
Abstract
Air and nitrogen gas are commonly used feed gases for plasma generation and are economically useful in industrial applications. The two gases were compared in dielectric barrier discharge plasma for the inactivation of Campylobacter jejuni on an agar surface. Plasma treatment with nitrogen gas for 20 s did not yield any reduction (p>0.05) in viable cell count. However, a 0.8-log reduction (p<0.05) in colony-forming units (CFU) occurred when the nitrogen gas was supplemented with 2% (vol/vol) air. The use of air only, air supplemented with 2% (vol/vol) nitrogen, or oxygen only further decreased the viable cell counts by 0.7–1.7-log CFU (p<0.05). These results suggest that oxygen in plasma generation is critically important for the increased inactivation effect. Scanning electron microscopy analysis showed much cell debris including fragmented flagella in the sample exposed to air plasma, while no cell debris was found in the sample exposed to nitrogen plasma. In transmission electron microscopy analysis, many C. jejuni cells exposed to air plasma had truncated flagella with sharp bends, while the cells exposed to nitrogen plasma were normal, strongly suggesting that the air plasma can reduce the virulence of C. jejuni. A BacLight assay showed that air plasma damaged the cellular membrane (p<0.05), whereas nitrogen plasma did not after 5- or 20-s treatment. The damage to the membrane was consistent with the reduced viable cell count. Based on confocal microscopic analysis, the similar results were found by visualizing the fluorescent-dye-stained cells. In addition, the prolonged nitrogen plasma for 2 min also damaged many cellular membranes. This study shows that air, especially oxygen, is more effective and destructive than nitrogen and provides evidence that membrane damage may be a major mechanism for the inactivation of C. jejuni exposed to plasma.
Introduction
C
Several factors need to be considered to improve the bactericidal efficiency of plasma treatment. They include cell density (Fernández et al., 2012), relative humidity (Ragni et al., 2010), sample surface features (Fernández et al., 2013), and plasma generation parameters such as flow rate, feed gas composition, and power input (Niemira and Sites, 2008; Yang et al., 2011; Niemira, 2012b). Air, oxygen, nitrogen, argon, and helium are commonly used feed gases for plasma generation. Because argon and helium are expensive, both air and nitrogen may have greater potential for industrial applications. Even though air gas has been often used to generate plasma (Critzer et al., 2007; Leipold et al., 2010; Niemira and Sites, 2008), nitrogen gas has been used as often as air for inactivation studies of bacterial pathogens (Boudam et al., 2006; Fernández et al., 2012, 2013; Mols et al., 2013).
Gas composition can make a difference in the inactivation of foodborne pathogens (Niemira, 2012b); however, related information is still limited, and more information is necessary to better understand the effect of gas composition on bacterial pathogens.
C. jejuni is a major foodborne pathogenic bacterium worldwide and is highly associated with poultry products such as chickens and turkeys (Young et al., 2007; Scallan et al., 2011). Approximately 0.85 million illnesses are estimated to occur annually in association with C. jejuni in the United States alone (Scallan et al., 2011). In this study, we compared air, nitrogen gas, and other gas compositions in plasma generation to understand their effects on the inactivation of C. jejuni.
Materials and Methods
C. jejuni growth conditions and cell preparation
C. jejuni NCTC11168 (American Type Culture Collection, Manassas, VA) was used in this study. It was stored in glycerol at −70°C and routinely grown on tryptic soy agar supplemented with 5% sheep blood (TSAB) at 37°C under microaerobic conditions (6%–12% O2, 5%–8% CO2) generated using AnaeroPack-MicroAero (Mitsubishi Gas Chemical Co., Tokyo, Japan). After 32–48 h of incubation, the cultures were suspended in Mueller-Hinton Broth at an optical density (OD) of 0.1–0.2 at 600 nm. An aliquot (100 μL) of the suspension was spread on TSAB plates and grown for an additional 14–15 h under the same conditions. Using cell scrapers, the culture was finally suspended in Mueller-Hinton Broth as appropriate and then used for plasma treatment on the agar surface.
Plasma apparatus and treatment conditions
A DBD plasma under atmospheric pressure was used in this study (Fig. 1). Air (clean dry air) or nitrogen gas was fed into the top of the plasma generation head in which the electrode was installed (Fig. 1a). The gas discharge took place between a powered electrode and a ground electrode at 5.5 kV and 30 kHz (Fig. 1b). The gas flow rate was 20 L/min. A dielectric barrier made of ceramic was placed at the side of the powered electrode, and the gap between the dielectric barrier and ground electrode measured 0.5 mm. The generated plasma was flushed downward through the outlets, and the samples on the sample pedestal were exposed to the plasma. The distance between the gas discharge and the sample was approximately 2.5 cm.
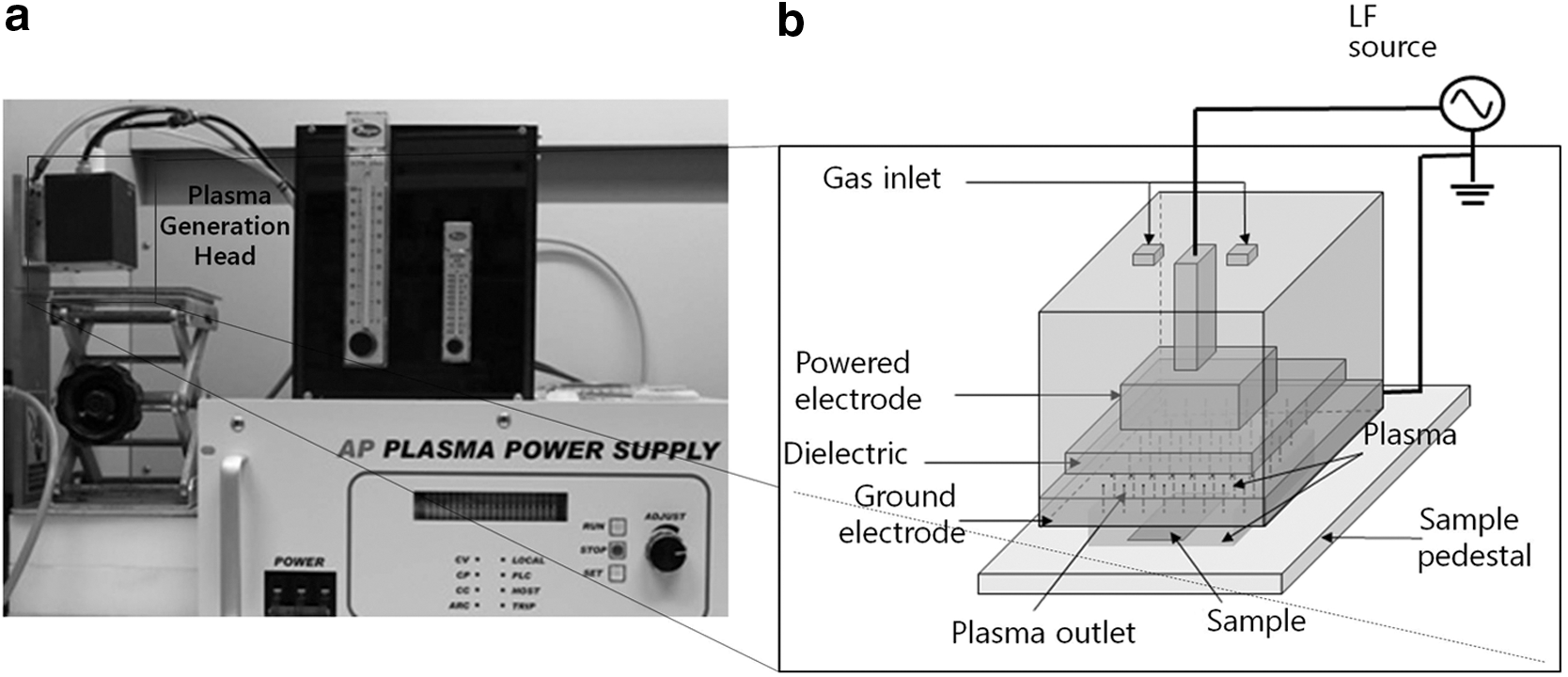
Dielectric barrier discharge plasma device used in this study.
Plasma treatment and viable cell counts
Aliquots (20 μL, 106–107 CFU) of the suspended cells at OD 0.1–0.2 were gently spread with the flat side of a pipette tip onto the surface of plate count agar (BD, Sparks, MD) in a 2.4-cm-diameter circle. Then, the plates were air-dried until no moisture was left on the agar surface and exposed to the plasma treatment for a specified time period. After plasma treatment of the samples, the bacterial cells were suspended in 10 mL of sterile 0.85% NaCl solution from the agar plates using cell scrapers, vigorously agitated using vortex, serially diluted in sterile 0.85% NaCl solution by 10-fold to 10−4, and then spread (100-μL aliquots of each dilution including 100) on TSAB plates. The plates were incubated at 37°C under microaerobic conditions for 2 days, and colonies were counted to determine the number of viable cells. By this method, around 80% of the initially inoculated cells were recovered from the plates.
Scanning electron microscopy (SEM) analysis and measurement of optical density
For SEM sample preparation, the suspended cells were concentrated to an OD of ∼5.0 (∼1010 CFU/mL) by centrifugation. Approximately 109 CFU (100 μL) were gently spread on plate count agar in a 3.5 cm diameter circle, air-dried, and treated with plasma for 2 min. After plasma treatment of the samples, the bacterial cells were suspended in 5 mL of phosphate-buffered saline. An aliquot of the samples was measured for OD at 600 nm, and the remainder was concentrated by 100-fold by centrifugation for SEM analysis. The concentrated cells were fixed in a 2.5% paraformaldehyde-glutaraldehyde mixture buffered with 0.1 M phosphate (pH 7.2) for 2 h. Then, the samples were postfixed in 1% osmium tetroxide in the same buffer for 1 h, dehydrated in graded ethanol, substituted by isoamyl acetate, and dried at the critical point in CO2. Finally, the samples were sputtered with gold in a sputter coater (SC502, Polaron) and observed under SEM, Hitachi S4300N (Hitachi, Japan), installed at the Korea Research Institute of Bioscience & Biotechnology.
Transmission electron microscopy (TEM) analysis
The samples were stained with uranyl acetate and lead citrate, then examined under CM 20 transmission electron microscope (Philips, Netherlands).
Measurement of cell membrane integrity by microplate reader
Aliquots (100 μL, ∼108 CFU) of the suspended cells at OD 0.6–0.8 were gently spread on plate count agar in a 3.5-cm diameter circle and air-dried. After plasma treatment for 5 or 20 s, the cells were suspended in 5 mL of sterile 0.85% NaCl, and cell membrane integrity was measured using LIVE/DEAD BacLight Bacterial Viability Kits (Molecular Probes, Eugene, OR) following the manufacturer's protocol. Briefly, 2X stain solution was prepared by diluting SYTO 9 and propidium iodide dyes to 0.01 mM and 0.06 mM, respectively. Then, the cell suspension and 2X stain solution were mixed at a 1:1 ratio at 100 μL each in a microplate. After 15-min incubation of the reaction in darkness, the fluorescence intensity was measured in triplicate using an Infinite® M200 PRO microplate reader (Tecan, Switzerland). The excitation wavelength was 485 nm. The emission wavelengths for green and red fluorescence were 530 nm and 630 nm, respectively. The cell membrane integrity was calculated using the ratio of green to red fluorescence.
Confocal microscopic analysis
Fluorescent dye–labeled C. jejuni cells were viewed using a ZEN 2009 confocal laser scanning microscope (model LSM710; Carl Zeiss Microscopy GmbH, Germany). The cells were visualized using a water-immersion objective, C-Apochromat 40×/1.20 W Korr M27, and a 2×scan zoom. The cells were prepared, plasma-treated, harvested as in the microplate reader experiment, 20-fold concentrated by centrifugation, and the fluorescent dyes were added to 0.0056 mM and 0.033 mM for SYTO 9 and propidium iodide, respectively. A fraction of the cells (5 μL) was visualized under the microscope.
Statistical analysis
For multiple comparison, the data were analyzed via analysis of variance and significant differences among the treatments were compared with Duncan's multiple range test at p=0.05 using the SAS statistical program (SAS Institute Inc., Cary, NC). For comparison of two treatment groups, a Student t-test was employed for any statistical difference at p=0.05 by using Microsoft Office Excel 2007.
Results
The effect of feed gas composition on the inactivation of C. jejuni was studied using a DBD plasma apparatus. Air, N2, O2, and mixtures of air and N2 (air supplemented with 2% N2 or N2 supplemented with 2% air) were compared to inactivate C. jejuni on an agar surface after 20-s treatment (Fig. 2a). Gas flow with air or N2 in the absence of plasma generation did not inactivate C. jejuni at all compared to no treatment control. Similarly, plasma treatment with only N2 gas did not decrease the viable cell counts. However, when N2 gas was supplemented with 2% air for plasma generation, it reduced the viable cell count by 0.8-log CFU compared to no treatment control in C. jejuni (p<0.05). When the concentration of air was increased in the feed gas, there was a further reduction in viable cell counts. Specifically, plasma treatment with mostly air, or with only air or O2 further reduced the viable cell counts by 0.7- to 1.7-log CFU compared to N2 gas supplemented with 2% air (p<0.05) (Fig. 2a). These data indicate that O2, but not N2, is effective for the inactivation of C. jejuni in short-term (20 s) treatment. Then, the effect of long-term plasma treatment (1–4 min) was studied by comparing air to N2 gas (Fig. 2b). In the long-term treatment, the N2 plasma treatment was somewhat effective with about 2-log reduction after 4-min treatment. However, the enhanced inactivation effect of air compared to N2 gas was also noticeable (p<0.05). With air plasma, about 3-log reduction was observed after only 1-min treatment, and the CFU levels fell below the detection limit after 4-min treatment (>4-log reduction). The effectiveness of DBD plasma using air was also confirmed by using Staphylococcus aureus. Approximately 1.3- and 2.6-log reductions occurred after 5-s and 1-min treatments of S. aureus, respectively. Those were comparable to 1.5- and 3-log reductions, respectively, for C. jejuni.
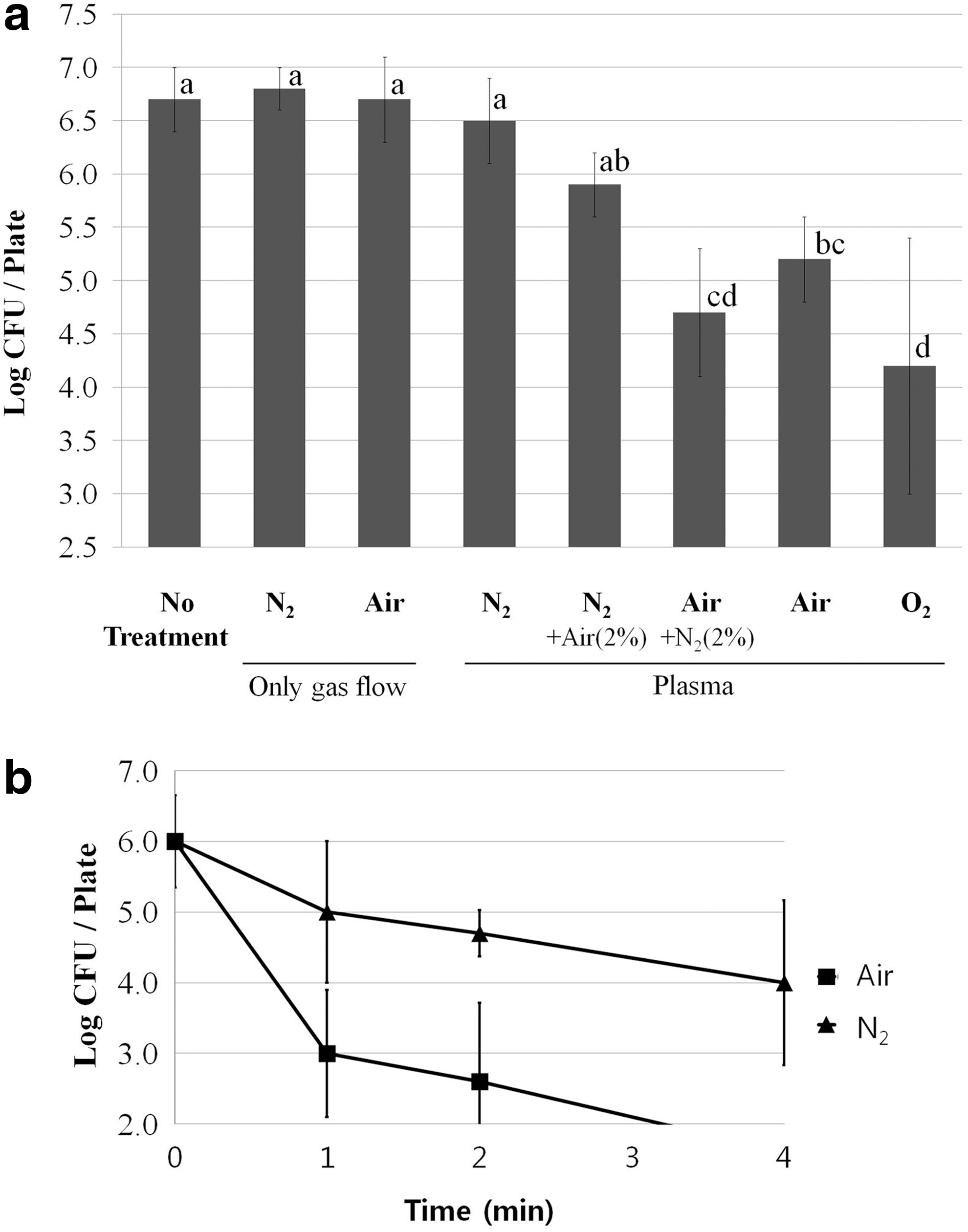
Comparison of air, nitrogen, and oxygen plasma for the inactivation of C. jejuni on agar plates.
To further understand the effects of air and N2 plasma on the morphology of C. jejuni cells, SEM analysis was implemented (Fig. 3). Compared to no treatment control, the cells treated with air plasma were damaged, and many fragments of cell debris were found. Some of the cell debris appeared to be fragmented flagella. In contrast, the cells treated with N2 plasma were not damaged, similar to no treatment control. Cellular damage was also confirmed by suspending plasma-treated cells and measuring the OD of the suspension at 600 nm. The OD values of the no-treatment control and the N2 plasma-treated samples were 0.15 and 0.13, respectively. In contrast, the OD of the air plasma-treated sample was only 0.065, far less than the others. The effects of air and N2 plasma on the flagellar structure of C. jejuni were further studied by analyzing TEM images for individual cells (Fig. 4). Many C. jejuni cells exposed to air plasma for 20 s had truncated flagella with sharp bends (Fig. 4b), compared to long, smooth, and straight flagella in the samples of no-treatment control (Fig. 4a) and N2 plasma (Fig. 4c).
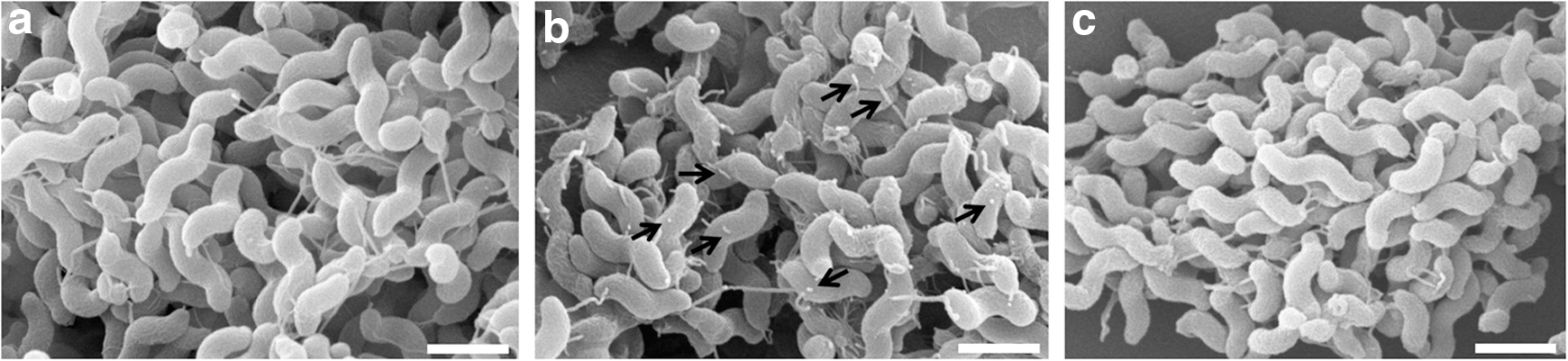
The effect of air and nitrogen plasma on the cell morphology of Campylobacter jejuni in scanning electron microscopy analysis (scale bar=1 μm; arrows indicate cell debris).
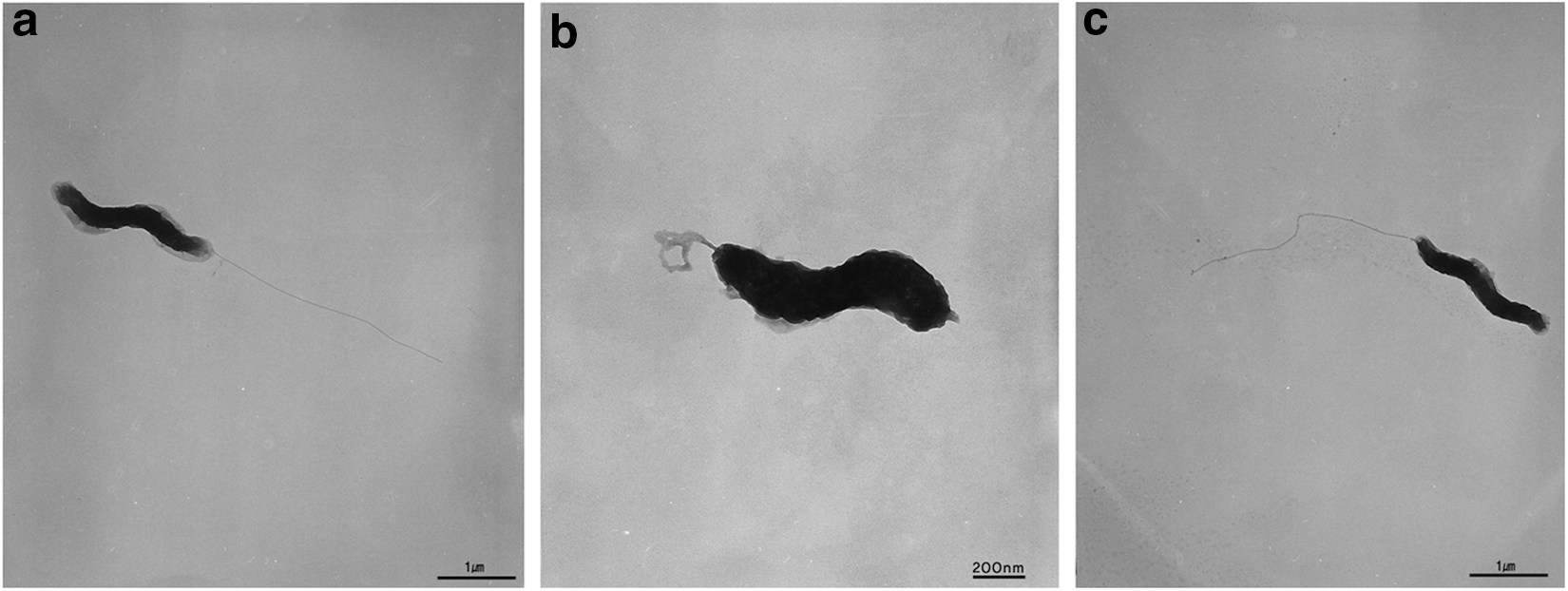
The effect of air and nitrogen plasma on the cell morphology of Campylobacter jejuni in transmission electron microscopy analysis:
Because the cellular membrane is the most exposed to plasma treatment, its impairment in C. jejuni was studied after plasma treatment (Fig. 5a). The membrane integrity was studied using two different fluorescent dyes: SYTO 9 green-fluorescent (G) nucleic acid stain and the red-fluorescent (R) nucleic acid stain, propidium iodide. The bacterial cells with intact membranes are labeled only with SYTO 9, whereas the bacterial cells with damaged membranes are labeled both with SYTO 9 and with propidium iodide. The fluorescence intensity of the SYTO 9 stain is reduced when the propidium iodide stain is present. Thus, the membrane integrity of C. jejuni cells treated with air or N2 plasma was studied by comparing the ratio of green (G) to red (R) fluorescence in the samples. The ratio was lower for the C. jejuni samples treated with air plasma for 5–20 s compared to no-treatment control (p<0.05) (Fig. 5a). In contrast, the ratio did not change for the C. jejuni samples treated with N2 plasma for 5 or 20 s (p>0.05) (Fig. 5a). These results clearly show that air plasma is destructive to the cellular membranes of C. jejuni, while N2 plasma is not, at least for the treatment duration. The damage to the membrane was consistent with the reduced viable cell counts in the same samples (Fig. 5b). While N2 plasma treatment did not affect the viable cell counts, air plasma treatment reduced the viable cell counts (p<0.05).
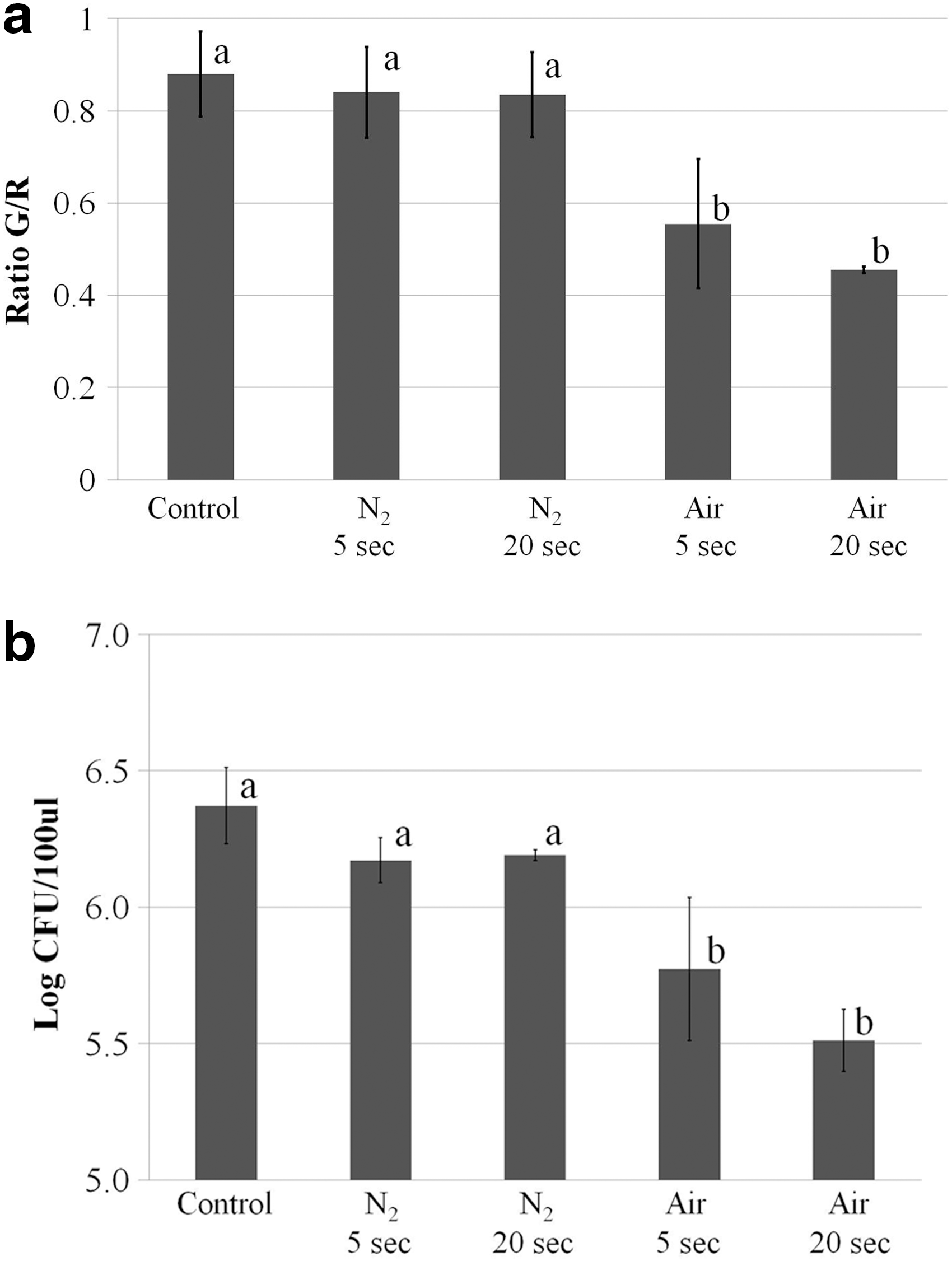
The effect of air or nitrogen plasma on the membrane integrity and viability of Campylobacter jejuni cells. The data are based on two independent experiments.
To visualize the intact and the membrane-damaged cells, confocal microscopic analysis was employed after staining plasma-treated cells with fluorescent dyes (Fig. 6). Consistent with the microplate reader measurement, the ratio of green- to red-fluorescent cells was to a large extent decreased in the sample treated with air plasma for 5 or 20 s compared to the no-treatment control, confirming that air plasma can effectively damage the cellular membranes of C. jejuni (Fig. 6). While most cells were green-fluorescent in the no-treatment control, the air plasma–treated cells were mostly red-fluorescent. Based on individual cell counts, the ratio of green- to red-fluorescent cells was 93/7(%) for the no-treatment control while 9/91(%) and 7/93(%) for 5- and 20-s air plasma-treated cells, respectively. In contrast, little change was found for cells treated with N2 plasma for 5 or 20 s. The ratio of green- to red-fluorescent cells was 96/4(%) and 94/6(%) for 5- and 20-s N2 plasma-treated cells, respectively. However, 2-min N2 plasma treatment noticeably increased the proportion of red-fluorescent cells to 61%, suggesting that prolonged N2 plasma treatment can still damage the cellular membranes of C. jejuni.
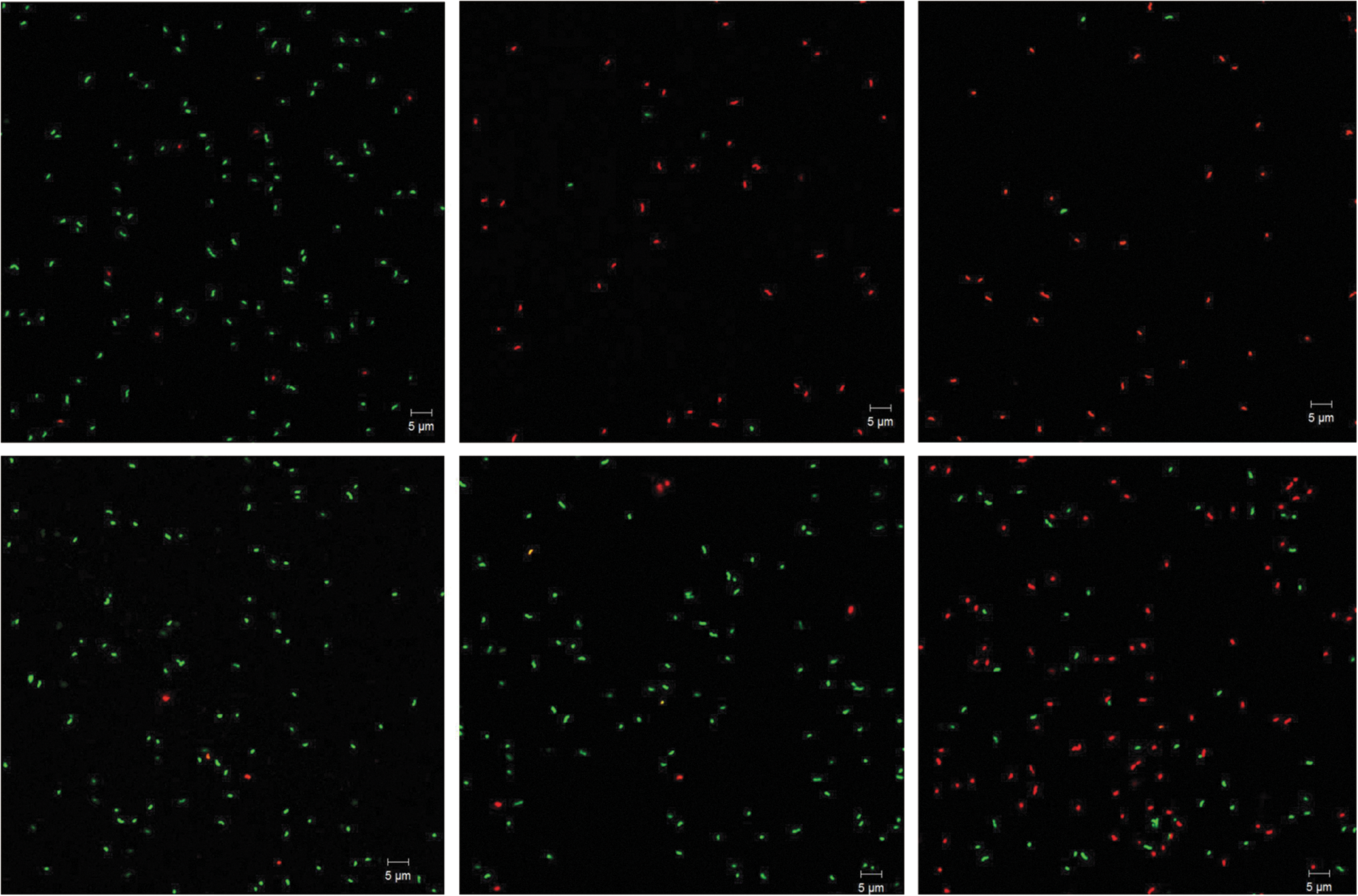
Confocal microscopic analysis of fluorescent-dye-stained Campylobacter jejuni cells after air or nitrogen plasma treatment. The scale bar represents 5 μm. Treatments employed: top left, no treatment control; top middle, air plasma for 5 s; top right, air plasma for 20 s; bottom left, nitrogen plasma for 5 s; bottom middle, nitrogen plasma for 20 s; bottom right, nitrogen plasma for 2 min.
Discussion
Recently, cold atmospheric plasma has received much attention as an emerging nonthermal inactivation method for foodborne pathogens. Because C. jejuni is one of the most common foodborne pathogens worldwide, it is worthwhile to study the parameters of plasma treatment for the efficient inactivation of C. jejuni. A feed gas is one of the factors that can affect the inactivation of foodborne pathogens in plasma treatment. Nevertheless, its effect has not been well studied in plasma for the inactivation of C. jejuni. As shown, the data indicate that air plasma is more effective and destructive than N2 plasma for the inactivation of C. jejuni. The evidence includes (1) significant colony-forming units reduction under air or O2 plasma compared to N2 plasma in short-term treatment, (2) more inactivation under air plasma than under N2 plasma in long-term treatment, and (3) apparent damage to cellular structure and membrane by air plasma in contrast to little to no cell damage by N2 plasma. These data suggest that the feed gas composition for plasma generation can make a significant difference in terms of bacterial inactivation. Our observation is also consistent with previous studies. Niemira (2012b) found that air is more effective than N2 for the inactivation of E. coli O157:H7 and Salmonella isolates on almonds. Several studies also demonstrated that the addition of O2 to nitrogen plasma enhanced sterilization efficacies for bacteria (Deng et al., 2010; Sureshkumar et al., 2010; Kim et al., 2011). Deng et al. (2010) found that the addition of 3.59% O2 to argon plasma generated ozone and increased the intensity of oxygen radicals based on optical emission spectrum. Sureshkumar et al. (2010) found that the addition of 2% O2 enhanced inactivation of Staphylococcus aureus compared to nitrogen gas alone through the emission of nitric oxide and other species. In air plasma, electrical energy-driven collisions between air and electrons generate reactive chemical species through electron impact excitation and dissociation (Laroussi, 2009). The reactive species generated include atomic oxygen (O), singlet oxygen (1O2), hydrogen peroxide (H2O2), ozone (O3), hydroxyl radicals (OH), and nitric oxide (NO) (Laroussi, 2009; Deng et al., 2010; Joshi et al., 2011). Based on these studies, it is assumed that ozone, oxygen radicals, and NO in air plasma may be major contributors to the enhanced inactivation and severe destruction of C. jejuni cells. These chemicals are bactericidal and effective against C. jejuni as well (Oyarzabal, 2005).
A flagellum is an important cellular apparatus in the pathogenesis of C. jejuni. It plays an important role for C. jejuni in the colonization and invasion of animal hosts (Young et al., 2007). A truncated flagellum is often associated with reduced motility and virulence in C. jejuni (Nachamkin et al., 1993; Yao et al., 1994). As shown, the truncated flagella suggests that the plasma can reduce the virulence such as motility of C. jejuni by destroying the cellular structure.
Cell membrane integrity is critical for maintaining viability in bacteria. LIVE/DEAD BacLight Kit is commonly used to assess the membrane integrity of bacterial cells (O'Neill et al., 2004; Berney et al., 2007; Kim et al., 2010). Because dead cells are not necessarily damaged in the cell membranes (Joux and Lebaron, 2000), we used the kit to study whether inactivation of the cells by air plasma treatment is associated with membrane damage. The data clearly show that inactivation of C. jejuni by air plasma treatment is associated with the membrane damage. Thus, it seems apparent that air plasma treatment damaged the cell membranes of C. jejuni and it could be a major mechanism for the inactivation of C. jejuni. Additionally, the membrane damage by the prolonged N2 plasma treatment suggests that it may be a common plasma-induced bacterial inactivation mechanism. Indeed, a recent article also showed that plasma treatment can damage the cell membranes of bacterial species such as S. aureus and Pseudomonas aeruginosa (Kvam et al., 2012). The present study shows that the feed gases used in plasma can make a large difference for the inactivation of bacterial pathogens, and the composition of feed gases is important to increase inactivation efficiency.
Conclusions
This study clearly shows that feed gas composition plays an important role in the inactivation of C. jejuni exposed to plasma as demonstrated with air compared to nitrogen. The oxygen in air appears to be very effective for destroying cellular structures such as membranes and flagella of C. jejuni. This work provides evidence that membrane damage may be a major inactivation mechanism for C. jejuni exposed to plasma.
Footnotes
Acknowledgments
This research was funded by the Korea Food Research Institute. We are thankful to FEMTO Science Co. for providing the DBD plasma device.
Disclosure Statement
No competing financial interests exist.