
Abstract
The aim of the study was to investigate the prevalence, distribution, and diversity of three foodborne bacteria in kiwifruit orchards and processing plants. Fourteen kiwifruit orchards and two processing plants in Shaanxi province were visited for sampling in 2012. Fruit samples and environmental samples in orchards and plants were taken for isolation of Escherichia coli, Staphylococcus aureus, and Salmonella. All isolates were characterized by antimicrobial susceptibility testing and detection of virulence genes. Selected isolates were further examined by pulsed-field gel electrophoresis (PFGE) analysis. In total, 160 E. coli isolates and 14 S. aureus isolates were recovered from 407 samples from orchards and plants, while no Salmonella was recovered. E. coli isolates displayed resistance most frequently to streptomycin (65.6%), and S. aureus isolates displayed resistance most frequently to erythromycin (21.4%). Three E. coli isolates (1.9%) were positive for stx2 and two S. aureus isolates (14.3%) were positive for both seb and seh. Seventy-seven E. coli isolates and 14 S. aureus isolates were analyzed by PFGE. PFGE results showed that both E. coli and S. aureus isolates were diverse, and blades for slicing during the processing could be an important contamination source. This study could provide useful information for kiwifruit growers and industry to establish proper management practices that help minimize the chance of microbial contamination from farm to table.
Introduction
F
Pathogenic Escherichia coli, Salmonella, and Staphylococcus aureus have been responsible for the majority of foodborne outbreaks all around the world. Many of these outbreaks were traced back to environmental reservoir and plant processing (Kaneko et al., 1999; Strawn et al., 2013). In 2006, the outbreak of foodborne E. coli O157:H7 in spinach had a close connection with pre-harvest contamination, and the outbreak-associated E. coli O157:H7 strain was isolated from swine feces, cattle, surface water, sediment, and soil (Steele and Odumeru, 2004; Jay et al., 2007). From 2001 to 2005, the total number of food poisoning cases in South Korea was 33,353, 10.8% of which was caused by enterotoxins produced by S. aureus (Yoon et al., 2011). In the 2008 outbreak of Salmonella Saintpaul infections associated with produce, jalapeño pepper and Serrano pepper were implicated, which highlighted the importance of preventing raw produce contamination (Barton Behravesh et al., 2011).
Fresh produce, including apples, grapes, lettuce, peaches, peppers, spinach, sprouts, and tomatoes, have been shown to harbor various levels of bacterial loads (Nguyen-the and Carlin, 1994; Badosa et al., 2008; Rastogi et al., 2012), but the information about microbial contamination during kiwifruit production has rarely been reported (Balestra, 2008). Kiwifruit is a nutritious fruit mainly grown in China, New Zealand, and Italy. Since China is the biggest kiwifruit exporter in the world, the kiwifruit industry in China is important both domestically and internationally (Testolin and Ferguson, 2009). The main objective of this work was to assess the prevalence, distribution, and diversity of three types of foodborne bacteria in kiwifruit orchards and processing plants in Shaanxi, China to gain useful data for the kiwifruit industry to establish proper management practices.
Materials and Methods
Experimental design and sample collection
Fourteen orchards and two plants in the main kiwifruit production region were selected in Shaanxi province. Orchards and plants were sampled once from September to October 2012. Selected orchards were located in three cities: Zhouzhi (n=8), Meixian (n=5), and Yangling (n=1). Two plants mainly producing frozen kiwifruit slices were located in Zhouzhi city and Baoji city, respectively.
Gloves and boot covers were changed in each orchard and plant, and gloves were disinfected with 75% ethanol prior to sample collection. A total of 407 various types of samples were collected. The samples in kiwifruit orchards included kiwifruits on trees or fallen kiwifruits on the ground, soil, irrigation water, and air. Detailed sampling design of kiwifruit orchards is listed in Table 1. The samples in plants were taken to cover as many processing stages as possible. In the plant in Zhouzhi city, 30 samples were collected from blades for slicing (n=12), air of operation rooms (n=4), water for washing (n=3), kiwifruits without pericarp (n=3), pericarps (n=4), and slices (n=4). In the plant in Baoji city, 29 samples were collected from fresh kiwifruits (n=3), kiwifruits washed with NaClO (n=3), kiwifruits on the conveyor (n=6), frozen kiwifruits (n=3), NaClO solution used for disinfection (n=3), water for washing (n=3), rinse (n=3), and air of operation rooms (n=5).
Escherichia coli isolation and identification
Each sample (25 g or mL) was placed in 225 mL of Buffered Peptone Water (BPW; Beijing Land Bridge Technology Ltd., Beijing, China) and homogenized for 2 min at 320 rpm. After incubation for 10 min at room temperature with periodic shaking, the culture was transferred into a sterile container. After incubating for 3 h at 35°C, the culture was transferred to 225 mL of double-strength Tryptone Phosphate Broth (Oxoid) in a sterile container for incubation. After incubation at 44.0°C for 20 h, a loopful of the culture was streak onto eosin–methylene blue agar (Beijing Land Bridge Technology Ltd.) and MacConkey agar (Beijing Land Bridge Technology Ltd.). After incubation for 20 h at 35 °C, the plates were examined for morphologically typical (with metallic green sheen) E. coli colonies. For each sample, one presumptive E. coli isolate was picked and confirmed by polymerase chain reaction (PCR) detection of uidA gene (Jefferson et al., 1986). These E. coli isolates were stored at −80°C in Luria Bertani (LB; Beijing Land Bridge Technology Ltd.) containing 30% glycerol for further analysis.
Staphylococcus aureus isolation and identification
Isolation of S. aureus was performed according to procedures described previously (Wang et al., 2012). Each sample (25 g or mL) was placed in 225 mL of BPW and incubated at 37°C for 24 h. After pre-enrichment, a 5-mL aliquot of the culture was transferred to 50 mL of Trypticase Soy Broth (Beijing Land Bridge Technology Ltd.) containing 7.5% NaCl. After 18–24 h incubation at 35°C, a loopful of the culture was inoculated onto Baird-Parker agar (Beijing Land Bridge Technology Ltd.) plates with 5% egg yolk and tellurite. After incubation for 20 h at 35°C, plates were examined for morphologically typical (black colonies surrounded by a pale halo) S. aureus colonies. For each sample, one presumptive S. aureus isolate was picked and confirmed by PCR detection of the thermonuclease gene (Rahn et al., 1992). These S. aureus isolates were stored at −80°C in LB broth containing 30% glycerol for further analysis.
Salmonella isolation and identification
Isolation of Salmonella was performed according to procedures described in the National Standards of the People's Republic of China (GB 4789.1-2010). Briefly, each sample (25 g or mL) was placed in 225 mL of BPW and incubated at 37°C for 8–18 h. Ten-milliliter aliquots of pre-enriched cultures were transferred to 100 mL each of Tetrathionate Broth Base (TT; Beijing Land Bridge Technology Ltd.) and Selenite Cystine Broth (SC, Beijing Land Bridge Technology Ltd.), respectively. After incubation at 42°C for 18–24 h in TT broth and in SC broth, one loopful of each broth was streaked onto bismuth sulfite (BS; Beijing Land Bridge Technology Ltd.) and xylose lysine desoxycholate agars (XLD, Beijing Land Bridge Technology Ltd.). The BS plates were incubated at 37°C for 40–48 h and XLD plates were incubated at 37°C for 18–24 h. After incubation, plates were examined for morphologically typical Salmonella colonies. For each sample, one presumptive Salmonella isolate was picked and confirmed by PCR detection of the invA gene (Rahn et al., 1992). These Salmonella isolates were stored at −80°C in LB broth containing 30% glycerol for further analysis.
PCR detection of virulence genes
Six virulence genes specific for each type of diarrheagenic E. coli were tested by PCR: the eae gene for enteropathogenic E. coli, the ipaH gene for enteroinvasive E. coli, the elt and est genes for enterotoxigenic E. coli, the aggR gene for enteroaggregative E. coli, and the stx gene (useful for detection of both stx1 and stx2) for Shiga toxin–producing E. coli (STEC). S. aureus isolates were tested by PCR for nine S. aureus enterotoxin genes (sea, seb, sec, sed, see, seg, seh, sei, and sej), tsst-1 gene, exfoliative toxin genes (eta and etb), Panton-Valentine leukocidin gene, and methicillin resistance gene (mecA) as previously described. Primers for the PCR assays are listed in Table 2. The PCR products were resolved by 1.0% (wt/vol) agarose gel electrophoresis in 0.5×Tris-borate-EDTA (TBE) buffer.
PCR, polymerase chain reaction.
Serotyping
E. coli isolates positive for virulence genes were serotyped in Henan Center for Disease Control and Prevention, China. O and H antigens were characterized using slide agglutination with hyperimmune sera (S&A Company, Thailand) and the serotype was assigned following the manufacturer's instructions.
Antimicrobial susceptibility testing
Antimicrobial susceptibility test of E. coli isolates were performed by the agar dilution method for ciprofloxacin, nalidixic acid, gatifloxacin, ampicillin, gentamicin, kanamycin, amikacin, streptomycin, cefoxitin, cefoperazone, ceftriaxone, trimethoprim/sulfamethoxazole, chloramphenicol, and tetracycline. E. coli ATCC 25922 and Enterococcus faecalis ATCC 29212 were used as quality control strains in each run. The breakpoints used were taken according to guidelines developed by the Clinical Laboratory Standards Institute (CLSI, 2012), with the exception of streptomycin (resistance breakpoint was set at 8 μg/mL).
Antimicrobial susceptibility tests of S. aureus were performed by the agar dilution method for oxacillin, cefoperazone, cefoxitin, erythromycin, gentamicin, chloramphenicol, ciprofloxacin, tetracycline, amikacin, vancomycin, amoxicillin, and nalidixic acid. S. aureus ATCC 25923 and S. aureus ATCC 29213 were used as quality-control strains in each run. The breakpoints used for S. aureus were taken according to guidelines developed by the Clinical Laboratory Standards Institute (CLSI, 2012), with the exception of cefoxitin (resistance breakpoint is set at 8 μg/mL).
Pulsed-field gel electrophoresis (PFGE)
PFGE of XbaI (TaKaRa, Dalian, China)–digested genomic DNA of E. coli isolates was carried out as described previously (Gautom, 1997). The PFGE conditions of XbaI macrorestriction analysis were 6 V/cm for 19 h with pulse times ranging from 2.16 to 54.17 s at a temperature of 14°C and an angle of 120°.
Genomic DNA fingerprints of 14 S. aureus isolates were determined using PFGE as previously described (McDougal et al., 2003). PFGE conditions of SmaI macrorestriction analysis were 6 V/cm for 21 h, with pulse times ranging from 5 to 40 s at a temperature of 14°C and an angle of 120°.
The gels were stained with ethidium bromide, and images were taken under ultraviolet transillumination (Bio-Rad). The images were analyzed using BioNumerics Software (Applied-Maths, Kortrijk, Belgium) by using dice coefficients and the unweighted-pair group method to achieve dendrograms with an optimization value of 0.5% and a 1.0% band position tolerance. The cluster cutoff was set at 85% similarity, and all clusters were designated with a single letter. Genomic DNA of Salmonella serotype Branderup strain H9812 digested with XbaI was used as a molecular size marker.
Results
Isolation of E. coli, S. aureus, and Salmonella
Of the 407 samples, 160 (39.3%) samples were positive for E. coli (Table 3), including 19 (18.3%) of 104 fresh fruits samples, 36 (35.0%) of 103 fallen fruits samples, 72 (69.2%) of 104 soil samples, and 3 (25.0%) of 12 samples from irrigation water in the kiwifruit orchards. Twenty (66.7%) of 30 samples from the kiwifruit plant in Zhouzhi and 10 (34.5%) of 29 samples from the plant in Baoji were also positive for E. coli.
Of the 407 samples, 14 (3.4%) samples were positive for S. aureus (Table 3), including 4 (3.8%) of 104 fresh fruits samples, 1 (1.0%) of 103 fallen fruits samples, and 9 (15.3%) of the 59 samples from 2 plants. Among the nine positive samples in plants, seven were blades for slicing in the plant of Zhouzhi, and two samples were raw kiwifruits in the plant of Baoji. No Salmonella was isolated from all the samples taken in this study.
PCR detection of virulence genes
Among 160 E. coli isolates, 2 isolates recovered from soil in the orchard and 1 from a fallen fruit in orchard were positive for the stx gene. Further test using stx1 and stx2 specific primers showed that both isolates contained only the stx2 gene. No other virulence genes specific for diarrheagenic E. coli were detected among the isolates. Among 14 S. aureus isolates, 2 isolates recovered from raw kiwifruit in the plant in Baoji were positive for seb and seh. None of other virulence genes of S. aureus chosen were detected.
Serotyping
The serotypes of three E. coli isolates containing the stx2 gene were O86a: MH, O158: H21, and O158: H2, respectively.
Antimicrobial susceptibility testing
Of the 160 E. coli isolates, resistance was most frequently observed to streptomycin (65.6%), followed by trimethoprim-sulfamethoxazole (25.6%), and to a less extent cefoxitin (20.6%), ceftriaxone (19.3%), nalidixic acid (16.9%), tetracycline (15.0%), ampicillin (8.1%), ciprofloxacin (5.0%), gatifloxacin (4.4%), gentamicin (2.5%), cefoperazone (2.5%), chloramphenicol (2.5%), and kanamycin (1.3%). All E. coli isolates were susceptible to amikacin (Table 4). One hundred thirty-five E. coli (84.4%) isolates were resistant to at least 1 antimicrobial, and 47 (29.4%) to 3 or more antimicrobials. Among three stx2-positive E. coli isolates, one isolate exhibited resistance to six antimicrobials, while the other two were susceptible to all antimicrobials tested (Table 5).
Minimum inhibitory concentrations (μg/mL) determined via agar dilution in accordance with Clinical and Laboratory Standards Institute (CLSI).
No CLSI breakpoint.
Of the 14 S. aureus isolates, resistance was most commonly detected to erythromycin (21.4%), followed by oxacillin (7.1%), and to a less extent cefoxitin, amoxicillin, and ampicillin (7.1% for each of the 3). All S. aureus isolates were susceptible to cefoperazone, chloramphenicol, ciprofloxacin, tetracycline, amikacin, gentamicin, and vancomycin (Table 6). Of all S. aureus isolates tested, 21.4% were resistant to at least 1 antimicrobial (Table 5).
All isolates were susceptible to cefoperazone, chloramphenicol, ciprofloxacin, tetracycline, amikacin, gentamicin, and vancomycin.
Minimum inhibitory concentrations (μg/mL) determined via agar dilution in accordance with Clinical and Laboratory Standards Institute (CLSI).
No CLSI breakpoint.
PFGE
A total of 77 E. coli isolates, selected based on their sources and places of location, were analyzed for genetic relatedness using PFGE with XbaI. The selected isolates were recovered from the kiwifruit (n=20), soil (n=24), and irrigation water (n=3), plants (n=30). Fifty-four different PFGE patterns were observed among the 77 E. coli isolates. Using a cutoff value of 85% similarity, 11 clusters and 43 individual types were generated (Fig. 1). The three most predominant PFGE clusters were observed in seven isolates grouped in cluster D, 5 isolates in cluster K, and 4 isolates in cluster G.
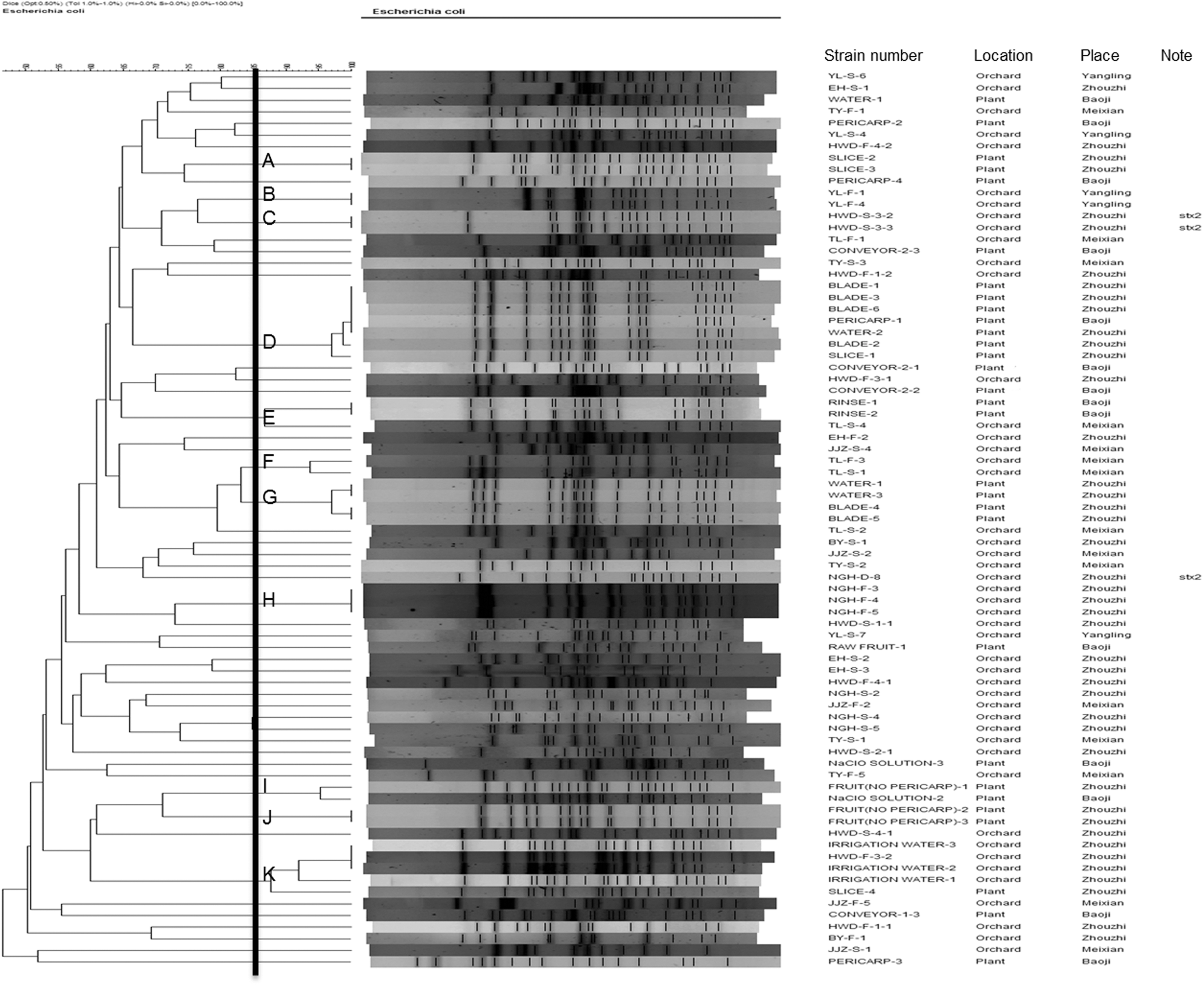
Dendrogram of pulsed-field gel electrophoresis patterns showing the relatedness of Escherichia coli isolates recovered from orchards and plants. The cluster cutoff was set at 85% similarity.
All 14 S. aureus isolates from orchard (n=5) and plant (n=9) were analyzed for genetic relatedness using PFGE with SmaI. Four different PFGE patterns were observed among S. aureus isolates. Using a cutoff value of 85% similarity, three clusters and one individual type were generated (Fig. 2). The most predominant PFGE clusters were observed in nine isolates grouped in cluster A.

Dendrogram of pulsed-field gel electrophoresis patterns showing the relatedness of Staphylococcus aureus isolates recovered from orchards and plants. The cluster cutoff was set at 85% similarity.
Discussion
To our knowledge, field data on the nature of environmental pathogen reservoirs in kiwifruit fields and plants are lacking. Therefore, farmers and food safety professionals have few data on which to base sampling schemes that are intended to detect foodborne pathogens in the preharvest environment and plant environment (Strawn et al., 2013). To maintain kiwifruit consumption safety, knowledge of potential microbial risk from farm to table is a prerequisite for the kiwifruit industry. This study was designed to investigate the distribution of three foodborne bacteria in the kiwifruit orchards and plants. We aimed to obtain meaningful data on microbiological contamination during kiwifruit production and processing and to identify possible sources of contamination.
In this study, Salmonella was not detected in 407 samples from orchards and plants, although some reports showed that the irrigation water, soil, air, and plants are modes of transfer of Salmonella (Nakamura et al., 1997; Hanning et al., 2009; Lapidotand and Yaron, 2009). Considering the limited sampling times (only once) and numbers of samples tested in the present survey, it cannot be inferred that kiwifruits are free of Salmonella contamination. Several studies have shown that E. coli and S. aureus can survive for hours or even days on hands, sponge/cloths, or utensils (Kusumaningrum et al., 2003; Wilks et al., 2005). The existence of E. coli and S. aureus were found both in orchards or processing plants. We also noticed that E. coli was more prevalent and diverse than S. aureus, considering the rate of isolation and PFGE patterns. This may indicate that E. coli has more sources for contamination. Although only three potentially pathogenic (STEC) isolates were detected among all E. coli isolates and posed potential hazards to consumers, the relatively high prevalence of E. coli still highlighted the importance of keeping good agriculture practices, since E. coli has been considered a good indicator of fecal contamination. The isolation rates of both E. coli and S. aureus in plants are higher than those in orchards, which suggest that postharvest processing contamination should also be paid attention to, although the preharvest stage is an important period of contamination. An interesting finding of our study is that we found the existence of E. coli even in the NaClO solution used for disinfection. Even though the initial concentration of NaClO used is 100–150 ppm, the concentration of NaClO may decrease greatly after long-time use without change (Taormina and Beuchat, 1999).
The antimicrobial resistance rates of E. coli isolates in this study were similar to those reported for E. coli isolates recovered from raw vegetables in the Czech Republic (Skockova et al., 2013), while lower than those reported for E. coli isolated from animals (Tadesse et al., 2012). The large amount of antimicrobial used in animal husbandry may partly account for the difference of antimicrobial resistance rate between produce-source E. coli and food animal–source E. coli. It is assumed that some E. coli isolates may come from fecal contamination from wild animals roaming in the orchards or birds, which have rare antimicrobial contact; therefore, a relatively low resistance rate was observed in these isolates.
Only three (1.9%) E. coli isolates were found to carry stx2, and other virulence genes for diarrheagenic E. coli were not detected. This positive rate was lower than the rate reported in a Mexico study in which 8.9% of food-source E. coli isolates carry stx genes (Canizalez-Roman et al., 2013). Currently available data indicate that strains that produce only stx2 are more commonly associated with hemolytic uremic syndrome infections than strains that produce both stx1 and stx2, or only stx1. The E. coli isolates carrying stx2 were from soil and fallen fruits in the orchards, but we lack the evidence to confirm whether the feces of wild animals or birds are the source of these E. coli isolates (Fukushima et al., 1999). Some reports showed that in feces from cattle, chicken, and pigs there was a high positive rate of STEC (Horton et al., 2011; Amézquita-López et al., 2012). In China, some farmers would prefer to use the feces of livestock as fertilizer to reduce fertilizer expenses, and we did find feces in some orchards during sampling. This practice greatly increases the chance of microbial contamination, and its use should be discouraged. Previous reports (Rhee and Woo, 2010) analyzing the enterotoxin genes (sea through sej), eta and etb genes, and tsst gene in foodborne S. aureus revealed a wide diversity in the virulence gene patterns. In the present study, only 14.3% of strains carried toxin genes tested, compared to 29.6% for food samples in Korea (Rhee and Woo, 2010) and 84.9% for food samples in the United States (Pu et al., 2011). The difference in type of samples, origin of samples, and environments may contribute to the difference in the rates.
The PFGE patterns of E. coli isolates revealed genetic diversity between different isolates. Several identical patterns were found from different types of samples inside the processing plants. A plausible explanation for the finding is that there was cross-contamination among blades, water, and fruits during the processing. Some isolates from irrigation water and kiwifruits were identical, indicating that irrigation water may be a reservoir and a transmission pathway for contamination. This finding was consistent with results from previous studies (Oliver et al., 2005; Lyautey et al., 2010). PFGE patterns of S. aureus isolates from two plants showed high similarity among each other. One uniform PFGE pattern for isolates recovered from different blades for slicing inside the plant in Zhouzhi provides the evidence again that blades for slicing are an important source of contamination during processing.
Conclusions
This study demonstrated the presence of E. coli and S. aureus in the kiwifruit orchards and plants in Shaanxi Province, China, while no Salmonella was detected. E. coli was distributed in both orchards and plants and S. aureus mainly existed in the plants. Irrigation water and slicing blades could be potential sources of microbial contamination during the kiwifruit supply chain. These findings provide useful information for kiwifruit growers and industry to establish proper management practices that help minimize the chance of microbial contamination from farm to table. For instance, reduction of the use of raw manure and use of good-quality irrigation water are useful to reduce contamination during the growth stage. Regular monitoring of the chlorine concentration and disinfection of the blades for slicing are strongly recommended during kiwifruit processing.
Footnotes
Acknowledgments
This work was supported in part by the Twelve-Five Science and Technology Support Program (No. 2012BAK17B06), New Century Excellent Talent Support Plan (NCET-13-0488), International Collaboration Partner Plan (No. A213021203) in Northwest A&F University, and Science and Technology Development Plan Program of Shaanxi Province (No. 2013KJXX-16).
Disclosure Statement
No competing financial interests exist.