
Abstract
Salmonella is the most common cause of bacterial food poisoning in humans worldwide. Thus, rapid and reliable methods for the detection of this pathogen are required. Real-time reverse-transcriptase polymerase chain reaction (rt-RT-PCR), which detects the presence of mRNA (shorter half-life than DNA) has shown great potential for detecting viable pathogens. We recently identified a few new potential specific DNA sequences for Salmonella enterica using a comparative genomics method (Chen et al., 2010). In the present study, we examined the expression of the Salmonella-specific sigDE operon (encoding invasion proteins within the pathogenicity island 5) under typical growth conditions to determine whether sigDE could be a useful viability marker for the bacterium. We then assayed sigDE mRNA from cells heat-treated at 60°C, 100°C, and 121°C (autoclaved), and found that mRNA was degraded in autoclaved bacterial samples. These results showed that the sigDE transcript is a suitable mRNA target for rt-RT-PCR with samples pretreated at 121°C. Thus, an rt-RT-PCR using sigDE primers was developed for the detection of viable Salmonella. An RNA internal amplification control was constructed by overlap extension PCR, synthesized using in vitro transcription with a T7 RNA polymerase promoter, and incorporated into the rt-RT-PCR system to eliminate false-negative results. The rt-RT-PCR system has the capability of specifically detecting all the tested S. enterica serovars, and the detection limit of this assay with cultures of Salmonella Typhimurium ATCC 13311 was 101 colony-forming units (CFU)/mL. After 18-h enrichment, sigDE-based rt-RT-PCR could detect as low as 100 CFU/mL of Salmonella from egg broth and milk.
Introduction
S
The traditional methods used to detect Salmonella in food, which rely on laborious bacteriological and serological identification, take 4–7 days to complete (Rodriguez-Lazaro et al., 2003). In recent years, significant advances have been made in the development of molecular-based methods, particularly polymerase chain reaction (PCR)–based assays. Genes used for the identification of Salmonella by PCR detection methods include invA (encoding Salmonella invasion protein) (Ferretti et al., 2001; Malorny et al., 2003), the ttrRSBCA locus (encoding proteins for tetrathionate reduction) (Malorny et al., 2004), fimA (encoding a major fimbrial subunit) (Doran et al., 1994), stn (encoding enterotoxin) (Dinjus et al., 1997), and fliC (encoding flagellin) (Song et al., 1994). PCR-based assays are specific and sensitive, and results are obtained in a few hours. However, the main drawback with PCR methods is their apparent inability to differentiate between live pathogens and killed ones. Bacterial DNA usually remains in a food sample after the microorganisms have died, potentially leading to false-positive results. Ethidium bromidemonoazide (EMA) and propidium monoazide (PMA) are dyes that can bind to DNA of dead cells and prevent its amplification by PCR. An EMA (or PMA) staining step prior to PCR allows for the effective inhibition of false-positive results caused by the presence of dead cells (Wang and Mustapha, 2010). However, EMA/PMA staining is technically difficult and poisonous to the operator.
mRNA is considered a more appropriate target than DNA to assess cell viability since most mRNA species have a short half-life of only a few minutes (Coutard et al., 2005; Liu et al., 2010). Thus, the application of reverse-transcriptase PCR (RT-PCR) for the amplification of mRNA targets, as opposed to PCR amplification of genomic DNA, has greater potential for ensuring that the target microorganisms are viable (Maurer, 2006). The use of mRNA as a target has been applied to the detection of Salmonella Enteritidis based on the sef gene (Szabo and Mackey, 1999). Recently, several studies showed that the invA mRNA is a candidate for detection of Salmonella spp. by real-time RT-PCR (rt-RT-PCR) (Fey et al., 2004; Techathuvanan and D'Souza, 2011) and conventional RT-PCR (Jacobsen and Holben, 2007). However, the invA gene has been reported to be absent in some strains of Salmonella Litchfield, Salmonella Senftenberg (Rahn et al., 1992) and Salmonella Poona (Dione et al., 2011). To be used as a detection target and indicator of cell viability, the gene should be present in all strains and specific to the target pathogen, and its mRNA should be constitutively expressed and have a short half-life in dead cells. Therefore, it is necessary to identify and carefully evaluate new targets for the detection of living Salmonella.
Abundant genomic sequence information makes it possible to apply bioinformatics tools for identification and development of specific and sensitive detection targets for bacterial pathogens. We have recently identified a few new potential specific sequences for Salmonella enterica using a comparative genomics method, and several species-specific target sequences were evaluated for specificity and sensitivity (Chen et al., 2010). In the present study, we examined the expression of the Salmonella-specific sigDE operon under a range of conditions, and evaluated the stability of mRNA by thermal treatments, to demonstrate that sigDE was a suitable mRNA target for rt-RT-PCR. The sigDE operon is located on a Salmonella pathogenicity island (SPI)-5, a 7.6-kb chromosomal locus known to play a role in enteropathogenesis of Salmonella. The sigD and sigE genes form an operon (Wood et al., 1998). The sigD was identified as invasion gene, and the encoded protein is translocated into eukaryotic cells to promote fluid secretion and inflammatory response (Wood et al., 1998). SigE acts as a chaperone for secretion of SigD (Hong and Miller, 1998). Our previous study showed that sigDE was present in all tested Salmonella enterica serovars (Chen et al., 2010). The goal of this study was to develop a sigDE-based rt-RT-PCR assay for viable Salmonella enterica with increased sensitivity and specificity.
Materials and Methods
Bacterial strains
Salmonella enterica serovar Typhimurium (Salmonella Typhimurium) ATCC 13311 and Salmonella enterica serovar Abony (Salmonella Abony) NCTC 6017 were used for evaluation of the sigDE rt-RT-PCR assay. For rt-RT-PCR specificity testing, 147 S. enterica strains, including 21 strains from culture collections and 126 isolates from clinical and food samples, and 28 non-Salmonella bacteria were used (Table 1). Bacterial strains were kept in our laboratory. Salmonella spp. were grown in tryptic soy broth (TSB; Difco, Sparks, MD), and other bacterial species were grown in Brain Heart Infusion (BHI) broth (BD, NJ) overnight at their optimal temperatures.
+, rt-RT-PCR positive; −, rt-RT-PCR negative.
Primer design
A Salmonella-specific genetic locus, the sigDE operon (NCBI accession no. AF021817), was selected for development of the rt-RT-PCR assay based on previous research from our laboratory (Chen et al., 2010). Primers for amplification of sigDE as well as 16S rRNA and recA were designed using Primer Premier 5.0 software (Premier Biosoft Intl., CA). Primers are listed in Table 2. The 16S rRNA and recA were used as controls.
RNA extraction and rt-RT-PCR
Total RNA was extracted from cultures of the bacteria using the RNAprep pure Bacteria kit (TianGen Biotech, Beijing, China) according to manufacturer's instruction. The RNA extract was treated with DNase I (Takara BioTech [DaLian], DaLian, China). RNA purity and quantity were determined spectrophotometrically
Fifty microliter reactions contained 5 μL diluted RNA extracts, PrimeScript® RT-PCR Perfect Real Time Kit reagents (Takara BioTech) with SYBR Green I, 25 pmol of oligo (dT), and 50 pmol of random primers. Mixtures were incubated at 37°C for 30 min and then inactivated at 85°C. Real-time PCR was performed using the SYBR® Premix Ex Taq™ kit (Takara BioTech), cDNA template, 0.02 μmol of each primer, and 0.06 mg/mL bovine serum albumin. Optimal cycling conditions were at 50°C/2 min, 95°C/10 min, followed by 40 cycles at 95°C/45 s, 58°C/45 s, and 72°C/45 s, and final extension at 72°C/7 min in an ABI 7500 rt-PCR system (Applied Biosystems, Foster City, CA). The ABI 7500 Software v2.0.1 was used to determine cycle threshold (Ct) values, melting temperature (Tm), and the standard curve.
Detection of mRNA in cells at different growth periods
Salmonella Typhimurium ATCC 13311 and Salmonella Abony NCTC 6017 cultures were grown overnight. Growth during a 24-h period (growth cycle) was monitored by plate count. One-milliliter samples were collected at 3 h, 5 h, 9 h, 11 h, 15 h, and 24 h for RNA isolation and DNase-treatment. The RNA samples were used for rt-RT-PCR to investigate the effect of physiological state on expression of the sigDE operon.
Heat inactivation
Mid-log phase cells (∼108 colony-forming units [CFU]/mL) of Salmonella Typhimurium ATCC 13311 and Salmonella Abony NCTC 6017 suspended in Tris-EDTA buffer were placed in either a 100°C water bath for 20 min or 60°C water bath for 30 min or autoclaved at 121°C for 15 min. Portions of heat-treated cells were plated onto tryptic soy agar (TSA; Difco, Sparks, MD) plates and incubated at 37°C for 48 h to determine whether the heat-inactivation process was complete. Following treatment, cells were serially diluted (107 CFU/mL to 100 CFU/mL) and held at room temperature for the remainder of the experiment. Total RNA was extracted before and 2 h after the heat treatment, and used in the sigDE-based rt-RT-PCR assay. This experiment was conducted twice in triplicate.
Preparation of the internal amplification control (IAC)
Based on the sequence exclusivity to bacterial genomes, a 243-bp segment of the Chlorella protothecoides strain CS-41 ζ-carotene desaturase (zds) mRNA (NCBI accession no. GU269622) was selected to construct a RNA IAC. The IAC was constructed as previously described (D'Souza et al., 2009). A 309-bp DNA sequence with a T7 RNA polymerase promoter sequence driving transcription of the zds gene was constructed by sequential PCR using three primer sets (zds-F/zds-R, sigDE-zds-F/sigDE-zds-R and T7-sigDE-F/sigDE-R [Table 2]). In vitro transcription of this 309-bp product using a T7 in vitro Transcription Kit (TIANDZ, Beijing, China) produced the RNA IAC. An optimal IAC concentration for the rt-RT-PCR was assessed to be 3 fg/mL. The IAC amplification product (Tm of 88.9°C) was clearly distinguishable from target products (Tm of 83.0°C).
Real-time RT-PCR specificity and sensitivity
Specificity of the sigDE primers was tested against a total of 147 strains of Salmonella enterica and 28 non-Salmonella bacterial strains (Table 1) by rt-RT-PCR with the presence of the IAC. For sensitivity testing, total RNA was extracted from serial dilutions of Salmonella Typhimurium ATCC 13311 cells and tested by rt-RT-PCR. Sensitivity evaluations were done twice in triplicate with or without the presence of the IAC. PCR efficiencies were calculated from the standard curves using the following equation: PCR efficiency=[10(−1/M)] −1, where M is the slope of the standard curve (Rasmussen, 2001).
Artificial contamination of egg broth and milk
Raw Salmonella-free eggs were purchased from a local market and egg white and yolk were beaten to obtain a homogeneous mixture. Ten milliliters of the egg broths and 10 mL of milk were artificially inoculated with different final concentrations of Salmonella Typhimurium ATCC 13311 viable cells (10−1 to 102 CFU/mL), and then diluted with 90 mL buffered peptone water. The samples were incubated at 37°C for 0, 6, 18, and 24 h. One milliliter from each enriched sample was centrifuged to harvest cells, which were subjected to RNA extraction and rt-RT-PCR detection with the presence of an IAC. This experiment was conducted twice in triplicate.
Results
Expression of 16S rRNA, recA, and sigDE mRNA during the growth of S. enterica
Results of the rt-RT-PCR analysis showed that the growth phase of the cell influences the levels of some mRNA targets examined (Fig. 1). 16S rRNA and recA are housekeeping genes and were examined as controls. Based on the Ct values and melting curves, 16S rRNA, recA mRNA, and sigDE mRNA were detected in all samples from early exponential to late stationary phase. The 16S rRNA was detected at all time points with a Ct <8. For recA mRNA detection, the order of Ct values was Ct 20.5 (3 h)<Ct 21.5 (5 h, 9 h, and 24 h)<Ct 22 (11 h)<Ct 23 (15 h). For sigDE mRNA detection, the order of Ct values was Ct 20.5 (5 h)<Ct 21.6 (9 h, 11 h)<Ct 23.5 (3 h)<Ct 26.5 (15 h, 24 h).
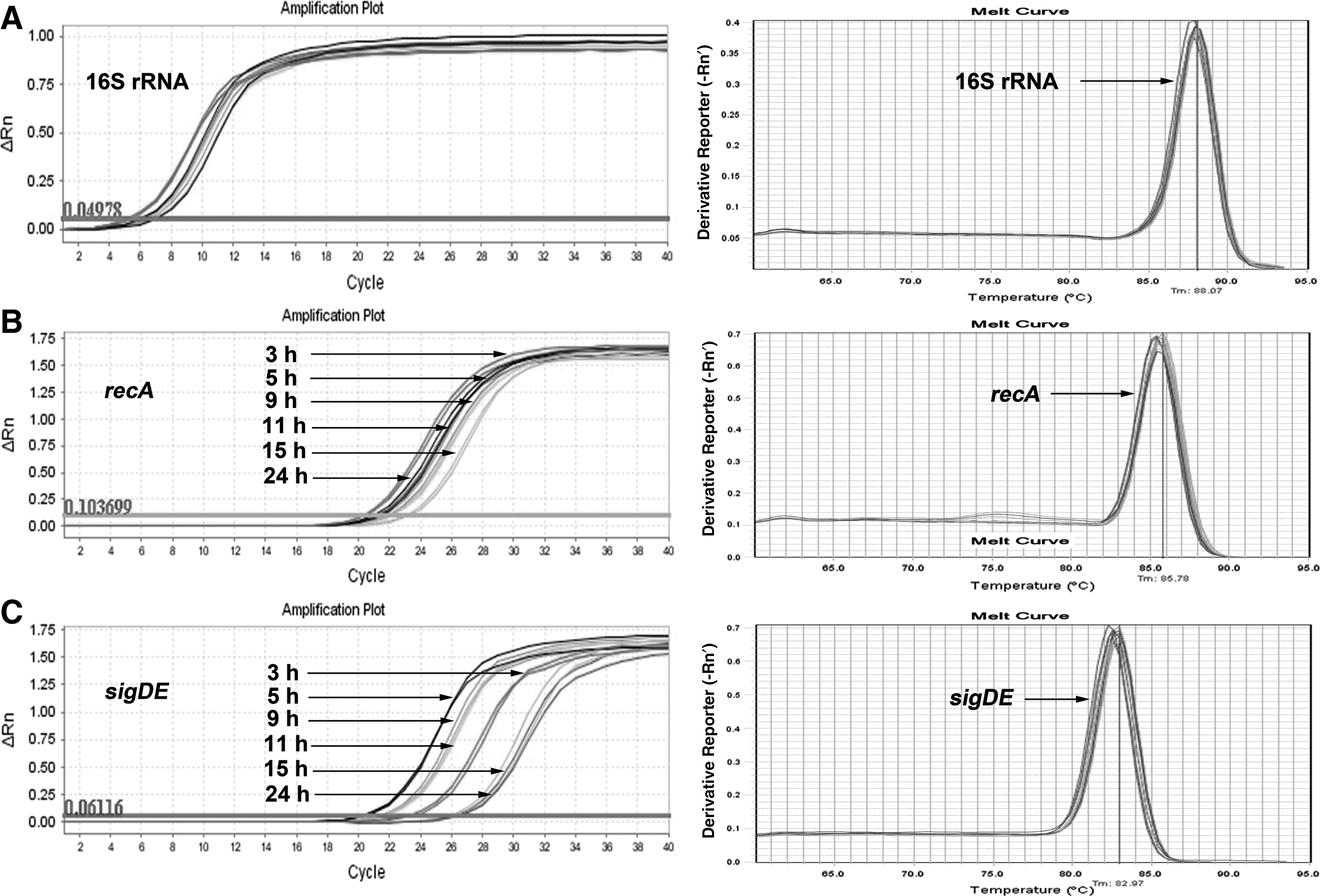
Amplification and melting temperature curves of real-time reverse-transcriptase polymerase chain reaction using 16S rRNA
Ability of the sigDE-based rt-RT-PCR assay to discriminate between viable and nonviable S. enterica cells
Thermal methods (heating at 60°C for 30 min or 100°C for 20 min and autoclaving at 121°C for 15 min) were used to kill Salmonella cells. No growth was observed on plates inoculated with treated samples following incubation at 37°C for 48 h, indicating that inactivation treatments were effective. The results of stability of sigDE mRNA following heat treatment, based on rt-RT-PCR assay, are shown in Figure 2. The sigDE-based rt-RT-PCR using mRNA prepared from autoclaved heat-inactivated cells (107 CFU/mL—100 CFU/mL) showed that no sigDE amplification products were obtained (Fig. 2D). The sigDE target was identified by rt-RT-PCR on 107 CFU/mL cells heat-inactivated at 60°C, and 107 CFU/mL, 106 CFU/mL as well as 105 CFU/mL cells heat-inactivated at 100
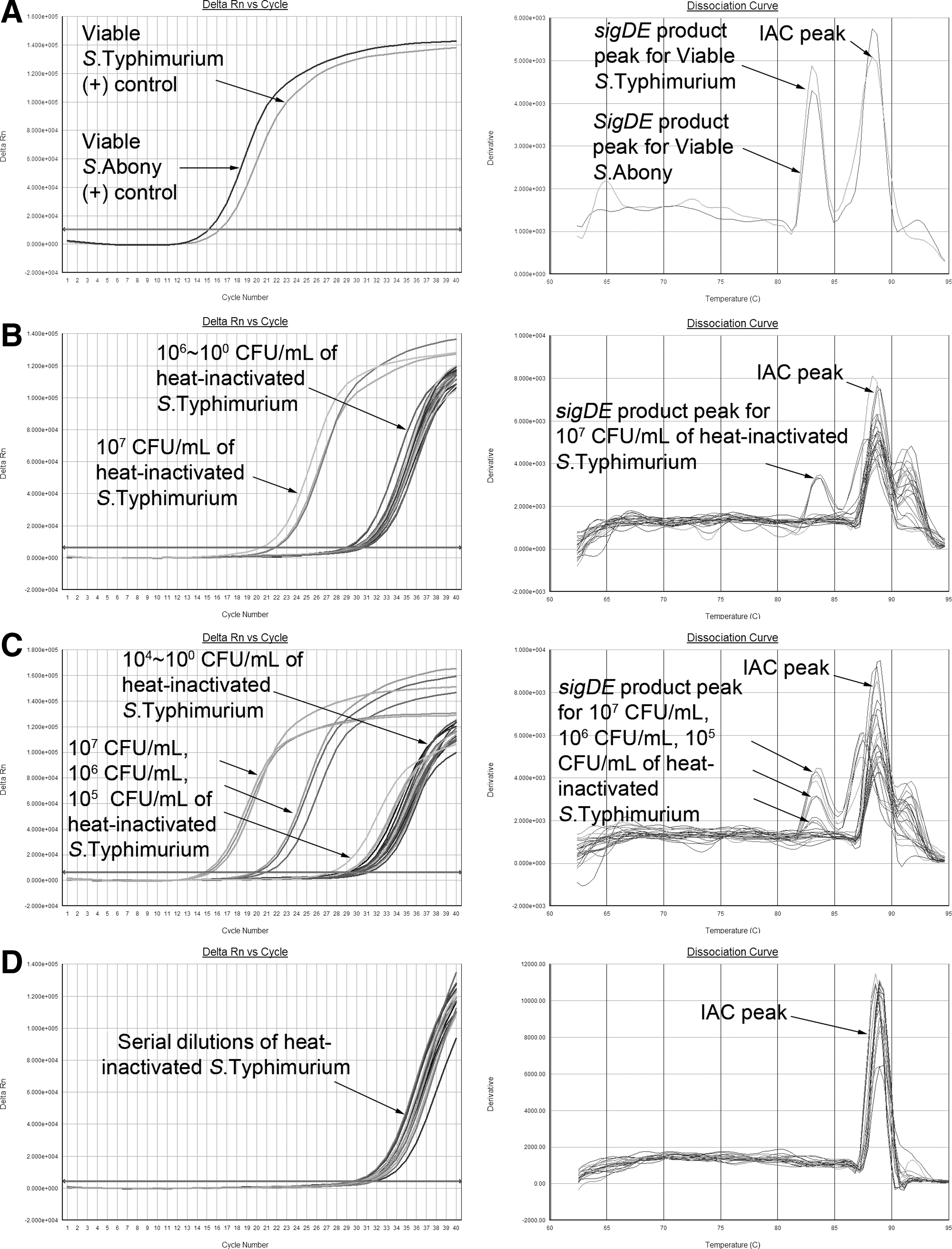
Amplification and melting temperature curves of sigDE-based real-time reverse-transcriptase polymerase chain reaction using RNA extracts from Salmonella Typhimurium ATCC 13311 and Salmonella Abony NCTC 6017 viable cells
Limit of detection
Using RNA extracted from serially diluted pure cultures, a standard curve for the rt-RT-PCR assay was obtained with a linear range from 104 to 101 CFU/mL for Salmonella Typhimurium ATCC 13311 (Fig. 3A). The reaction efficiency was 89.6% and the correlation coefficient was 0.997. It was shown that the detection limit of the sigDE-based rt-RT-PCR assay for Salmonella Typhimurium ATCC 13311 was 101 CFU/mL. A specific amplification product showing a Tm peak at 83°C was obtained for sigDE target.
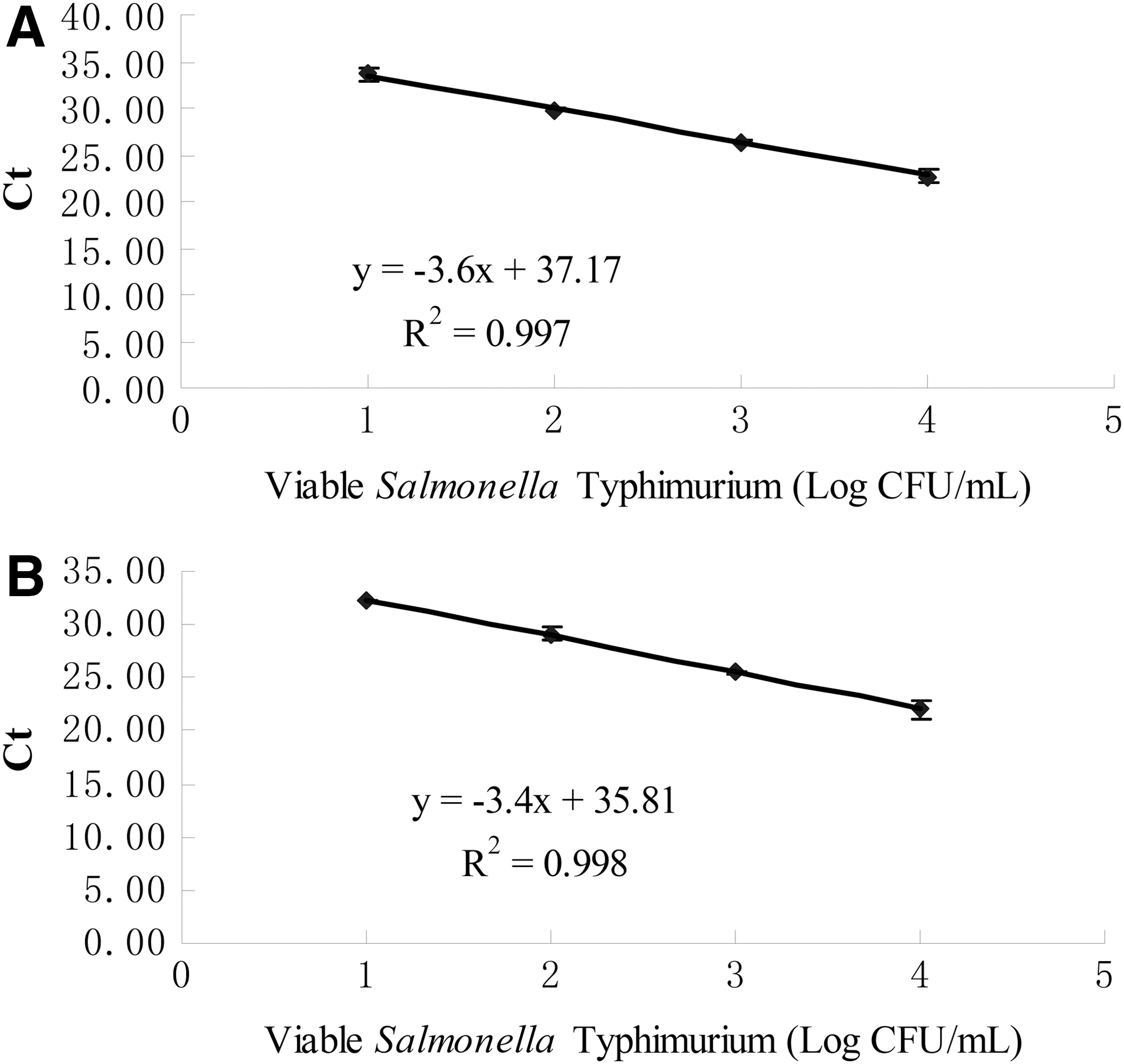
Standard curve for sigDE-based real-time reverse-transcriptase polymerase chain without RNA internal amplification control (IAC)
An RNA IAC was constructed for incorporation in the rt-RT-PCR assay to eliminate false-negative results. When the IAC sequence was searched against NCBI nucleotide collection (nr/nt) database using BLAST, no hits of bacterial genomic DNA were obtained. In addition, the IAC primers did not show any amplification from the genomic DNA of 175 different bacterial strains (representing 25 genera and 47 species) when tested by standard PCR (data not known), suggesting a high exclusivity of the IAC sequence to bacterial DNA. It was necessary to optimize the amount of IAC to be added so that the interference between the targets and the IAC would be minimized. A final concentration of 3 fg/mL of the newly constructed RNA IAC was found to be optimal for the assay. This optimization resulted in a detection limit of 101 CFU/mL for Salmonella Typhimurium ATCC 13311 (Fig. 3B). The reaction efficiency was 94.7% and the correlation coefficient was 0.998 under these conditions (Fig. 3B). This showed that the presence of the small of amount of the IAC RNA did not affect the measured efficiency of sigDE-based rt-RT-PCR. Two distinct melt peaks at 83°C and 88.9°C were obtained for the sigDE target and IAC, respectively.
Specificity of rt-RT-PCR assay using sigDE primer
The mRNA prepared from a panel of 147 bacteria (Table 1) was used to determine the specificity of rt-RT-PCR assay in the presence of the RNA IAC using sigDE primers. Twenty serovars of Salmonella enterica representing different serogroups were tested and all showed positive results with specific Tm peaks at 83°C. Non-Salmonella bacteria (14 genera and 25 species) only showed amplified products with Tm peaks at 88.9°C representing IAC amplified products, indicating the absence of PCR inhibition. Water was used for negative controls and also showed only the IAC amplification product.
Artificially contaminated samples
To further validate the applications of the rt-RT-PCR assay in food, the egg broths and milk were artificially inoculated with different concentrations of Salmonella Typhimurium ATCC 13311 viable cells and enriched for 0, 6, 18, and 24 h. Salmonella detection limits of inoculated egg broth were assessed to be 102 and 100 CFU/mL after 6 h and 18 h enrichment, respectively (Table 3). After 18 h of enrichment, rt-RT-PCR generated positive results from artificially contaminated milk at an inoculum level of 100 CFU/mL (Table 3).
CFU, colony-forming unit;+, rt-RT-PCR positive; −, rt-RT-PCR negative.
Discussion
The rt-RT-PCR using sigDE primer has a detection limit of 101 CFU/mL and is capable of detecting all the tested serovars (e.g., Paratyphi A [Group A], Typhimurium [Group B], Infantis [Group C1], Enteritidis [Group D], Anatum [Group E], Aberdeen [Group F], and Poona [Group G]). The inclusion of the IAC did not affect either the amplification or the detection limit of the rt-RT-PCR assay. The detection limits of the assay in the presence of internal amplification controls were also 101 CFU/mL.
The inclusion of IAC in rt-RT-PCR is needed to indicate the presence of PCR inhibitors that can cause false-negative results for Salmonella enterica–positive samples (Hoorfar et al., 2004; Hartman et al., 2005). PCR inhibitors may be lipids, proteins, enzymes, chemical additives, fiber, other bacteria, or pH (Lee et al., 2006). Therefore, we developed an RNA IAC. The size of the IAC used in the present study was smaller than the sigDE target amplicon, making it easy to differentiate the two amplicons by difference in their Tms.
In Salmonella enterica grown in TSB, the 16S rRNA, the recA mRNA, and sigDE mRNA were detectable by rt-RT-PCR throughout the growth curve. The expression of the sigDE operon was stable in the logarithmic phase and early stationary phase. In nonhost environments, Salmonella is most likely to persist in a starved and highly stressed state, which might not produce sigDE mRNA. However, for nearly all detection methods for foodborne bacteria, food samples are enriched (Feder et al., 2001). This step increases cell number and consequently the total mRNA production. Techathuvanan and D'Souza (2011) developed a method to detect viable Salmonella Enteritidis from spiked liquid whole eggs within 24 h using SYBR Green I–based rt-RT-PCR targeting the invA gene. Salmonella Enteritidis could be detected from liquid whole eggs down to 104 and 102 CFU/25 mL after enrichment for 6 and 12 h, respectively. In our study, the sigDE-based rt-RT-PCR detection limits of inoculated egg broth were assessed to be 102 and 100 CFU/mL after 6 h and 18 h enrichment, respectively. The whole procedure is much faster than traditional culture-based detection that takes several days. Thus, a pre-enrichment culture step can provide an essential preliminary step to achieve reliable and sensitive detection of Salmonella enterica. In this case where culture enrichment is used, the number of dead cells is expected to be low and rt-RT-PCR would lack some of its advantage over PCR. However, in case either the enrichment is not needed or amount the of dead cells is much more than that of live cells, rt-RT-PCR would generally have the ability to distinguish live cells from dead ones.
Numerous studies, performed to establish the use of RNA detection as proof of bacterial viability, employed heating of the cells (Fykse et al., 2007). Heating has been suggested to disrupt the bacterial cell wall, facilitating the release of cellular components and exposing RNA to RNases (McKillip et al., 1999). In our study, the sigDE mRNA target was detected by rt-RT-PCR with cells (107 CFU/mL) heat-inactivated at 60°C, and cells (107 CFU/mL–105 CFU/mL) heat-inactivated at 100°C, which could be attributed to some persistent sigDE mRNA. Persistence of mRNA in dead cells is found to be dependent on the method of cell inactivation, the mRNA species, and subsequent holding conditions of the cells (Sheridan et al., 1999). The sigDE mRNA appeared to be degraded more completely in cells autoclaved at 121°C than those treated at 100°C or 60°C water bath.
In this study, we have developed an rt-RT-PCR method based on a new detection target, sigDE. The method has the potential to be used as a preliminary screening tool for detecting viable S. enterica. This assay will reduce the overall time and resources expended in the lab because only positive samples containing viable cells will be processed. Using this method, accurate and reliable data for risk assessments can be achieved.
Footnotes
Acknowledgments
This work was jointly supported by grant nos. 31230058, 31150001, 31000779, and 31101246 from the National Natural Science Foundation of China, grant no. 2012BAD29B02-2 from the Ministry of Science and Technology of China, and grant no. 11DZ0502700 from Science & Technology Commission of Shanghai Municipality.
Disclosure Statement
No competing financial interests exist.