn genes (Frana et al., 2001), chloramphenicol resistance coding cml gene, sulfisoxazole and trimethoprim/sulfamethoxazole resistance coding sulI (Briggs et al., 1999) and sulII genes (Aarestrup et al., 2003), tetracycline resistance coding tet(A), tet(B), tet(C), and tet(G) genes, and class1 and class 2 integrons (Ng et al., 1999). Template DNA was purified using the DNeasy blood and tissue kit (Qiagen, Valencia, CA) according to the manufacturer's recommendations. Amplification reactions were carried out as described in the above studies.
Pulsed-field gel electrophoresis (PFGE) analysis
A subset of Salmonella isolates (pigs, n=46; environment, n=80; humans, n=271) that were purposively chosen to be representative of different sources of origin, serovar, counties, and AMR profiles were genotyped using PFGE, following the PulseNet protocol (Ribot et al., 2006). BioNumerics software version 6.1 (Applied Maths, Kortrijk, Belgium) was used to analyze the PFGE images. A dendrogram was generated to determine the clonal relationship among human, pig, and environmental isolates using an unweighted-pair group method with average linkages (UPGMA), and with band position tolerance and optimization of 1.5% each.
Statistical analysis
We used odds ratios (OR) with 95% confidence intervals (CI) to explore the associations between MDR and the predominant serovars detected in our study (SigmaPlot 11.2; Systat Software, Inc., Chicago, IL). A p-value of 0.05 or lower was considered statistically significant.
Results
Distribution of Salmonella serovars
The predominant serovars identified among human clinical Salmonella isolates (n=572) were Salmonella Typhimurium (50.6%), followed by Salmonella Typhimurium var. 5- (23.6%), Salmonella Muenchen (13.8%), and Salmonella Heidelberg (8.7%). Serovars from pig (n=212) and environmental (n=274) origin were predominantly represented by Salmonella Typhimurium (pigs, 59.4%; environment, 67.1%) and Salmonella Derby (pigs, 34.9%; environment, 11.6%).
Antimicrobial resistance profiles
A total of 1058 Salmonella isolates from humans (n=572), pigs (n=212), and their environment (n=274) were tested against a panel of 15 antimicrobials. The overall MIC levels and frequency of AMR of Salmonella isolates are presented in Table 2 and Figure 1. All of the Salmonella isolates, irrespective of source of origin, exhibited similar MIC50 and MIC90 with the exceptions being the following: TET MIC50 in humans, which was 4 μg/mL (versus pigs and environment: 32 μg/mL); FIS MIC50 in pigs, which was 256 μg/mL (humans and environment, 64 μg/mL); and CHL MIC90 in environmental isolates, which was 16 μg/mL (pigs and humans, 32 μg/mL). Overall, Salmonella isolates exhibited the highest frequency of resistance to tetracycline (TET; 50%), followed by sulfisoxazole (FIS; 36%) and streptomycin (STR; 27%). In addition, Salmonella isolates from human clinical cases exhibited slightly higher frequencies of resistance to cephalosporins including AXO, FOX, and TIO (3% each) compared to pigs and environmental isolates (2% each). Overall, the frequency of AMR was higher in Salmonella isolates of pig origin (82.6%) followed by environmental (67.6%) and human clinical isolates (28%), with the exception of AMI (0.7%) and CIP (0.3%) resistance, which were only exhibited by human clinical isolates.
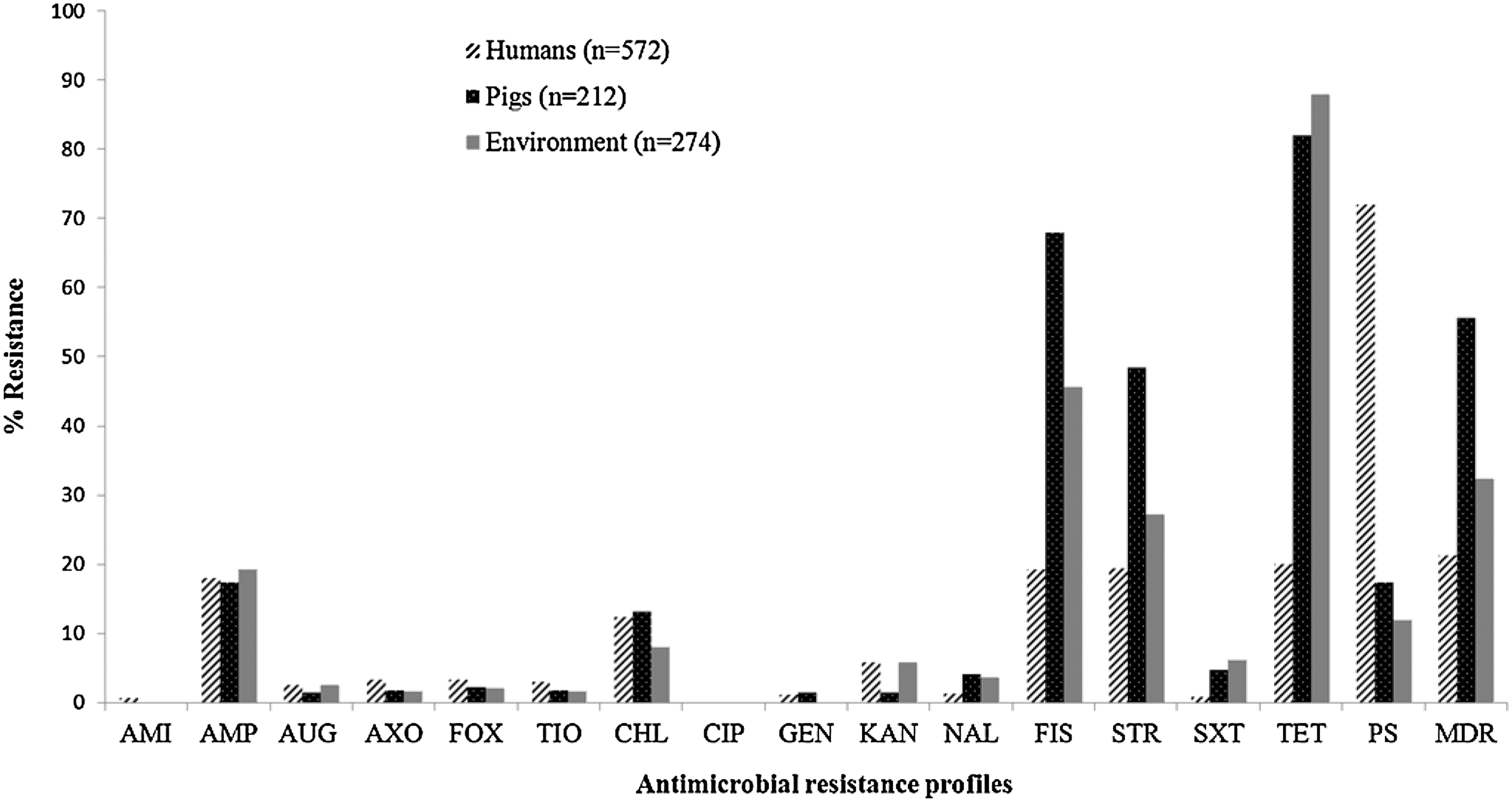
Antimicrobial resistance profile of Salmonella isolates from humans, pigs, and environment. Antimicrobials: AMI, amikacin; AMP, ampicillin, AUG, amoxicillin/clavulanic acid; AXO, ceftriaxone; CHL, chloramphenicol; CIP, ciprofloxacin; FIS, sulfisoxazole; FOX, cefoxitin; GEN, gentamicin; KAN, kanamycin; STR, streptomycin; SXT, trimethoprim/sulfamethaxazole; TET, tetracycline; TIO, ceftiofur. MDR, multidrug resistance (resistance ≥3 classes of antimicrobials); PS (pansusceptible): susceptible to all the antimicrobials tested.
Antimicrobials: AMI, amikacin; AMP, ampicillin; AUG, amoxicillin/clavulanic acid; AXO, ceftriaxone; CHL, chloramphenicol; CIP, ciprofloxacin; FIS, sulfisoxazole; FOX, cefoxitin; GEN, gentamicin; KAN, kanamycin; NAL, nalidixic acid; STR, streptomycin; SXT, trimethoprim/sulfamethoxazole; TET, tetracycline; TIO, ceftiofur.
% R, percent resistance.
MDR patterns identified
The major MDR patterns identified in humans, pigs and their environment are represented in Table 3. The highest frequency of MDR patterns was detected in Salmonella isolated from pigs (50.9%) compared to the environment (22.6%) and human (19.2%). The most common MDR patterns identified in all the three sources were FIS-STR-TET, and AMP-CHL-FIS-STR-TET associated with Salmonella Derby and Salmonella Typhimurium, respectively. The Salmonella Derby serovar was significantly (p<0.001) associated with an MDR pattern FIS-STR-TETwith an OR of 27 (95% CI=15.6–46.8) compared to Salmonella Typhimurium isolates with this MDR pattern. Specific MDR patterns were detected in particular hosts including FIS-STR-TET and AMP-AUG-AXO-FOX-TIO-TET in Salmonella Typhimurium isolated from pigs and environmental isolates. In contrast, the AMP-KAN-STR-TET (n=5) pattern was found only in Salmonella Heidelberg isolates of human origin (Table 3). A single human clinical Salmonella Typhimurium isolate was resistant to 12 antimicrobials (including quinolones and multiple cephalosporins) out of 15 tested with a MDR pattern of AMI-AXO-CHL-CIP-FOX-FIS-GEN-KAN-NAL-STR-TIO-TET.
Antimicrobials: AMP, ampicillin; AUG, amoxicillin/clavulanic acid; AXO, ceftriaxone; CHL, chloramphenicol; FIS, sulfisoxazole; FOX, cefoxitin; KAN, kanamycin; STR, streptomycin; TET, tetracycline; TIO, ceftiofur.
bla TEM /bla PSE, genes encoding ampicillin; cmlA, chloramphenicol; aadA1/A2 and str A/B, streptomycin; tet (A) (C), tetracycline; bla CMY-2, extended-spectrum β-lactamases.
H, MDR Salmonella isolates representing humans; P, MDR Salmonella isolates representing pigs; E, MDR Salmonella isolates representing environment.
Molecular characterization of AMR determinants
Using PCR, we identified 16 different AMR genes conferring resistance to a number of classes of antimicrobials (Table 3). Ampicillin resistance was predominantly encoded by the extended-spectrum β-lactamase–producing bla TEM gene (humans, 71%; pigs, 49%; environment, 81%), followed by the bla PSE-1 gene (humans, 17%; pigs, 49%; environment, 11%). All the Salmonella isolates from humans, pigs, and the environment that were resistant to third-generation cephalosporins carried the AmpC β-lactamases–producing bla CMY-2 gene. Tetracycline-resistant isolates from pigs and the environment harbored four different genes, predominantly tet(A) (pigs, 72%; environment, 68%) followed by tet(B) (pigs, 10%; environment, 19%), tet(C) (pigs, 4%; environment, 5%), and tet(G) (pigs, 15%; environment, 9%) genes. Among Salmonella isolates from humans, tetracycline resistance was predominantly encoded by tet(G) (58%), tet(A) and tet(C) (each at 17%), and with the notable absence of the tet(B) gene. The Salmonella Typhimurium penta-resistant (AMP-CHL-FIS-STR-TET) isolates from humans, pigs, and the environment harbored similar resistant genes, including bla TEM/bla PSE, cmlA, aadA1/A2 and tet(G), encoding resistance to AMP-CHL-STR-TET, respectively (Table 3). The FIS encoding gene sul2 was only detected in the isolates with FIS-STR-TET resistance pattern (Table 3). We detected class I integrons of 1 kb and 1- and 1.2-kb size (humans, 66%; pigs, 85%; environment, 58%) and class II integrons of 2 kb (humans, 0%; pigs, 10%; environment, 19%) in MDR Salmonella isolates from pigs and the environment. All the penta-resistant Salmonella Typhimurium from each source exhibited class I integrons of 1- and 1.2-kb size.
Pulsed-field gel electrophoresis (PFGE)
Genotyping of temporally and spatially related Salmonella isolates (n=397) by PFGE with the XbaI restriction enzyme distributed them into 74 major clusters consisting of isolates with similar fingerprint profiles, and another 118 unique PFGE patterns represented by a single isolate each (data not shown). Salmonella Muenchen isolates from each source had more diversified fingerprint profiles compared to other study serovars. All the Salmonella Heidelberg isolates were closely related based on fingerprint profiles and distributed into one large cluster with 80% genotypic similarity.
We created individual dendrograms, one each for Salmonella Derby and Salmonella Typhimurium, to represent relationship between counties and AMR patterns. Identical fingerprint profiles were found among Salmonella Derby isolates of pig and environmental origin from Cumberland, Johnston, and Sampson counties (cluster 3–5; Fig. 2). All the Salmonella Derby isolates with MDR pattern FIS-STR-TET from the three sources were clustered closely in the dendrogram (cluster 2–6; Fig. 2). Even though the Salmonella Derby from humans (ID HS749 and HS289) had an identical resistance pattern (FIS-STR-TET) to those that originated from pigs and the environment in the same county (Sampson and Cumberland), they had different fingerprint profiles (Fig. 2). Genotypic (100%) and phenotypic similarity based on MDR pattern (AMP-CHL-FIS-STR-TET) among isolates of human, pig, and environmental origin was detected only among the Salmonella Typhimurium serovar (cluster 1 and 2; Fig. 3). Overall, the isolates from human, pigs, and the environment exhibited higher levels of genotypic diversity among Salmonella Derby, Salmonella Heidelberg, and Salmonella Muenchen isolates and much more genotypic similarity in Salmonella Typhimurium isolates.

Dendrogram showing genotypic similarity among Salmonella Derby isolated from humans, pigs, and environment. PFGE, pulsed-field gel electrophoresis; AR, antimicrobial resistance.

Dendrogram showing genotypic similarity among Salmonella Typhimurium isolated from humans, pigs, and environment. PFGE, pulsed-field gel electrophoresis; AR, antimicrobial resistance.
Discussion
The objective of this study was to compare temporally and spatially related NTS isolates from humans, pigs, and the environment based on their AMR phenotypes and genotypic fingerprint profiles. The frequency of AMR was higher in Salmonella isolates from pigs (82.6%) and their environment (67.6%) compared to human clinical Salmonella isolates (28%). In addition, we observed a higher MIC50 (32 μg/mL) for TET in pig and environmental isolates compared to the MIC50 (4 μg/mL) for human isolates. One reason could be that the pig and environmental isolates were from conventional production systems that routinely use antimicrobials for prophylaxis (tetracycline and macrolides as growth promoter) and therapeutic purposes, thereby contributing to an increased frequency of resistance and MIC to antimicrobials (Keelara et al., 2013).
Resistance to β-lactams, including ampicillin and cephalosporins, was observed at low levels in human, pig, and environmental isolates.
Human clinical Salmonella isolates exhibited resistance to CIP (0.3%), which was never observed in pig or environmental isolates. AMR trend to CIP in Salmonella isolates of humans appears to be increasing every year based on the National Antimicrobial Resistance Monitoring System (NARMS) annual report on AMR of NTS isolates from 1999 to 2010, as well as other studies (Patchanee et al., 2008; Dorr et al., 2009; NARMS, 2010; Boxstale et al., 2012). The use of ciprofloxacin for treating severe bacterial infections could potentially explain the increase in AMR to this important antimicrobial. AMR among Salmonella to ciprofloxacin and third-generation cephalosporins is concerning since they are the drugs of choice for treating human invasive Salmonella infections.
A higher frequency of MDR isolates was observed in Salmonella isolates of pigs (50.9%) and their environment (22.6%) compared to humans (19.2%). According to the NARMS executive report (1999–2010), the number of reported NTS MDR isolates in humans dropped to 9% from 2007 to 2010. Interestingly, in our study the frequency of MDR isolates from human clinical NTS isolates was higher (19.2%) compared to recent NARMS data (NARMS, 2010). This variation could be attributed to regional differences in the definition of MDR used, reporting practices, human antimicrobial use, or regional strain variation. The two most common patterns observed in this study among all the three sources were FIS-STR-TET and AMP-CHL-FIS-STR-TET. The Salmonella Derby serovar was 27 times (odds ratio) more likely to be associated with the FIS-STR-TET pattern than Salmonella Typhimurium, which suggests establishment of this serovar with a specific MDR pattern in this region. Emergence of Salmonella Derby with this specific MDR pattern is concerning, as it is the top serovar isolated from pigs (NARMS, 2010). Importantly, all of the three classes of antimicrobials to which it is resistant are currently considered critically important by the World Health Organization (WHO, 2012).
The MDR pattern AMP-CHL-FIS-STR-TET was detected at a higher frequency among the human Salmonella Typhimurium isolates when compared to pigs and their environment. This penta-resistant pattern is specific to Salmonella Typhimurium phage type DT104 and is commonly associated with clinical salmonellosis and human foodborne outbreaks worldwide (Helms et al., 2005; Gebreyes et al., 2009; Medalla et al., 2013). This penta-resistant phenotype is often a component of higher-order MDR patterns that include resistance elements against the critically important fluoroquinolones and third- and fourth-generation cephalosporins. As an example, a single Salmonella Typhimurium isolate from a clinical human case exhibited resistance to 12 antimicrobials: very importantly, including resistance against both quinolones and multiple cephalosporins. This pattern in Salmonella Typhimurium has been previously reported from human clinical cases (Patchanee et al., 2008; Glenn et al., 2013).
The molecular characterization of these isolates using PCR to detect the presence of genes encoding resistance to various antimicrobials was consistent among the Salmonella isolates from all three sources. Detection of the AmpC β-lactamases producing gene bla CMY-2 among all the Salmonella isolates resistant to third-generation cephalosporins, including AXO and TIO, was in agreement with previous reports (Zhao et al., 2001; Van et al., 2012). This is the first report of AmpC β-lactamases producing (bla CMY-2) Salmonella from this region. Presence of this gene is concerning because Salmonella isolates carrying these genes are at an increased risk of acquiring resistance to other classes of antimicrobials (Hamilton et al., 2012; Glenn et al., 2013).
The majority of penta-resistance in Salmonella Typhimurium was encoded by resistance genes carried on class I integrins, and these are believed to play an important role in the dissemination of AMR among susceptible populations of Salmonella both in humans and animals (Aarestrup et al., 2008). This was evidenced by detection of class I integrons of 1 and 1.2 kb in all the penta-resistant (AMP-CHL-FIS-STR-TET) Salmonella Typhimurium isolates of human, pig, and the environment. All of these isolates carried similar resistance genes, including bla TEM/bla PSE, cmlA, aadA1/A2, and tet(G). All of the Salmonella Derby with FIS-STR-TET pattern carried a Class I integron of 1-kb size, suggesting too that this serovar could play a potentially important role in transmission of AMR to a susceptible population as for Salmonella Typhimurium. We identified the rarely reported Class II integrons of 2-kb size in pig and environmental Salmonella isolates. To the best of our knowledge, this is the first report of Class II integrons in Salmonella in this region. Further analysis of the class II integrons detected in our study will be performed at a later date to identify additional resistant gene cassettes that may be located on them.
A subset of Salmonella isolates from humans, pigs, and the environment exhibiting similar AMR patterns and originating from pig-producing and non-pig-producing counties were genotyped using PFGE. We identified 74 major clusters consisting of isolates with similar fingerprint profiles and an additional 118 unique profiles. Isolates of Salmonella Muenchen from human clinical cases that originated from the same county were grouped into two different clusters. However, the majority of Salmonella Muenchen isolates from humans had a unique fingerprint profile, which has previously been reported in Salmonella Muenchen and other Salmonella serovars (Gebreyes et al., 2005; Patchanee et al., 2008). This diversity indicates that there are numerous clones circulating for serovar Muenchen in this region. Similarly, Salmonella Derby isolates from humans were more diverse compared to pig and environmental isolates, even though they had similar resistance patterns and were isolated from counties adjacent to major pig-producing counties; this indicates that different genotypes of Salmonella Derby are prevalent in North Carolina. We identified 100% genotypic similarity only in penta-resistant Salmonella Typhimurium isolates from humans, pigs, and the environment originating from the major pig-producing counties of North Carolina (cluster 1 and 2; Fig. 3). It is worth mentioning that the human Salmonella Typhimurium isolates in these clusters were reported from counties (Cumberland and Wake) adjacent to the major pig-producing counties of North Carolina. However, it is important to mention that no human Salmonella outbreak associated with pork has been reported from this area.
Conclusions
In conclusion, this is the first report to compare temporally and spatially related NTS isolated from humans, pigs, and the environment from this region of the United States. Detection of similar MDR patterns, Class I and II integrons in Salmonella of pigs, their environment, and of human origin is a growing public health concern. Even though the human isolates were not directly linked to pigs/farm environment, identical fingerprint profiles suggest that the same strains are circulating in North Carolina. However, it is important to mention that this study cannot predict the direction of Salmonella transmission. Further studies to determine the role played by different reservoirs in determining the occurrence and dissemination of AMR Salmonella in the food chain will be key to identifying these sources.
Footnotes
Acknowledgments
We thank the swine producers and swine-processing facilities in North Carolina for allowing access to their facilities and the North Carolina State Laboratory of Public Health for providing human clinical Salmonella strains. This work was funded by the United States Department of Agriculture (USDA) National Research Initiative grant (2008-529245), USDA National Integrated Food Safety Initiative grant (2008-529461), and National Pork Board grant (555105).
Disclosure Statement
No competing financial interest exists.