
Abstract
Arcobacter species have a widespread distribution with a broad range of animal hosts and environmental reservoirs, and are increasingly associated with human illness. To elucidate the routes of infection, several characterization methods such as pulsed-field gel electrophoresis (PFGE), amplified fragment-length polymorphism, and enterobacterial repetitive intergenic consensus (ERIC)-PCR have already been applied, but without proper validation or comparison. At present, no criterion standard typing method or strategy has been proposed. Therefore, after the validation of PFGE, those commonly applied typing methods were compared for the characterization of six human- and animal-associated Arcobacter species. With a limited number of isolates to be characterized, PFGE with restriction by KpnI is proposed as the first method of choice. However, ERIC-PCR represents a more convenient genomic fingerprinting technique when a large number of isolates is involved. Therefore, a first clustering of similar patterns obtained after ERIC-PCR, with a subsequent typing of some representatives per ERIC cluster by PFGE, is recommended. As multiple genotypes are commonly isolated from the same host and food, genomic plasticity has been suggested. The in vitro genomic stability of Arcobacter butzleri and A. cryaerophilus was assessed under two temperatures and two oxygen concentrations. Variability in the genomic profile of A. cryaerophilus was observed after different passages for different strains at 37°C under microaerobic conditions. The bias due to these genomic changes must be taken into account in the evaluation of the relationship of strains.
Introduction
S
One of the main challenges in Arcobacter research is to identify the different transmission routes. Various studies reported multiple species and moreover several strains present in the same host and on food products (Houf et al., 2002; Van Driessche et al., 2004; Van Driessche et al., 2005), hampering correct identification of vectors and reservoirs. Currently, genotypic methods are commonly applied, but not all have been validated (e.g., pulsed-field gel electrophoresis [PFGE]) or used (e.g., multilocus sequence typing) for all Arcobacter species (Hume et al., 2001; Son et al., 2006; Miller et al., 2009; Giacometti et al., 2013). At present, no criterion standard typing method or strategy has been proposed. Furthermore, strain or genotype delineation is currently arbitrary without guidelines specified. Besides the interpretation challenges due to different typing methods, the possibility of genomic rearrangements, as also described in C. jejuni (Wassenaar et al., 1998; Ridley et al., 2008; Leblanc-Maridor et al., 2011), may have their basis in the large strain diversity. However, whether or not these rearrangements are induced by in vitro manipulation is currently unknown.
The aims of this study were to validate PFGE for its use with six human- and animal-associated Arcobacter species, and to propose a typing strategy for arcobacters based on the comparison of PFGE with two other previously validated and commonly used typing methods: amplified fragment gel polymorphism (AFLP) and enterobacterial repetitive intergenic consensus (ERIC)-PCR. Finally, the in vitro genomic stability of A. butzleri and A. cryaerophilus was evaluated by multiple cultivations under different temperature and atmosphere conditions.
Materials and Methods
Bacterial strains and growth conditions
For the validation and comparison of the typing methods, 46 A. butzleri, 33 A. cryaerophilus, 33 A. skirrowii, 38 A. thereius, 11 A. cibarious, and 12 A. trophiarum isolates as well as 25 Arcobacter collection strains (Table 1) were included. The Arcobacter isolates were obtained from food, human stool, and feces from food-producing animals by the Department of Veterinary Public Health, Ghent University, Belgium, using Arcobacter selective isolation methods for food and feces (Houf et al., 2001; Van Driessche et al., 2003; Houf and Stephan, 2007). For the in vitro genomic stability assays, 24 arcobacters were included: 6 A. butzleri and 7 A. cryaerophilus strains recently isolated from pig feces (De Smet et al., 2011), 5 A. butzleri and 4 A. cryaerophilus isolates recently isolated from human stool (Saint-Lucas Hospital, Ghent), as well as 2 A. cryaerophilus collection strains LMG 9904T and LMG 9947.
LMG, Culture Collection of the Laboratory for Microbiology, Ghent University, Belgium; T, type strain; ATCC, American Type Culture Collection.
All strains and isolates were grown on Mueller–Hinton agar (CM 0337, Oxoid, Basingstoke, UK) supplemented with 5% defibrinated horse blood (E&O Laboratories Ltd., Bonnybridge, Scotland) and incubated for 48 h at 30°C under microaerobic conditions by evacuating 80% of the normal atmosphere and introducing a gas mixture of 8% CO2, 8% H2, and 84% N2 into the jar.
Validation of PFGE
Different protocols, based on the PulseNet protocol for Campylobacter jejuni, were tested (PulseNet 2009:
Subsequently, 173 Arcobacter isolates, comprising the six human- and animal-related species and identified by an Arcobacter species-specific multiplex-PCR (Douidah et al., 2010) were characterized with the validated PFGE protocol, using the most discriminatory restriction enzymes.
Comparison of different typing techniques
Preparation of DNA templates
The 25 collection strains and 173 Arcobacter isolates used in the validation of PFGE were cultivated as described above. A cell suspension of each strain with an optical density of 0.074±0.002 (measured at 660 nm [Biophotometer, Eppendorf]), corresponding to approximately 107 colony-forming units/mL was prepared in 10 mL of PBS. Template DNA of each strain was extracted from a 0.5-mL cell suspension by the guanidinium thiocyanate method (Pitcher et al., 1989). Five microliters of each DNA template was size separated by electrophoresis to evaluate their integrity. The concentration as well as the purity of each DNA template was determined spectrophotometrically. The DNA templates were adjusted to 50 ng/μL and stored at −20°C until further analysis.
AFLP
For AFLP analysis, the validated protocol for Arcobacter species by Debruyne et al. (2010) was applied, using HindIII and HhaI as restriction enzymes (New England Biolabs Inc.) and analysis performed by capillary gel electrophoresis using an automated 16-capillary sequencer (3130xl Genetic Analyzer, Applied Biosystems, Carlsbad, CA).
ERIC-PCR
ERIC-PCR assay was exactly performed according to the previously validated protocol for Arcobacter by Houf et al. ( 2002).
Assessment of the in vitro genomic stability of A. butzleri and A. cryaerophilus
In a first experiment, the A. cryaerophilus strains LMG 9947 and LMG 9904T, and four isolates—A. butzleri R643, A. cryaerophilus R637 and R671 from pig feces, and A. cryaerophilus R206 from human stool—were included. Strains were cultivated on Mueller–Hinton agar supplemented with 5% defibrinated horse blood at 30°C under microaerobic conditions for 48 h. One colony was picked and recultivated on Mueller–Hinton agar plates under the same conditions. To avoid subjective selection of specific colonies, further subcultivations were performed by picking a loopful of bacterial growth, striking them on blood plates, and incubating them at 30°C and 37°C, each under aerobic and microaerobic conditions. The strains were further subcultivated in the same way twice a week for 8 weeks (=16 passages). At the start and on even-numbered passages, PFGE was performed as described above. A second experiment was performed based on the results obtained in the first experiment but with an at-random selection of 10 A. butzleri and 8 A. cryaerophilus strains isolated from humans and pigs, as well as again strain R671. The strains were now cultivated for three generations at 30°C and 37°C under microaerobic conditions as described above and subsequently characterized by PFGE.
Data analysis
The PFGE and ERIC profiles were analyzed with GelCompar 3.0 (Applied Maths, Sint-Martems-Latem, Belgium). For PFGE, the Salmonella serovar Braenderup H9812 strain was restricted with XbaI enzyme and used as a molecular size standard. Computer-based normalization and interpolation of the ERIC-PCR profiles was performed using TrackIt™ 100-bp DNA Ladder (Cat. No. 10488-058, Invitrogen). AFLP profiles were collected with the Data Collection software v 3.0 (Applied Maths). Profiles were imported in BioNumerics v 4.61 (Applied Maths) for further analysis using the CrvConv filter. Similarities between fingerprints were calculated using the Dice coefficient for PFGE, and Pearson correlation coefficient for ERIC-PCR. AFLP normalization was performed using the internal LIZ-600 standard, after which similarities between profiles were determined by the Pearson and product moment correlation coefficient. PFGE, ERIC, and AFLP dendrograms were constructed by numerical analysis of obtained profiles using the unweighted-pair group method using arithmetic means algorithm. Profiles were automatically assigned to groups according to (restriction) pattern similarities, which were visually controlled and edited if needed. The relationship between Arcobacter strains was expressed as a percentage of similarity.
Unrelated strains were included to compare the discriminatory ability of the three typing methods using the Simpson's index of diversity (Hunter and Gaston, 1988; Simpson, 1949). This index indicates the probability of two strains sampled randomly from a population belonging to two different types. Furthermore, the Adjusted Rand (AR) and the adjusted Wallace (AW) were calculated in order to compare the congruence between typing methods (Severiano et al., 2011). Online tools were used for calculation of these indexes (
Results
Validation of PFGE
Besides the two restriction enzymes EagI and KpnI previously used in Arcobacter typing (Hume et al., 2001; Son et al., 2006), only the restriction enzymes NruI and StuI generated the required minimum of 10 and a maximum of 30 fragments after in silico restriction of the A. butzleri genome ATCC 49616 (van Belkum et al., 2007). In the characterization of the 25 Arcobacter collection strains, fingerprints for all strains were obtained with all enzymes, but the enzymes EagI and StuI were shown to be less discriminative, since for some strains low numbers of fragments (<5) were obtained (data not shown). Furthermore, they generated identical fingerprints for some strains of different Arcobacter species (data not shown). For further experiments, the restriction enzyme KpnI was selected as first choice as it generated an average number of 15.5 fragments (ranging from 10 to 30), in contrast to an average of 8 bands (ranging from 5 to 13) with the NruI (data not shown). Optimal analyses were achieved with PFGE carried out in a 1% SeaKem (wt/vol) gold agarose (Cambrex) in a PFGE apparatus (Bio-Rad, Hercules, CA) for 18 h with a pulse time of 6.676–13.68 s for KpnI (Son et al., 2006), and 5.2 s–42.3 s for NruI under a constant temperature of 14°C, a gradient of 6 V/cm, an angle of 120°, and a range of 30 kbp–400 kbp. A 100% repeatability and reproducibility of the PFGE protocol, both with KpnI and NruI, was obtained.
Comparison of different typing techniques
Similarities between fingerprints were calculated as described above, with an optimization of 1.0% and a position tolerance of 1.0% for PFGE. For ERIC-PCR, an optimization of 0.5% and 0.5% position tolerance was applied. Due to the high resolution of capillary migration, an optimization and a position tolerance of 0.01% was used for AFLP (Debruyne et al., 2010).
All 198 Arcobacter collection strains and isolates could be characterized by ERIC-PCR and AFLP. With PFGE, no fragment patterns were obtained for one A. cryaerophilus, one A. skirrowii, and nine A. thereius isolates using restriction with both KpnI and NruI. The inability to type all Arcobacter strains resulted in a 94.5% typeability for PFGE, to 100% for ERIC-PCR and AFLP. The discriminatory powers of the three typing methods were determined by calculating the Simpson's index of the unrelated A. butzleri and A. cryaerophilus isolates typed by these methods. Due to the large diversity of Arcobacter isolates, all three methods showed high discriminatory power, with a discriminatory index of 1. AR and AW coefficients were also calculated to explore the correlation between the typing methods; however, the same result of 1 was obtained. Weighing the results of the three typing methods for the unrelated Arcobacter isolates showed comparatively high discriminatory power, revealing no difference between the methods.
In general, ERIC-PCR generated the lowest number of bands with an average between 12 and 15 bands per profile for the six Arcobacter species, except for A. thereius and A. trophiarum, which generated in AFLP the lowest number of bands (i.e., six and nine bands per AFLP profile, respectively). The number of bands generated by PFGE and AFLP was quite similar, except for A. thereius and A. trophiarum, as explained above. Cluster analysis per method of the DNA patterns showed no clear distinction at Arcobacter species level. A large heterogeneity between the isolates was observed, with related or identical strains clustering more closely with all three methods than nonrelated strains. However, no clear relationship between sources of origin or between isolates from animal and human origin was observed. Isolates with similar profiles generated by one method were compared with the corresponding profiles obtained by the other two typing methods. For A. butzleri, two isolates from the Italian outbreak (Vandamme et al., 1992a) (LMG 10900 and 11120) with an identical PFGE profile also generated high similar (>97.57%) ERIC-PCR and AFLP profiles (Fig. 1A). Furthermore, two A. skirrowii isolated from the feces of related cows (R392, R393) showed similar PFGE profiles and highly similar AFLP (96.95% similarity), but less similar (>91%) ERIC-PCR profiles (Fig. 1B). The isolates R396 and R411, isolated from feces of cattle from the same farm, generated both ERIC-PCR and AFLP profiles that were more than 91% and 93% similar, respectively. In contrast, PFGE patterns showed only 85% similarity (Fig. 1C).

Comparison of PFGE, ERIC-PCR, and AFLP patterns of different Arcobacter species.
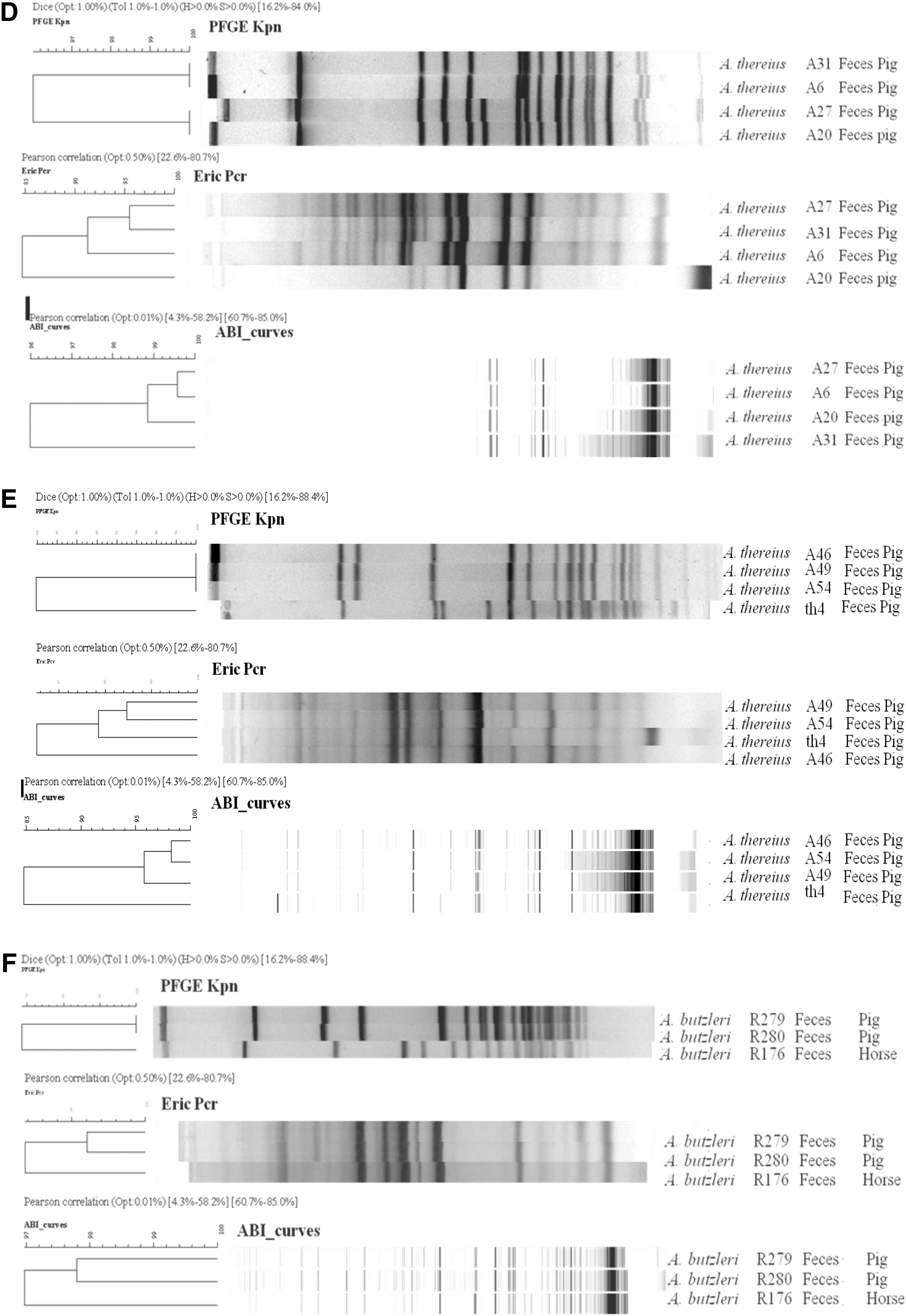

Examining four closely related A. thereius isolates (A6, A20, A27, and A31) from pig feces, PFGE showed profiles with more than 96% similarity (Fig. 1D) (De Smet et al., 2011). The isolates A20 and A27 generated different PFGE profiles in comparison to the other profiles, due to the presence of an additional band. In AFLP, all four isolates were similar when applying a 96% cut-off value. In addition, isolates A27 and A6 generated more than 99.5% similar AFLP patterns. In contrast to the PFGE and AFLP patterns, ERIC-PCR analyses showed a similarity of 95% for isolates A27 and A31, but a lower similarity between those two and isolates A6 and A20 (Fig. 1D). Also for the related A. thereius isolates A46, A49, and A54 from pig feces (De Smet et al., 2011), identical PFGE profiles were obtained, and ERIC-PCR and AFLP discriminated them at a 98% and 95% similarity level, respectively. Although a similarity of 97.8% was observed with isolate Th4 by ERIC-PCR, both AFLP and PFGE profiles showed a discordance for the same isolate compared to the A46, A49, and A54 profile (<85%) (Fig. 1E). A. butzleri R176 and R280, isolated from horse and pig feces, respectively, generated PFGE profiles with less than 70% similarity, while more than 97% similarity was obtained by AFLP and ERIC analyses (Fig. 1F). A. trophiarum strains with highly similar PFGE profiles (96.8%; strains R490, R497) showed almost 90% and 95% similarity by ERIC-PCR and AFLP, respectively (Fig. 1G).
Assessment of the in vitro genomic stability of A. butzleri and A. cryaerophilus
The genomic stability of six Arcobacter strains was evaluated by PFGE after 16 subcultivations under two different temperature and two oxygen conditions. In general, PFGE profiles of all strains were identical throughout the experiment under aerobic conditions both at 30°C and 37°C. However, variations in PFGE profiles were seen under microaerobic conditions at 37°C, except for A. butzleri R643 and A. cryaerophilus LMG 9904T (data not shown). From passage 4 on, an additional band in A. cryaerophilus strain R206 and a displacement of a single fragment in A. cryaerophilus strain R637 were detected. For strain LMG 9947, two bands disappeared while a smaller band became visible from passage 12 on (Fig. 2). From passage 2 on, a large band disappeared and an additional smaller band became visible in the profiles of A. cryaerophilus strain R671 at 37°C, both under microaerobic and aerobic conditions. The strains showing changing PFGE patterns were also screened by ERIC-PCR, but no differences in ERIC-PCR profiles could be observed.

UPGMA dendrograms from PFGE profiles using restriction with KpnI showing the assessment of the in vitro genomic stability of A. cryaerophilus strain (LMG 9947) isolated from bovine fetus.
This first experiment showed that exposure to environmental conditions of 30°C and 37°C together with microaerobic oxygen levels can induce changes in PFGE patterns. To assess whether this genomic instability already occurs at earlier passages, 10 A. butzleri and 8 A. cryaerophilus strains were cultivated for 3 generations at 30°C or 37°C under microaerobic conditions. No variations in band profiles were obtained for any of the A. butzleri strains (data not shown). With A. cryaerophilus, identical fingerprints were obtained for all strains, except for strain R861, where in both passages identical PFGE profiles at 30°C were obtained, while two new PFGE profiles were generated at 37°C, with a difference of five and seven bands on passages 1 and 3, respectively (data not shown).
Discussion
In the present study, the molecular typing methods ERIC-PCR, PFGE, and AFLP were compared in their ability with six Arcobacter species for the first time. Analysis of a set of 198 Arcobacter isolates confirmed once again the large strain diversity existing in the six Arcobacter species. No correlation between typing patterns from strains isolated from humans, animals, or food was noticed. These findings are similar to those in the multilocus sequence typing study, where no association with either host or geographical source was observed (Miller et al., 2009).
In general, with a limited number of isolates to be characterized and taking into account the recommendations by van Belkum et al. (2007), PFGE with restriction by KpnI is proposed as the first method of choice. When identical fragments for apparently unrelated isolates occur, a second PFGE analysis with the restriction enzyme NruI is recommended. The reason no fingerprints were obtained for some strains could be due to an excessive restriction by endogenous endonucleases or by DNA methylation activities (van Belkum et al., 2007). However, as PFGE is time consuming and labor intensive, ERIC-PCR represents a more convenient genomic fingerprinting technique, and has already been used for a limited number of species in the past (Houf et al., 2002; Aydin et al., 2007) and in an A. butzleri outbreak investigation (Vandamme et al., 1993). This method has the advantage of being rapid and easy to perform compared to the other two typing methods. The reproducibility of this method is often questioned, though, as shown in previous studies, the fingerprints as such cannot be exchanged but the interpretation is reliable. Nevertheless, delineation of genotypes or strains remains challenging when PCR-based methods are applied. No guidelines for genotype delineation have been proposed, and a fixed percentage above which isolates are considered identical is not possible. Within each fingerprint cluster generated by PCR-based methods, three classes of fingerprints are obtained: isolates clearly different, isolates clearly identical, and a group of fingerprints with minor variations in the presence or intensity of some amplicons. The intensity of the amplicons is often not interlaboratory reproducible, in contrast to their presence or absence (Houf et al., 2002; Van Driessche et al., 2004; Van Driessche et al., 2005). Therefore, in the case that genotype delineation is strictly necessary, isolates differing in one or two amplicons in PCR-generated fingerprints should be further examined by PFGE.
Though analysis by AFLP generates comparable results as obtained by ERIC-PCR, it is more expensive, demands specific equipment, and has been shown to be less discriminative for both A. thereius and A. trophiarum, and therefore, less recommended for Arcobacter typing.
In the present study, genomic instability due to in vitro manipulation was only observed in some strains of A. cryaerophilus, especially under microaerobic conditions at 37°C, partly explaining the enormous heterogeneity reported in this species. During transmission and the colonization of the intestinal tract of humans and animals, Arcobacter undergoes different stress factors comparable to those in the experiments, such as temperature and oxygen concentration. The Arcobacter genome seems to be exposed to relatively rapid genomic changes in vitro, but data on in vivo conditions, as performed for Campylobacter (Wassenaar et al., 1998; Ridley et al., 2008; Leblanc-Maridor et al., 2011), are lacking. One should keep in mind that these differences in profiles were not picked up by ERIC-PCR, meaning that the variation in PFGE profiles are probably caused by (point) mutations. Nevertheless, the present study showed once again that the large heterogeneity is not artificial or completely induced by laboratory actions of the typing methods applied. This still leaves the question on the origin of this diversity and the impact on epidemiological studies. Genetic alteration, if occurring in vivo, may explain the high diversity of Arcobacter present in different hosts. This variation may be caused by mutations or rearrangements in genes that are needed under stress conditions, which may increase the survival capacity of Arcobacter, but this hypothesis needs confirmation in future studies.
Footnotes
Disclosure Statement
No competing financial interests exist.